











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIn the urban environment; soils, microclimate, and other environmental conditions are often hostile to trees. Many urban microhabitats are not favorable for their growth and development, which frequently produces damage to the crown, branches, bark, and roots. Factors such as soil impermeability, hardness, and chemical composition, low nutrient and water inputs, the effect of contamination (high emissions of SO2, NOx, PM 2.5, PM 10, acid rain, among others), and thermal stress induced by urban heat islands cause physiological imbalance, by increasing the susceptibility to the attack of pests and diseases, and finally the tree’s death (Jochner and Menzel, 2015).
Because of the restrictions imposed by the urban environment, many species do not survive or grow well in cities; features such as reduced planting space, high tree costs, low availability of planting material in local nurseries, and current societal needs -e.g., trees without allergenic pollen- reduce the number of potential species to be used in future plantations. The stress of trees in the urban environment affects their populations in two ways: firstly, the increase in mortality, which raises the need for subsequent removals and replacements; and secondly, and perhaps more important, the removal and replacement of trees tend to generate an oversimplification of the urban ecosystem (Conway and Vander Vecht, 2015).
From the management perspective, the main negative effect of the dominance of one or a few species in urban forests is the increased probability of death due to outbreaks and attacks of pathogens and insects, which may result in the devastation of whole areas in one single event (Tomlinson et al., 2015). Therefore, species diversity has been considered a good alternative to increase the resilience of the urban forests against the attacks of insects and diseases. However, a high diversity of species brings difficulties of management since it supposes a greater range of requirements and potential phytosanitary problems (e.g., pruning time, nutritional demands, pests, and diseases).
The health of urban trees is essential for them to provide the desired ecological and economic benefits, which is one of the most significant challenges for managers of the urban forest. It has been found that the application of systemic insecticides and fungicides on trees affected by phytosanitary problems of subtropical species (elm, ash, among others) generates benefits in the short and medium term (Dal Maso et al., 2014; Mercader et al., 2015). Likewise, preventive actions such as sanitary pruning and fertilization are important for the management of trees since they reduce the stress generated by the urban environment (Fini et al., 2015).
Among the most common active components in insecticides used for phytosanitary management of urban trees are imidacloprid, bifenthrin, and thiacloprid, as they act selectively on the central nervous system of insects and present a low risk for non-target organisms such as mammals (Poland et al., 2006; Jeschke and Nauen, 2008). Although these substances have been applied in different parts of the tree, the application in the stem through injections has become one of the most used methods because it generates the minimum risk of exposure and release of their active components to the environment. The insecticide applied in the injection is transported to the insect feed site through the xylem sap (Mota-Sanchez et al., 2009).
Few studies have focused on tree management and health in Latin American cities (Escobedo et al., 2006 in Chile; Tavares et al., 2013 in Brazil; Restrepo et al., 2015 in Colombia). These studies have evidenced a deteriorating trend in the urban forests of Latin America associated with the growing population and the expansion activities derived (i.e., Industrial agglomerations, slum growth, increased automobile fleet), as well as pests and diseases that attack plant species when they are under persistent stress situations such as those prevailing in the urban environment. However, few studies have focused on the phytosanitary diagnosis and management strategies of urban trees in Latin America (Sepúlveda Cano et al., 2009; Rodrigues et al., 2014; Zaragoza et al., 2014), so the knowledge about optimal protection and remediation measures for urban forests in the cities of this part of the planet is limited.
