











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
El gen SLC6A3 codifica la proteína transmembrana DAT1 que tiene como función recapturar dopamina desde el espacio sináptico, por lo que es considerado fundamental para la finalización de la neurotransmisión dopaminérgica en las terminaciones presinápticas1. Las vías reguladas por el neurotransmisor de dopamina han sido implicadas en la regulación neuroendócrina y alimentaria, la actividad locomotora, la afectividad y la emotividad2. La región no traducida UTR3' de SLC6A3 es una región variable en el que una secuencia puede repetirse múltiples veces en tándem, este polimorfismo se denomina Número Variable de Repeticiones en Tándem (VNTR). En esta región, este gen presenta un VNTR de 40 pb que puede repetirse entre 3 a 13 veces3. Aunque la distribución de estos VNTR varía significativamente entre poblaciones, en Latinoamérica, los más frecuente son los de 10 (10 %-85 %) y 9 repeticiones (15 %-22 %)4. En contraste, en Europa se presenta en una baja frecuencia (1 %) los alelos de 6 y 11 repeticiones4,5.
Estas variaciones polimórficas no afectan la secuencia de aminoácidos del transportador DAT1, pero pueden afectar la expresión génica al influenciar la estabilidad, el transporte, maduración y degradación del ARNm6. Se ha reportado que ciertos VNTR en DAT1, pueden afectar la recaptación de dopamina, prolongando su presencia en la hendidura sináptica y activando mecanismos de regulación negativa que conducen a su degradación e inestabilizan sus receptores postsinápticos7. Así, concentraciones elevadas de dopamina podrían contribuir a una menor actividad dopaminérgica, potencialmente causando trastornos neuropsiquiátricos relacionados con el agotamiento de dopamina como la depresión y la ansiedad7. Estudios en adultos con síntomas de depresión han observado una mayor frecuencia significativa del genotipo 9R/9R, asimismo en este grupo el genotipo 10R/10R genotipo se relacionó con síntomas de ansiedad severa y moderada8. A su vez han relacionado que portar el alelo 10R facilita la respuesta a la terapia antidepresiva9. En contraste, en niños con síntomas de ansiedad social relacionado al trastorno del espectro autista, el genotipo 10R/10R tuvo mayor frecuencia10.
Para el 2017 se estimó que 4,4 % de la población mundial sufrió un trastorno depresivo y 3,6 %, un trastorno de ansiedad11. En Colombia para el 2015 se estimó una prevalencia de 5.4 % de depresión y 3.9 % de ansiedad en la población entre los 14 -44 años, teniendo mayor relevancia la región pacífica con 7.1 % y 3.9 % respectivamente12. Estas cifras se incrementaron en adultos (18-29 años) en 48 % depresión y 37 % ansiedad durante el periodo de confinamiento por el COVID 19 a nivel mundial13.
Si bien aún no existe un consenso con respecto a la contribución de los VNTR de DAT1 en la susceptibilidad a trastornos neuropsiquiátricos como la depresión y la ansiedad14, estos trastornos son cada vez más frecuentes en población joven universitaria generando ausentismo, bajo rendimiento académico, abandono universitario o incluso casos de suicidio15. Por lo tanto, es muy importante estudiar la frecuencia de estos polimorfismos en poblaciones con bajos niveles de depresión y ansiedad, información de base que permita realizar comparaciones con otras poblaciones universitarias y futuros estudios de susceptibilidad a estos trastornos. Por esta razón, esta investigación tuvo como objetivo describir la frecuencia de polimorfismos VNTR en la región UTR 3' del gen DAT1 en una muestra de estudiantes de pregrado con bajos niveles de depresión y ansiedad de una universidad privada de Cali, Colombia.
Materiales y Métodos
Diseño y población estudio
Estudio transversal, con enfoque cuantitativo y de diseño descriptivo. Con un modelo de selección de muestra no probabilístico y de participación voluntaria en población universitaria de la Pontificia Universidad Javeriana sede Cali- Colombia. Se invitaron 143 estudiantes universitarios que estuvieran matriculados entre segundo y décimo semestre de pregrado en el año 2018. Fueron seleccionados 62 estudiantes, los cuales cumplieron los siguientes criterios de inclusión: 1) edad entre 18 -25 años, 2) bajos niveles de depresión y ansiedad según los inventarios de Beck. Los criterios de exclusión fueron: 1) consumo diario o semanal de sustancias psicoactivas como marihuana, cocaína, alucinógenos, tranquilizantes y estimulantes, 2) consumo diario o semanal de alcohol.
