











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkSe cree que, al igual que Aedes (Stegomyia) aegypti (Linnaeus, 1762), Ae. (Stegomyia) albopictus ( Skuse, 1894) es una especie importante en la transmisión de arbovirus como los del dengue, la fiebre amarilla, el chikungunya y el Zika en diversas regiones del mundo. No obstante, su papel real como vector en Latinoamérica apenas comienza a estudiarse (1).
La fiebre por dengue se reconoce ampliamente como una de las infecciones virales con mayor incidencia en el mundo, tal como ocurre en grandes ciudades de Colombia, como Medellín, Cali y Cúcuta, entre otras. La infección es causada por algunos de los genotipos que componen el acervo genético de los cuatro serotipos del virus del dengue (DENV-1 a DENV-4). El surgimiento de los focos epidémicos de dengue y de la fiebre hemorrágica por dengue se ha asociado a la presencia de uno cualquiera de estos cuatro serotipos, así como a la circulación simultánea de genotipos debida a la colonización o al surgimiento de linajes más patógenos en un periodo determinado (2). Se han descrito cinco genotipos epidémicos del DENV-2: el asiático I (AS-I), el asiático II (AS-II), el asiático-americano (AS/AM), el cosmopolita (COS) y el americano (AM) (3), de los cuales el AM y el AS/AM son los más frecuentemente asociados a focos epidémicos de dengue en muchas regiones tropicales y subtropicales de Suramérica, incluida Colombia (4).
Aedes albopictus es originario del oriente de India (5), desde donde se dispersó por el oeste hasta las islas Mauricio y Madagascar y, posteriormente, colonizó múltiples regiones en el continente americano. En América se ha reportado desde 1981 en los Estados Unidos (6), país en el cual se distribuye ampliamente, incluso en zonas con presencia de Ae. aegypti, por lo que representa un gran riesgo para la población y un importante reto para las autoridades de salud (7). Aedes albopictus se encuentra también en México (8) y en diversos países de Centroamérica. En Suramérica, Ae. albopictus se detectó en Brasil en 1986 (9), donde la especie se ha distribuido rápidamente hacia zonas urbanas (10), y se ha evidenciado como vector del virus del dengue (11). También se ha reportado en Argentina (12), desde donde se cree que ha seguido dispersándose hacia las regiones más al norte del Cono Sur. Aunque su rango de expansión es aún difícil de precisar, continúa siendo objeto de interés por su potencial papel como vector (1,5).
En Colombia, Ae. albopictus se reportó inicialmente en 1998 en la ciudad de Leticia, en el departamento de Amazonas, limítrofe con Brasil (13). Años más tarde, se reportó en Cali (14) y, más recientemente, en el municipio de La Tebaida, departamento del Quindío (15), y en Istmina, departamento del Chocó, limítrofe con Panamá (16). En Medellín, la presencia de Ae. albopictus ha sido constante desde el 2011, y frecuentemente se han encontrado individuos en zonas céntricas de la ciudad como el Jardín Botánico Joaquín Antonio Uribe (17) y el campus de la Universidad Nacional de Colombia, sede Medellín (18).
A pesar de su importancia como potencial vector del dengue y otros arbovirus en Colombia, la información sobre su ecología y evolución, así como sobre su papel epidemiológico, es aún escasa. El único trabajo publicado hasta la fecha sobre la infección viral en Ae. albopictus en el país, se hizo en individuos recolectados en el municipio de Buenaventura (Valle del Cauca) durante los años 2002 y 2004 (19). En ese estudio se reportó la infección natural de Ae. albopictus con el DENV-1 y el DENV-2, y se sugirió su posible papel en la dinámica de trasmisión del dengue en esa localidad (19). En el resto del país, a pesar de que la especie se viene reportando cada vez con mayor frecuencia, persiste un gran desconocimiento sobre su capacidad de infección, así como sobre los serotipos y los genotipos virales circulantes.