The urban trees in the Metropolitan Area of the Aburrá Valley (Colombia) have been presenting phytosanitary problems and symptoms of deterioration and dieback for at least the last decade. The affected individuals have shown the presence of several species of insects and fungi, which could be responsible; however, no systematic search for the causal agents of this phenomenon has been done yet. As the first step in this search, a field experiment was performed to evaluate the effect of applying systemic insecticides and fungicides on the health of the crown of 15 tree species. The hypothesis is that the deterioration of tree crowns is generated by the action of insects and fungi, which is favored by the stress of trees in the urban environment. The specific objectives of the study are 1) to determine the effectiveness of treatments to reverse the symptoms associated with the deterioration of the aerial part of the treated individuals; 2) to identify some factors that could affect the temporal behavior of the individuals evaluated (during the experimental period, 2013 - 2015). It is expected that the results obtained will contribute to improving the knowledge of the management and health of trees in tropical cities.
MATERIALS AND METHODS
Study area
This study was carried out in the urban area of four municipalities (Itagüí, Medellín, Sabaneta, and Envigado), belonging to the Metropolitan Area of the Aburrá Valley, the urban center with the second largest concentration of population in Colombia, with an urban area of 345.6 km2. The altitude varies between 1400 and 1600 m.a.s.l., with an average annual temperature between 22 and 25 °C, and an average rainfall of 2500 mm year-1 (AMVA and CONCOL, 2007). According to data collected from the weather station Olaya Herrera Airport during the experimental period (2013-2015), the rainfall is bimodal, with two humid periods in April-May and October-November, when the lowest average monthly temperatures occurred; and two dry periods (in June-July and December-January), with the highest temperatures (Figure 1).

Figure 1 Monthly precipitation (mm) and average temperature (°C) at Olaya Herrera Station 2013 - 2015. (IDEAM, 2015).
Definition of the study variable and species selection
The variable of interest in this study was called progressive deterioration (PD), a phenomenon associated with symptoms of sudden death, wilting, branch rot, and downward drying in trees of the urban areas of the Aburrá Valley (Restrepo et al., 2015). Symptoms start with the drying of upper branches, continuing with the intermediate branches, and ending with general deterioration and the death of the tree. The evaluation methodology consisted of recording the absence or presence of PD in three levels of severity: low (less than 20% of the crown affected), medium (affectation between 20 and 60% of the crown), and high (more than 60% of the crown affected).
It has been found that some species are more susceptible to exhibit the PD symptoms (Restrepo et al., 2015; UNAL and AMVA, 2015); for this study 15 of them were selected, which also were abundant and widely distributed in the area (Table 1). It was evaluated the effect of a combined treatment, consisting of the application of systemic insecticides and fungicides plus a general treatment of sanitary pruning and fertilization as phytosanitary protection measures to reverse the PD affectation.
Table 1 List of species studied, value of incidence of progressive deterioration (PD) in years 2012 and 2014 (Restrepo et al., 2015; UNAL and AMVA, 2015, respectively), and wood density.

The treatments evaluated were (Table 2):
1. . Negative control: healthy trees, which did not receive any treatment.
2. . Positive control: trees affected by PD that did not receive any treatment.
3. . Treatment 1 (T1): trees affected by PD and free of soil pathogens. This group received the application of systemic insecticide and beneficial fungi. The treatment consisted in the application in the base of the tree (in four equidistant points) of an intracambium injection of 20 cm3 of systemic insecticide (Bifenthrin (50 g L-1) + Imidacloprid (250 g L-1)) and the application to the soil of 20 g of a mixture of antagonistic fungi (Trichoderma harzianum, T. koningii, and T. viridae) dissolved in 10 L of water, as a preventive measure to inhibit the growth and development of soil phytopathogenic fungi.
4. . Treatment 2 (T2): trees affected with PD and by soil pathogenic fungi. This group received the application of systemic insecticide and fungicide. The treatment consisted of the application in the base of the tree (on four equidistant points) of an intracambium injection of 20 cm3 of systemic insecticide (Bifenthrin (50 g L-1) + Imidacloprid (250 g L-1)) plus 20 cm3 of a systemic fungicide (Carbendazim 500 g L-1) dissolved in water to a concentration of 0.6 cm3 L-1.