Instrumentos
Fueron considerados sujetos con bajos niveles de depresión aquellos que obtuvieron un puntaje inferior de 19 en el inventario Beck Depression Inventory (BDI) versión II16 validada en población universitaria colombiana17. Los estudiantes fueron clasificados con bajos niveles de ansiedad cuando el puntaje en el inventario Beck Anxiety Inventory versión original (BAI)18 fue inferior a 15, inventario que ha sido validado en población joven universitaria19,20,21. El protocolo de evaluación de los inventarios fue presentado a cada participante de forma individual, por asistentes de investigación, formados en el campo de la psicología, previamente entrenados por las investigadoras del estudio, para la presentación y recolección de la información. La calificación e interpretación de los resultados de las escalas BDI-II y BAI se realizaron de acuerdo con las instrucciones de los manuales originales16,18.
Adicionalmente, para indagar la condición médica de los participantes relacionado al sistema nervioso central, se realizó un cuestionario adicional en donde se preguntó si la persona alguna condición médica o se encontraba bajo algún tratamiento farmacológico, en caso afirmativo se solicitó detallar la condición médica y el nombre del medicamento. Por otro lado, el consumo de sustancias psicoactivas (SPA) se exploró mediante el cuestionario de estilos de vida en jóvenes universitarios (CEVJU-R), un cuestionario de autorreporte validado en población universitaria colombiana22, el cual incluye 7 preguntas sobre el consumo regular de SPA evaluado en una escala Likert: no consumo, consumo ocasional (social), una vez por semana, una vez al día o varias veces al día.
Consideraciones éticas
Todos los participantes firmaron un consentimiento informado, el cual contó con la aprobación del comité de ética institucional. Además, este estudio se acogió a la resolución 8430 de 1993 y a la 2378 de 2008 de la legislación colombiana, y a la declaración internacional de Helsinki.
Toma de muestras de sangre
El día previo a la toma de muestra de sangre, se solicitó a los participantes dormir un promedio de 8 horas, no consumir cafeína, ni sustancias psicoactivas, por lo menos, 12 horas antes. En total, se realizaron 6 jornadas de toma de muestras, entre las 7 am y 11 am. Las muestras de sangre fueron colectadas por tres enfermeras de una institución de salud, en tubos anticoagulados con EDTA, para ser posteriormente almacenados a -80°C.
Extracción de ADN
La extracción del ADN se realizó a partir de sangre periférica se realizó con el DNeasy Blood & Tissue Kit (Cat No. 69506 QIAGEN) siguiendo las instrucciones del fabricante, y la concentración se determinó espectrofotométricamente, midiendo la absorbancia (A) a 260 nm, según la siguiente fórmula: [ADN ng/μL] = A260 nm x 50 ng/μL. La pureza del ADN se estimó con la relación A260 nm/ A280 nm. Luego, el ADN fue almacenado a -30 °C.
Genotipificación de polimorfismos
La genotipificación del VNTR en la región UTR 3' de DAT1 mediante PCR convencional, usando dos cebadores con las siguientes secuencias: F:5'-TGTGGTGTAGGGAACGGCCTGAG-3' y R:5'CTTCCTGGAGGTCACGGCTCAAGG-3' descritos originalmente por Waldman et al (23). La PCR se realizó utilizando dos DNA polimerasas: la Phusion™ High-Fidelity DNA Polymerase (Thermo Scientific, catálogo No. F553L) y la GenTaq (laboratorio de genética y biología molecular Colombia LTDA). La mezcla de la reacción contenía: buffer de amplificación 1x, 0.2 mM dNTPS, 0.1 - 0.15 pM de cada cebador, 1.5 - 2.5 m MgCl2 y 0.2 - 0.33 UI de DNA polimerasa para la Taq Phusion y GenTaq respectivamente. La cantidad de ADN genómico adicionado fue 100 ng. Las condiciones de amplificación incluyeron una desnaturalización inicial de 5 min a 95 °C en el caso de la Gen Taq o 30 s a 98 °C para la Taq Phusion, seguido por 35 ciclos a 95 °C por 30s (Gen Taq) o 98 °C por 10 s, el anillamiento 64 °C por 30 s, 72 °C por 30 s. Para la Taq Phusion la etapa de anillamiento y extensión se realizó en un solo paso a 72 °C por 35 s, la extensión final fue a 72 °C por 10 min para ambas polimerasas. En cada una de las PCR se corrió un control negativo, para descartar contaminación de las muestras.