En el presente trabajo se reporta la infección de Ae. albopictus con el genotipo asiático-americano del DENV-2 en Medellín y se aporta información sobre el posible origen de este vector en el Valle de Aburrá.
Materiales y métodos
Insectos y extracción de ADN
Se recolectaron nueve individuos en áreas de fitotelmata en un relicto de bambú americano (Guadua sp.) y en un árbol de jardín (Ficus sp.) (figura 1) durante enero y febrero de 2016 en el campus de la Universidad Nacional de Colombia, localizado en el área urbana de la ciudad.

Figura 1 Lugares de recolección de Ae. albopictus en el campus de la Universidad Nacional de Colombia, sede Medellín (coordenadas geográficas: 6°26’07,5’’N, -75°57’69,8’’O; World Geodetic System, WGS 84) Fotos: Alejandra M. Clavijo
Los mosquitos se almacenaron vivos en tubos cónicos de 1,5 ml hasta su identificación taxonómica con la clave de Forattini (20). A cinco de los nueve mosquitos se les removieron las dos patas posteriores para su validación molecular usando el gen citocromo oxidasa I (COI). Se extrajo el ADN total de cada ejemplar utilizando solución tampón de macerado según protocolo previamente reportado para mosquitos (21). Los restos de los mosquitos se almacenaron en RNAlater ® en grupos compuestos por uno a cuatro individuos según el lugar de su captura, para su posterior procesamiento.
Amplificación del gen COI en Aedes albopictus
Se amplificó un fragmento de 700 pares de bases (pb) del gen COI, propuesto como el código de barras genético en diferentes organismos (22), mediante reacción en cadena de la polimerasa (PCR) usando los iniciadores MTNF y MTNR, como se ha reportado previamente (23).
Las condiciones de amplificación consistieron en una etapa inicial de desnaturalización a 94 °C durante 10 minutos, seguida de 35 ciclos a 95 °C durante 60 segundos, a 50 °C durante 60 segundos y a 72 °C durante otros 60 segundos,y una etapa de extensión final a 72 °C durante cinco minutos.
Los productos de la amplificación se visualizaron en un gel de agarosa al 1 %, teñido con bromuro de etidio. Todos los productos positivos se purificaron y se enviaron a la empresa Macrogen (http://dna.macrogen.com) para su secuenciación.
Extracción del ARN, síntesis del ADN complementario y tipificación del DENV
En cada grupo de mosquitos se extrajo el ARN total usando el estuche RNeasy Mini Kit ® (Qiagen) según el protocolo recomendado por el fabricante. Para la síntesis del ADN complementario de un fragmento de 470 pb del gen NS3, se hizo una PCR con transcripción inversa (reverse transcription-PCR, RT-PCR) usando el iniciador DV3 (5’-AARTGIGCYTCRTCCAT-3’) previamente reportado para dengue (24). Las condiciones de la RT-PCR consistieron en un volumen final de 20 µl de una mezcla que contenía 10 µl de ARN total, 4 µl de solución tampón (5x), 2 µl de dNTP (10 mM), 2 µl del iniciador DV3 (10 µM), y 2 µl de 20 x 10 3 U/µl de transcriptasa inversa (Thermo ®). Esta mezcla se amplificó durante un ciclo de 60 minutos a 42 °C seguido por un ciclo de 10 minutos a 90 °C, según protocolo descrito previamente (24).
El DENV se tipificó mediante una PCR semianidada en dos pasos: inicialmente se usaron los iniciadores DV1 (5’-GGRACKTCAGGWTCTCC-3’) y DV3 (24) para amplificar un fragmento de 470 pb del gen NS3. Las condiciones de la PCR consistieron en un volumen final de 25 µl que contenía 3 µl de ADNc, 2,5 µl de solución tampón (10x), 2 µl de MgCl2 (25 mM), 2,5 µl de dNTP (2 mM), 0,5 µl de los iniciadores DV1 y DV3 (10 µM), y 5U/µl de polimerasa (Thermo®).