To enhance the effect of treatments, individuals from T1 and T2 received three sanitary pruning of affected branches (with 50% or more of their length without leaves and with evidence of decay) and three soil fertilization (circular at 50 cm from the tree base) with a mixture of 400 g of triple 15 (N: 15%, P: 15%, K: 15%) + 100 g of a fertilizer with major and minor nutrients (N: 8%, P: 5%, Ca: 18%, Mg: 6%, S: 1.6%, B: 1%, Cu: 0.75%, Mo: 0.005%, and Zn: 2.5%).
A total of 188 adult individuals from 15 species were sampled, which were evaluated five times including the baseline measurement. Treatments along with pruning and fertilization were applied three times, in the baseline (July - August 2013), in the second (October - November 2013) and fifth measurement (March - April 2015).
Statistical analysis
Contingency tables were built to evaluate the possible relationships between the categorical variables and the recovery of individuals between the baseline and the fifth measurement. The recovery was defined using a binary categorical variable, with 1 for recovery and 0 for no recovery of the individual; that is, if the individual went from any category of affectation (i.e., low, medium or high) to the absence of PD, this variable was equal to one.
Given that the nature of this study is longitudinal, and therefore the response has both a contribution of repeated measures per individual and a non-continuous response (binomial in this case), the construction of models to explain the presence of PD during the evaluation time was performed using generalized linear mixed models (GLMM) (Fitzmaurice et al., 2011). The model was based on the explanatory approach outlined by Schwartz and Lellouch (2009), and its purpose was to define the effect of how the tree was treated during the experimental period considering the variations that might occur (i.e., incomplete application of the treatment). The so-called full cases methodology was used for handling of dropouts, given the low percentage of dead individuals and/or cut trees. The model was fitted using the PROC GLIMMIX of the SAS® V9.4 software.
The adjusted generalized mixed model was:

Where Y ij was the response variable, presence of PD measured in the i -th subject at time j; i=1,2,..., N denotes the subjects and j=1,2,..., ni denotes the observations for the i-th subject at time j (longitudinal observations). Y ij is a binary response taking the value of 1 when a specific tree shows PD (any of the three categories above mentioned), and 0 for no presence of PD. In both models, the link function to connect the linear predictor and the response was the logit. b 0 corresponds to a random intercept which was specified for each subject under study. By doing so, it is believed that the model can capture extra variability that a model based only on fixed effects cannot capture. The vector of parameters b=(b1, b2,..., b6), includes the regression coefficients of the fixed effects of the model; b0 is the population intercept of the model. Among the fixed effects,t ij is the time of evaluation of the subject i at time j and was recorded as follows:
Baseline or first time: July-August 2013
Second time: October-November 2013
Third time: May-June 2014
Fourth time: March-April 2015
Fifth time: December 2015
spi corresponds to the i -th subject of the species (Table 1). ISSi,j=1 is the initial severity state of PD of the i-th subject at the baseline. T1ij and T2ij correspond to the dichotomous variables Treatment 1 and Treatment 2 that were applied to the i-th subject (which takes values according to the time j ), and Fij is a dichotomous variable taking the value of 1 if fertilization on the i-th subject was applied at time j. This differentiation was done because it was not possible to apply treatments as planned in some individuals.
The so-called Residual Pseudo-Likelihood (RSPL) technique was used to obtain the estimates of both fixed and random effects. This technique uses a subject-specific expansion through the Taylor series. The variance-covariance matrix of fixed effects was obtained using the empirical estimator (sandwich estimator) to obtain a robust analysis taking into account the lack of specification of the covariance structure. The structure called unstructured was selected to calculate the variance-covariance matrix of the random effects. Paired comparisons between the levels of the variables that were significant in the models were performed. In all comparisons, P<0.05 was considered statistically significant.
The model building had several stages. At each stage the inclusion of variables was evaluated; among such variables are planting site (hard floor, tree-grate, and green zone), traffic flow of the site (high, medium and low), and dasometric variables of trees (diameter and height). However, none of these variables were significant and for that reason were not included in the model presented above.