Los productos de PCR fueron separados en geles de agarosa al 2 % con buffer Tris-EDTA 1x, a 70 Voltios por 90 minutos, usando marcadores de peso molecular escalonados cada 50 bp (Nippon Genetics,). El gel fue teñido con la tinción Fluorescent Stainig Dye (Smobio) y visualizado en foto-documentador a 260 nm. Para establecer los VTNRs, se comparó el tamaño y número de las bandas con el patrón de los marcadores de peso molecular como control de que la PCR se realizó correctamente. Los tamaños esperados según el número de repeticiones fueron los siguientes: 320 pb (6 R), 360 pb (7 R), 400 pb (8 R), 440 pb (9 R), 480 pb (10 R) y 520 pb (11 R). La evaluación fue realizada por dos evaluadores independientes. En caso de discordancia se repitió la amplificación y la electroforesis.
Análisis estadístico
Las frecuencias alélicas de los VNTR para DAT1, se determinaron con base al total de muestras analizadas. Las relaciones entre las frecuencias alélicas y genotípicas por sexo se estimaron con Chi-cuadrado (X 2). El equilibrio Hardy-Weinberg fue determinado usando una prueba exacta multi alélica con un nivel de significancia de p > 0,05 (24), con el fin de establecer si las frecuencias genotípicas en función del alelo están constituidas por sujetos independientes que no tienen grado de consanguinidad, y no han presentado la aparición de nuevos alelos o la fijación de estos25. El análisis se realizó con el lenguaje de programación R con la plataforma R-studio versión 4.0.226
Resultados
Se evaluaron 143 estudiantes universitarios, de los cuales se excluyeron 37 por presentar valores mayores en el BDI (>19) y BAI (>15), y 44 sujetos no aceptaron seguir participando. Por lo tanto, se genotipificaron los VNTR del gen SLC6A3 para DAT1 de 62 sujetos que cumplieron los criterios de inclusión y decidieron participar en el estudio. Los estudiantes cursaban entre tercer y séptimo semestre, la media de edad fue de 20,1 (± 1,4), siendo mujeres el 52 %. Los puntajes para BDI en mujeres y hombres fueron 9,2 (± 5,3) y 7,2 (± 3,7), para BAI fueron 8,41 (± 4,85) y 6,50 (± 3,72), por lo que la muestra estaba constituida por sujetos con bajos niveles de depresión y ansiedad.
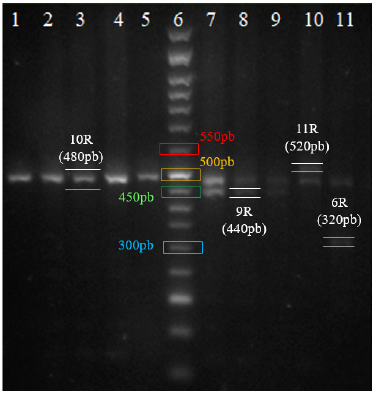
Fuente: autores.
Figura 1 Visualización de amplificación de polimorfismos de VTNR en gel de agarosa 2 %. De izquierda a derecha, en el pozo 6 se encuentra el marcador de peso (50 pb DNA Ladder). Desde el pozo 1 hasta el 5 se observan muestras con genotipo homocigotos 10 R (480 bp); desde el pozo 7 hasta el 9 se exhiben individuos heterocigotos con repeticiones 10R (480 bp) - 9 R (440 bp). En el pozo 10 y 11 se encuentran muestras heterocigotas con genotipo 11 R (520 bp) - 10 R, y 10 R - 6 R(320 bp) respectivamente.