El perfil de amplificación consistió en una etapa inicial de desnaturalización a 95 °C durante un minuto, seguida de 30 ciclos a 95 °C durante 30 segundos, a 50 °C durante 60 segundos y a 72 °C durante 40 segundos para cada uno y, por último, una extensión a 72 °C durante cinco minutos.
Posteriormente, se hizo una segunda amplificación que incluyó el iniciador DV1 y los iniciadores específicos DSP1 (5’ - AGTTTCTTTTCCTAAACACCTCG - 3’); DSP2 (5’ - CCGGTGTGCTCRGCYCTGAT - 3’); DSP3 (5’ - TTAGAGTYCTTAAGCGTCTCTTG - 3’) y DSP4 (5’ - CCTGGTTGATGACAAAAGTCTTG - 3’), los cuales amplifican tamaños de 169, 362, 265 y 426 pb para cada uno de los cuatro serotipos del DENV, respectivamente (25).
En esta PCR se empleó un volumen final de 25 µl que contenía 3 µl de ADNc, 2,5 µl de solución tampón (10x), 1,5 µl de MgCl2 (25 mM), 2,5 µl de dNTP (2 mM), 1,25 µl del iniciador DV1 y de cada uno de los iniciadores específicos DSP1 a DSP4 (10 µM), y 5 U/µl de polimerasa (Thermo ®).
El perfil de amplificación consistió en una etapa inicial de desnaturalización a 95 °C durante cuatro minutos, seguida de 35 ciclos a 95 °C durante 30 segundos, a 52 °C durante 60 segundos y a 72 °C durante 30 segundos para cada uno; finalmente, se hizo una etapa de extensión a 72 °C durante cinco minutos.
Los productos de la amplificación se visualizaron en un gel de agarosa al 1 % teñido con bromuro de etidio, y los productos positivos se enviaron, para su posterior purificación y secuenciación, a la empresa Macrogen (http://dna.macrogen.com).
Análisis de secuencias y tipificación genética del DENV
Las secuencias de nucleótidos obtenidas tanto para el gen COI en Ae. alpopictus como para el gen NS3 del DENV, se ajustaron manualmente usando el programa Bioedit, versión 7.0 (26); todos los alineamientos se hicieron usando la opción “auto” del programa MAFFT, versión 7.2 (27).
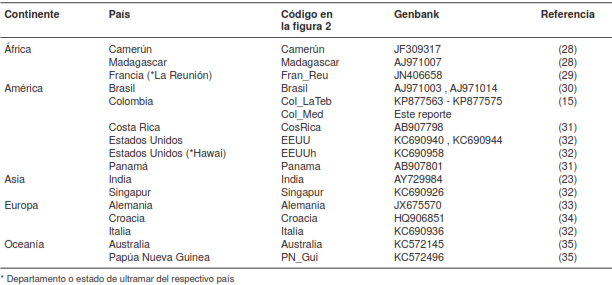
Las secuencias del COI se compararon con 17 secuencias tomadas del Genbank obtenidas de individuos de 14 países de diferentes regiones de África, América, Asia, Europa y Oceanía (cuadro 1). Además, se incluyeron 13 secuencias de individuos recolectados en el municipio de La Tebaida, Quindío (15). Para la confirmación molecular de la taxonomía de Ae. albopictus, se construyó una red haplotípica usando el algoritmo median-joining con el programa Network, versión 5.0 (http://www.fluxus-engineering.com/). Para la identificación del genotipo del DENV, las secuencias obtenidas se compararon con las de la base de datos del genoma del DENV mediante el programa Blastn (https://blast.ncbi.nlm.nih.gov/Blast.cgi).