Finally, the estimated probability of PD at the five evaluation times for each species was related to the wood density of the species (kg m-3). Also, with the individual probability of suffering PD, profile plots per species were built to evaluate the trend of PD over time.
RESULTS AND DISCUSSION
From the 188 individuals evaluated, 13 had incomplete responses because death and cut (eight and five, respectively) and therefore they were not analyzed. Significant differences between species in terms of its recovery between the baseline and the fifth measurement (Chi-squared=34.21, df=14, P<0.05) were found. In contrast, recovery showed no significant difference neither between planting sites (Chi-squared=5.4959, df=2, P>0.05) nor the traffic flow of the site (Chi-squared =2.1185, df=2, P>0.05) (Table 3).
Table 3 Contingency table for the variables planting site and vehicular traffic. (1) There was recovery; (0) there was no recovery (α=0.05). The Recovery columns show the number of individuals; the Comparison column shows the probability associated with the hypothesis test of equality of each comparison.

The positive coefficients estimated in the model indicate that the respective level of the categorical variable increased the presence of PD, while a negative coefficient indicates a decrease in the probability of PD. In this way, it was found that the probability of PD decreased during the study period (time), being lower in the last measurement (Table 4), presenting a temporal pattern of decreasing, which it was found to be general. Thus, differences in the presence of PD at the evaluation times (time) were higher, and statistically significant, as they were more spaced in time, being the baseline and the 2nd measurement the ones that showed a greater difference in comparison to the other measurements. The last two measurements did not show differences concerning the probability of having PD (between 0.18 and 0.12, t=1.69, df=692, P>0.05) (Figure 2).


Figure 2 Least squares means (LSM) of the paired differences of evaluation times (left) and the initial state of severity (ISS, right) (α=0.05, Bonferroni adjustment). Dark lines: non-significant, clear lines: significant. The reference line (dotted line of 45°) shows a value of 0 for the confidence interval; as a consequence, comparisons whose confidence intervals cover zero through the reference line of 45 ° are not significant; on the other side, if the difference in the LSM is significant, the solid line for the confidence interval is completely above or below the dotted line of equality (High, 2015).
On the other hand, the presence of PD, as a function of the initial state of severity (ISS), was not significantly different when the individuals were truly affected (i.e., when they had a low, medium or high affectation at baseline); however, these levels showed differences with respect to the unaffected trees, called “negative controls” (Figure 2).
The applied treatments (treatment 1, and treatment 2; plus pruning and fertilization) did not show any influence at the individual level since trees treated showed the same behavior as those untreated (positive control) (Table 4). Contrary to these results, intracambium injections have been effective as a remedial and sanitary measure in urban trees of subtropical and temperate species such as Ulmus spp. (Poland et al., 2006), Fraxinus spp. (Mota-Sanchez et al., 2009), and Tsuga spp. (Doccola et al., 2007). However, these results probably are not comparable with this study, since there are marked differences with the tropical climatic conditions and species composition (both of the pathogen and the host) of the present investigation; besides, most of these studies assessed individual plague-host relationships, which facilitates dose evaluation, mechanisms of application, and effectiveness. Similar studies in this part of the tropics have not been reported (in terms of species and application of intracambium injections) as a remedial measure against the progressive deterioration of standing trees.
The applied treatments assumed that the deterioration of tree crowns is caused by insects and fungi, which in turn is favored by the stress of trees in the urban environment. However, these results did not support this hypothesis. An alternative explanation emerges from a close look at the climate data during this study. In the period of selection of the individuals studied (July - August 2013), the climate was dry and hot, the highest temperatures recorded for that year (Figure 1). Before this period, there was a wet season whose maximum rainfall occurred in May, with a drastic decrease of rainfall by almost 50% in June and an even more drastic decrease in July (34 mm precipitation in that month), which is atypical for the mild weather conditions of the region. This decrease in the availability of water probably produced stress in the individuals evaluated and consequently, the appearance of symptoms of crown deterioration. However, tree recovery occurred as precipitation increased and temperature declined in subsequent months, suggesting a positive effect of increased water availability; which confirms the temporal pattern of PD. Therefore, although the attack of insects and fungi occurred, they appeared opportunistically, probably because of previous environmental stress; therefore, trees recovered as the stress diminished or disappeared.