La aproximación metodológica permitió identificar después de la electroforesis los distintos VNTRs de DAT1 en todas las muestras analizadas (ver figura 1), sólo se repitió la amplificación y electroforesis en dos muestras. No se encontraron diferencias en las frecuencias alélicas (p = 0,995) y genotípicas (p = 0,998) de los polimorfismos VTNR de la región UTR 3' del gen DAT1 según el sexo (ver Tabla 1). En la muestra de los 62 sujetos se logró identificar cuatro alelos, siendo los alelos de 10R y 9R los más frecuentes, y los alelos 11R y 6R, variantes raras en la población estudiada. En la figura 2 se observa que los genotipos más comunes fueron el 10R/10R, el 9R/10R y el 9R/9R (ver Figura 2). El análisis de la distribución de los genotipos VNTR del gen DAT1 mostró que la población se encontraba en equilibrio Hardy-Weinberg (p=0,844).
Tabla 1 Características generales, frecuencias alélicas y genotípicas de VNTR en el UTR 3' del gen DAT1 de los 62 sujetos seleccionados en el estudio.
Fuente: autores
De una muestra de 62 estudiantes universitarios con bajos niveles de depresión y ansiedad, se identificaron cuatro alelos VNTR para DAT1, los alelos más frecuentes fueron 10R y 9R, mientras que los VNTR 11R y 6R fueron poco frecuentes. Los alelos se encontraron en equilibrio Hardy-Weinberg (p=0,844). No se encontraron diferencias significativas en las frecuencias alélicas y genotípicas según el sexo (p=0,995 y p=0,998 respectivamente).
Discusión
Este estudio reporta por primera vez la frecuencia de alelos y genotipos tipo VNTRs de 40 bp en la región UTR 3' del gen DAT1 en una muestra de estudiantes universitarios con bajos niveles de depresión y ansiedad de una universidad privada de Cali.
Los alelos 10R y 9R fueron los más frecuentes en los sujetos estudiados, resultado similar al reportado en otras poblaciones adultas latinoamericanas como México (10R: 0,85; 9R: 0,15)5, Chile (10R: 0,74; 9R: 0,23)27, Brasil (10R: 0,70; 9R: 0,26)28 y en niños del centro de Colombia (10R: 0,78; 9R: 0,19)29. En contraste, la frecuencia del alelo 10R en la población de estudio fue menor a lo reportado en poblaciones indígenas colombianas, en las cuales la frecuencia del alelo 10R oscila entre 0,95 y 14, lo cual podría indicar que la muestra estudiada presenta una mayor variabilidad genética del VNTR DAT-1 asociada con el mestizaje de la población de origen.
Los alelos que presentaron baja frecuencia (0,01) en los sujetos estudiados fueron el de 6R y 11R. La presencia del alelo 11R se ha reportado con una frecuencia de 0,01 en población adulta sana de Rusia4, pacientes italianos con esquizofrenia30 y niños colombianos29 e ingleses31 con déficit de atención. Respecto al alelo 6R, este se ha registrado en Oriente Medio con una frecuencia de 0,0185. En todas las poblaciones reportadas al igual que en nuestro estudio, se encontró que las frecuencias genotípicas de los VNTR de DAT1 se encuentran en un equilibrio Hardy Weinberg, es decir, que la variación de esta región es constante de una generación a la siguiente gracias a un apareamiento aleatorio32.
Las frecuencias para los alelos 10R (0,75) y 9R (0,23) obtenidas en nuestra población de jóvenes con bajos niveles de depresión y ansiedad son muy similares a las encontradas en pacientes alemanes (n=190, edad=46 años) con síntomas depresivos, en donde los pacientes que portaban el alelo 10R (0,73) tuvieron una mejor respuesta (52 %) a tratamientos farmacológicos con antidepresivos que los pacientes portadores del genotipo 9R/9R (35 %)9. Asimismo, en una muestra de hombres rusos (n=657, edad=25-64 años) con antecedentes de ansiedad y depresión se observó que el alelo 10R de DAT1 (0.74) fue más frecuente entre hombres con un nivel severo de ansiedad, mientras que el alelo 9R (0,22) fue significativamente mayor en el grupo de hombres con depresión y agotamiento vital, sugiriendo que la presencia de estos alelos aumenta la posibilidad de presentar estos trastornos8. En contraste, Saung et al. (2014)14 no encontraron asociación entre el VNTR de 9R y 10R con el trastorno generalizado de ansiedad y depresión en pacientes que fueron tratados con el antidepresivo venlafaxina (n=156, edad=18-20 años). Estos resultados divergentes han llevado a proponer que los VNTR de la región UTR 3' de DAT1 no son los únicos determinantes de la expresión de este transportador de dopamina. Otras variantes en esta región génica como haplotipos con variantes tipo SNP ubicadas en el promotor y los intrones 1, 9 y 14 podrían estar contribuyendo al desarrollo de estas patologías psiquiátricas33.