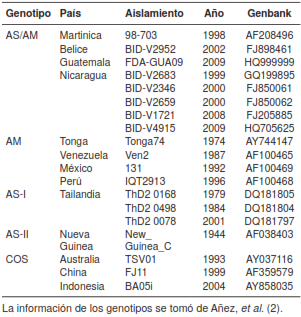
Posteriormente, se hizo el alineamiento de la secuencia parcial de 307 pb del NS3 con 19 secuencias (cuadro 2) de los genotipos reportados en América (2). A partir de la base de datos compuesta por las secuencias del gen NS3 del DENV, se estimó la distancia génica pareada basada en el modelo de sustitución de dos parámetros de Kimura (K2-p) (36) y a continuación se construyó un dendrograma de distancias para cada conjunto de datos basado en el algoritmo neighbor-joining con el programa MEGA, versión 6.0.6 (37). El sustento estadístico de los nodos se probó mediante bootstrap con 1.000 iteraciones (38).
Resultados
Análisis del gen COI en Aedes albopictus de Medellín
Se encontró un mismo haplotipo para el fragmento del gen COI analizado en los cinco individuos de Ae. albopictus recolectados en el municipio de Medellín, similar al observado en la mayoría de los individuos colombianos del municipio de La Tebaida en el Quindío (15) y de un individuo de la isla de Papúa, Nueva Guinea (35) (figura 2). No se encontró ninguna diferencia significativa entre los haplotipos de los distintos orígenes geográficos, ya que no los separan más de cuatro pasos de mutación (figura 2 ).
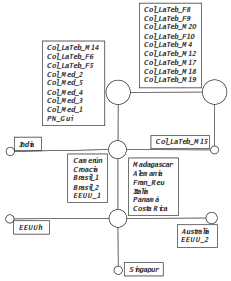
Figura 2 Red haplotípica estimada mediante el algoritmo median-joining del gen COI en Ae. albopictus. El tamaño de los círculos indica la frecuencia haplotípica y las líneas que conectan los haplotipos indican un paso mutacional. Los detalles del origen de las secuencias se presentan en el cuadro 1.
Genotipificación del DENV-2 e identificación del genotipo viral
Solo en uno de los tres grupos de insectos evaluados, la amplificación tuvo un tamaño similar al esperado para la presencia del DENV-2 (figura 3). El análisis filogenético del DENV-2 ubicó la secuencia identificada en Ae. albopictus de Medellín dentro del clado compuesto por los aislamientos pertenecientes al genotipo asiático-americano, con un alto sustento de nodo (bootstrap=100) y claramente diferenciado de los restantes genotipos (figura 4). Cabe resaltar que dentro del genotipo asiático-americano la secuencia del fragmento del gen NS3 de Ae. albopictus mostró más similitud con el aislamiento más antiguo (1998) e insular (Martinica) que con los restantes, más recientes y continentales (figura 4).
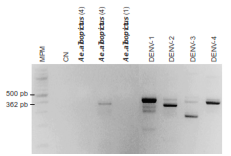
Figura 3 Gel de agarosa teñido con bromuro de etidio en el que se visualiza la amplificación de un fragmento aproximado de 362 pb indicativo de la presencia de DENV-2 en un grupo de individuos de Ae. albopictus recolectados en Medellín. Los números entre paréntesis indican el número de insectos incluidos en cada grupo según el sitio de recolección. MPM: marcador de peso molecular; CN: control negativo
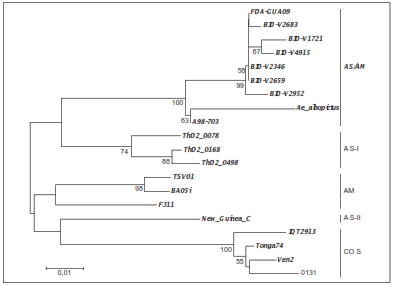
Figura 4 Dendrograma de distancias genéticas basadas en K2-p de un fragmento de 307 pb del gen NS3 del DENV-2. Se muestran los clados correspondientes a los genotipos del DENV-2 reportados en Añez, et al. (2); se incluye el encontrado en Aedes albopictus de Medellín en este trabajo. Los números bajo los nodos indican el valor de bootstrap obtenido de 1.000 iteraciones (solo se muestran los valores >50)
Discusión
El tigre asiático, como se conoce a Ae. albopictus, es considerado uno de los vectores de arbovirus más invasivos y ampliamente distribuidos en el mundo. Desde su ingreso al continente americano, presumiblemente en la década de los años 80, este insecto ha ido incrementando paulatinamente su distribución geográfica en áreas densamente pobladas con buenas condiciones de saneamiento, lo que llama la atención sobre su real papel en la emergencia y reemergencia de importantes virosis como el dengue, el chikungunya y el Zika.