Garrett et al. (2006) propose that there are climatic conditions that favor the pathogen and disfavor the host, which increases the potential for damage to trees. As a consequence, the pathogen-host relationship that occurs in some species of the Aburrá Valley probably obeys a seasonal pattern favored by the bimodal behavior of the climate in this zone, which causes stress in the trees during the times of greater radiation and temperature and lower precipitation, which benefits the proliferation of pathogens. Likewise, its harmful effect diminishes once the rains appear, when trees regain their vigor. This seasonal behavior of stressors would favor the existence of a physiological response called “acclimatization” (Niinemets, 2010), which would explain the recovery of most individuals, both treated and untreated.
At the species level, it was found that some of them were more susceptible than others. Species like A. cunninghamiana, S. malaccense, and C. pluviosa had a lower probability of occurrence of PD in contrast with S. campanulata, F. benjamina, and T. catappa (Table 4, Figure 3). Previous studies (Restrepo et al., 2015; UNAL and AMVA, 2015) for the same species in the study area reported a high correlation between the probability of PD occurrence and the incidence, confirming that the susceptibility to PD also depends on the species. Species such as S. campanulata, F. benjamina, and T. catappa have shown high susceptibility to the attack of sucking insects, borers, and defoliators in urban areas; which act as deleterious phytosanitary agents (Bito, 2007; Held and Boyd, 2008; Begoude et al., 2010; Arthurs et al., 2011; Maruyama et al., 2012; Tavares et al., 2013; Tavares et al., 2014; Herrera Isla et al., 2015; Lima et al., 2016) and explains in part the behavior shown by these species in this study.

Figure 3 Confidence intervals of the probability of PD by species. Different letters indicate significant differences. Sm: S. malaccense, Ac: A. cunninghamiana, Cp: C. pluviosa, Fc: F. chinensis, Rr: R. regia, Hc: H. chrysantha, Ex: E. japonica, Pd: P. dulce, Ls: L. speciosa, Lp: L. punicifolia, Jm: J. mimosifolia, Bp: B. picta, Tc: T. catappa, Fb: F. benjamina, Sc: S. campanulata. Values with the same letter are not significantly different among them (ls-means PROC GLIMMIX SAS® 9.4).
The estimated probabilities of PD during the study period were different for the evaluated subjects (Figure 4). The individual profiles by species showed a general trend in which a group of individuals maintained the very low probability of PD throughout the study; these were the subjects selected as positive controls, and their initial assessment of severity equaled to 0 (Figure 4). It was also noticed that subjects of A. cunninghamiana, C. pluviosa, and S. malaccense, affected at baseline, rapidly decreased the probability of PD and remained in that situation during the rest of the study. This trend was not observed in species such as B. picta, F. benjamina, S. campanulate, and T. catappa, whose probability of PD was high most of the time. Other species, such as E. japonica, F. chinensis, H. chrysantha, J. mimosifolia, L. punicifolia, L. speciosa, P. dulce, and R. regia did not show a clear trend in terms of the probability of suffering PD, and consequently, it was not possible to build a cluster with their profiles.

Figure 4 Individual probability profiles of PD during the study period by species according to the model. Vertical discontinuous lines show the times of treatment application. The color of the line indicates the initial state of affectation, gray: affected, black: healthy.