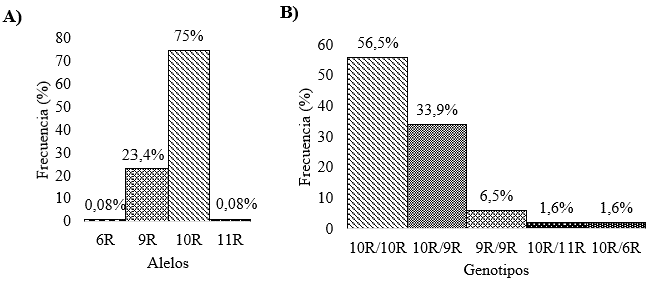
Fuente: autores
Figura 2 Distribución de los VNTR UTR 3' del gen DAT1 en los 62 sujetos estudiados. A) Frecuencias de los cuatro alelos encontrados, siendo el más frecuente el alelo de 10 repeticiones. B) Frecuencias de los genotipos observados, destacando el genotipo homocigoto 10R y heterocigoto con el alelo 9R.
La etiología de los trastornos psiquiátricos como la depresión y la ansiedad es compleja y se deben tener en cuenta diferentes factores ambientales y genéticos que pueden determinar su presencia en la población34, el VNTR del gen DAT1 es solo una de las variantes genéticas descritas (DRD4, BDNF, ACE, SERT,HTR2A, MTHFR,SLC6A4, TH, MAOA, FKBP5, CRHR1, COMT y CREB1)34-37 que se han asociado con el trastorno de depresión y ansiedad. Por lo tanto, el efecto de los polimorfismos de este gen debe ser considerado junto con otras variantes (VNTR o SNP) localizadas en regiones codificantes y reguladoras de genes relacionados con el sistema dopaminérgico.
Este estudio tuvo como limitaciones un tamaño de muestra pequeño, lo que puede afectar la precisión de la estimación de las frecuencias alélicas. Asimismo, dado que la muestra fueron estudiantes universitarios, puede que no sea representativa de la ciudad de Cali. Además, no se contó con un grupo control que presentara síntomas de depresión y ansiedad. Por otro lado, este estudio, aunque fue transversal, no tuvo seguimiento de los niveles de depresión y ansiedad de los participantes, así como tampoco consideró los antecedentes psiquiátricos o una evaluación por psiquiatría o neuropsicología para confirmar los resultados en las escalas de Beck, las cuales se aplicaron como autopercepción de síntomas de ansiedad y depresión. No obstante, como fortaleza, este es el primer estudio en reportar frecuencias de los VNTR del gen SLC63A, en una muestra universitaria de Cali.
Conclusiones
En la muestra estudiada se encontró que los VNTR de la región UTR 3' de DAT1, en estudiantes universitarios con bajos niveles de depresión y ansiedad de Cali, presentan frecuencias similares a las reportadas en otras poblaciones latinoamericanas y poblaciones con síntomas de ansiedad y depresión.
Si bien las variantes alélicas más frecuentes fueron los alelos 9R y 10R, los cuales se han asociado con trastornos de depresión y ansiedad en otras poblaciones, aún no es muy clara la participación de estas variantes en el desarrollo de estos trastornos. Por otro lado, las variantes menos frecuentes fueron las de 6R y 11R, alelos que pueden estar en nuestra muestra producto del mestizaje con poblaciones europeas y asiáticas. Las frecuencias reportadas podrían ser de utilidad para diseñar futuros estudios que investiguen asociaciones genéticas de los VNTR UTR3' de DAT1 con diferentes condiciones relacionadas con el sistema dopaminérgico en población universitaria colombiana.