A diferencia de Ae. aegypti, sobre cuyo origen y procesos de dispersión en América se han propuesto distintas hipótesis (39), hasta el momento no hay un consenso claro sobre la historia demográfica de Ae. albopictus y, menos aun, sobre los procesos que dieron origen a las poblaciones americanas (32). A pesar de que el primer registro de esta especie en Colombia data de más de 19 años, muy poco se ha estudiado su composición y estructura poblacional. En el trabajo más reciente se sugiere que las poblaciones colombianas, particularmente las del municipio de La Tebaida en Quindío, tienen un origen asiático (de Singapur) (15), dada la similitud de nucleótidos de los haplotipos del gen COI en este municipio, con dos haplotipos de Singapur reportados en un estudio anterior (32).
En este trabajo se encontró que los individuos de Ae. albopictus recolectados en Medellín presentaban el mismo haplotipo del COI reportado para varios de los individuos de La Tebaida e, igualmente, se observó una relación cercana con los demás haplotipos de Ae. albopictus en diversos países del mundo. Estos resultados aportan información comprobada en apoyo de la hipótesis de un único origen ancestral asiático de las poblaciones colombianas (ampliamente aceptado) y eventualmente permitirían sugerir una composición panmítica de las poblaciones de este vector en Colombia. No obstante, debido a la ausencia de información concluyente y de un tamaño de muestra representativo, no es posible establecer un país o lugar puntual de origen, ni una clara estructura o composición poblacional de Ae. albopictus en Colombia. En este sentido, llamamos la atención sobre la necesidad de desarrollar estudios que indaguen sobre su filogeografía y estructura poblacional en el país.
La presencia de Ae. albopictus naturalmente infectado con DENV-1 y DENV-2 en Colombia, se reportó inicialmente en el municipio de Buenaventura, Valle del Cauca (19). Allí se observó la concurrencia de Ae. albopictus y Ae. aegypti durante el periodo de estudio (2002-2004), así como una correlación positiva entre la tasa de infección entomológica y el incremento en el número de casos positivos de dengue en humanos (19). Aunque estos resultados no demuestran la incriminación vectorial de Ae. albopictus, su participación en la dinámica de transmisión del dengue a los humanos no puede descartarse.
En el presente estudio se reporta la presencia de Ae. albopictus infectado con DENV-2 en Medellín. Este hallazgo concuerda con lo reportado en Buenaventura, por lo cual también podría considerarse su eventual papel en la dinámica de transmisión del dengue en esta ciudad. Además, los resultados presentados indican la circulación del genotipo asiático-americano del DENV-2 en Ae. albopictus de Medellín. Este es el genotipo más frecuente en América desde los años 90, y en Colombia se ha reportado frecuentemente en pacientes febriles con cuadro clínico indicativo de dengue en cuadros todo el país (4), por lo que se sugiere que Ae. albopictus puede tener un papel en la transmisión del dengue a los humanos. Sin embargo, esta hipótesis, así como el grado de riesgo epidemiológico atribuible a Ae. albopictus, deberá comprobarse en futuros estudios.
Teniendo en cuenta el potencial papel de Ae. albopictus como vector en Colombia, se propone que esta especie se incluya en las estrategias de vigilancia y control vectorial del dengue en el país. En este sentido, es importante mencionar que desde el hallazgo de Ae. albopictus en el campus de la Universidad Nacional de Colombia, sede Medellín, en enero y febrero de 2016, periódicamente se hace la revisión y la descripción de sus criaderos, así como la recolección de adultos para el estudio de diversos aspectos de su ciclo de vida bajo condiciones de laboratorio, y la evaluación de su infección con el virus del dengue mediante RT-PCR.