Tree crown health may be impaired by multiple causes, including water deficit, attack of insects, fungi, viruses, phytoplasmas, nutritional deficiencies (and the interaction among them); therefore, the diagnosis and treatment are complex, especially if one considers that different attacks may exhibit similar symptoms in different species. It has been reported that not only species do differ widely in their tolerance to different environmental factors (Philip and Azlin, 2005, Percival et al., 2006), but also that the relation individual-species has a great effect on the longevity and survival of trees. Thus, a species can present individuals with high capacity for recovery (or acclimatization), as well as individuals that do not achieve a satisfactory recovery of their carbon stores after suffering successive stresses, causing irreversible physiological alterations (i.e., damage to photosystem II that inhibits photosynthesis, limited hydraulic conductivity, alteration in gas exchange, etc.), which may explain why some individuals did not show symptoms of recovery during the experimental period.
Finally, by species, it was observed that wood density showed a high correlation (nonlinear) with the probability of occurrence of PD (Spearman correlation coefficient=-0.70); in other words, such probability increased in species with low wood density. Likewise, the Kruskal-Wallis test showed that the classes of density were significantly different with respect to their probability of PD (P<0.05); thus, the pairwise comparisons of these classes showed that softwood species had significantly higher probability of having symptoms of PD (Figure 5) than those with medium and high density (P<0.05).
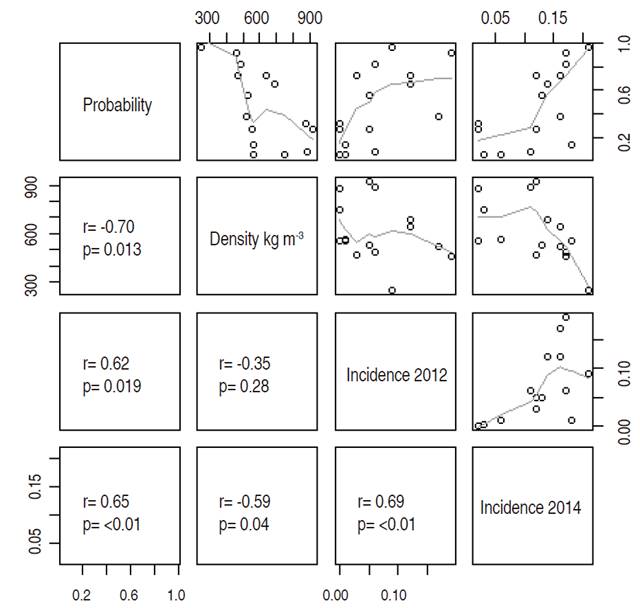
Figure 5 Matrix of non-linear correlation (Spearman) of the probability of PD (found in this study), wood density, the incidence in 2012 and incidence in 2014 per species.
It has been proposed that wood density is positively correlated with resistance to rot, since the high density may constitute a potential mechanism to reduce the risk of wounds (by falling branches) and the subsequent pathogen attack (Chave et al., 2009). Larjavaara and Muller-Landau (2010) propose that high-density woods rot slowlier since there is less surface on which spores can land, and more solid structures between the bark and pith, which increase resistance to penetration. In the same way Hacke et al. (2001), present that the size of the xylem vessels is related to wood density, as denser wood species generally have smaller vessels; this, in turn, leads to lower rates of water transport, but it represents greater resistance to xylem cavitation during droughts.
Individuals that remained unaffected throughout the study suggest high resistance to PD in most species analyzed. It is necessary to inquire about the physiological and genetic conditions that favor such phytosanitary status of this group of trees, which will help to improve the understanding of this phenomenon and its management. Also, the results suggest that effective treatments to maintain or improve urban tree health in subtropical areas are not necessarily applicable to the conditions of tropical cities.
CONCLUSION
The results obtained in the present study suggest that PD is a highly dynamic phenomenon in time that seems determined mainly by the climatic variability, which is facilitated by genetic characteristics of trees, such as wood density, and their physiological condition. The action of insects and pathogens seems to be opportunistic once trees are affected.

