











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Moringa oleifera Lamarck (MO) belongs to the Moringaceae family. Although this plant is originally from India, it is well distributed around the tropical zones of the planet. It is wildly recognized as a drought-resistant species with many applications in animal and human nutrition. MO can grow in many different environmental conditions and it has been reported to produce up to 580 t ha-1 per year of fresh sprouts. For these reasons, The Food and Agriculture Organization of the United Nations (FAO) recommends Moringa as a potential crop (FAO, 2019). There are no official reports of its growth in Ecuador, however, there are private initiatives such as Ecuamoringa, which manages a wide national market.
MO leaves have important nutritive qualities among perennial vegetables including their protein content between 24-29 g 100 g-1, and important amounts of vitamins A and C, and minerals such as Ca, Fe and P. MO protein contains all the essential amino acids in important concentrations and a good amino acid profile reaching 72.4% (Alain et al., 2016), despite the high amount of leaf required to obtain them, and the low protein digestibility-corrected amino acid score (PDCAAS) 41.42% related to antinutritional factors in the plant. Amino acids are essential for the human body, being used as synthesis precursors of proteins, peptides and low molecular weight molecules. The absence of these compounds causes sickness as kwashiorkor and marasmus, which are common in developing countries (Wu, 2016).
Finding new protein sources is a key strategy to face the fast population growth and the environmental problems of big-scale cattle growing. Although in developed countries animal meat and its derivatives are still the main sources of protein, investigation of alternative protein sources is growing in popularity (Haque and Varshney, 2015).
Peptides of MO have demonstrated antioxidant capacity and the ability to inhibit the angiotensin-converting enzyme (ACE). These effects depend on the amino acid type and composition of the peptides formed in the stomach through the synergism of internal peptide enzymes (Saucedo-Pompa et al., 2018).
Moringa has applications in traditional medicine to treat diabetes. This quality has been principally attributed to its secondary metabolites. Moreover, different studies have demonstrated the pharmacologic properties of the foliar protein of different vegetables. Specifically, Paula et al. (2017) have proved the possibility to reduce the blood glucose level up to 66.4% by administrating 500 mg kg-1 of an aqueous extract of MO leaves; as well as the significant reduction off malonaldehyde (MDA) synthesized by the body in the presence of high concentrations of reactive oxygen species (ROS), which cause oxidative stress and mutagenic problems (Nishikawa et al., 2000).
Despite the health benefits obtained from MO consumption, direct consumption of high amounts of vegetable leaves could cause different disadvantages, especially in nutritional terms due to their antinutritional components and fiber. Neither trypsin nor amylase inhibitors have been detected in MO leaves, but important concentrations of phenols, tannins, saponins and phytates decrease nutrients’ bioavailability in this crop (Makkar and Becker, 1997).
Furthermore, MO has a bitter and astringent flavor caused by flavonoids such as catechins and glucosinolates, specifically 4-(rhamnopiranosiloxi) benzil glucosinolate and monoacetil (rhamnopiranosiloxi) benzyl glucosinolate; this latter group is transformed to isothiocyanates by the myrosinase enzyme in the mastication process, incrementing the pungent and spicy flavor, so food likeness decreases (Doerr et al., 2009). In this context, it is valuable to isolate a leaf protein to use it as a supplement or raw material in the development of nutritious food products and further pharmacological applications.
Solubilization of protein is the first step in isolating protein from other leaf compounds. There are many different methods to separate protein such as the use of organic solvents, aqueous extraction and enzymes. The most used method is the aqueous extraction with addition of salts or changing the pH of the solvent, due to its low cost (Tan et al., 2011). Alkaline extraction based on Osborne (1924) protocol is commonly used in the industry, and the addition of different salts of Na+ and Ca+ has been reported beneficial for protein extraction in some food materials (Martinez-Maqueda et al., 2013). There is not much information about the interaction of salt concentration and alkaline pH in vegetable protein extraction and solubilization.
MO has all the protein fractions described by Osborne and Voogt (1978): water-soluble albumins, saline soluble globulins, 70-90% ethanol-soluble prolamins and acid alkali-soluble glutelins (Teixeira et al., 2014), which means that the alkaline and salt extraction could be used in order to obtain more protein while avoiding organic solvents such as ethanol because of the high economic and environmental costs. Furthermore, the leaf protein must be separated from the other compounds in the extracted solution. A good strategy is the use of isoelectric precipitation, as well as the use of temperature to reduce protein solubility.
As a strategy to deal with the rapid population growth and the environmental problems generated by large-scale livestock farming, it is important to seek new sources of protein. Therefore, this study aimed to optimize the conditions of the Moringa oleifera Lam. leaf protein extraction and precipitation processes, as a green and eco-innovative alternative to obtain protein, concentrates with nutritional quality by reducing the effect of other components present in the matrix food and that can compromise their quality.
MATERIALS AND METHODS
Raw material
The leaves of Moringa oleifera Lam. were obtained from Ecuamoringa S.A., an Ecuadorian company, headquartered in Guayaquil that operates mainly in the agricultural area and stands out as a producer of MO. The drying process was performed at 35 °C until obtaining constant weight. The material was grounded and standardized using a mesh sieve of 0.25 mm (Teixeira et al., 2014).
Reagents
Hydrochloric acid (PubChem CID: 313; 37%, MERCK); Sodium hydroxide (PubChem CID: 14798, Fisher Scientific); Kjeldahl catalyzer (Cu-Se), (Scharlau); sulfuric acid (PubChem CID: 1118, MERCK; Antifoam (Sodium sulfate 97% and silicone 3%, Velp Scientific); Boric acid (PubChem CID: 7628, Loba Chemie); Bradford reagent and Bovine Serum Albumin (BSA) (PubChem CID: 16132389, Sigma-Aldrich). These reagents were imported from the United States (Fisher Scientific, Merck, Sigma-Aldrich and Velp Scientific), Spain (Scharlau) and India (Loba Chemie).
Protein extraction process
The methodology of Tan et al. (2011) was followed with modifications: 20±0.1 g of milled leaf were weighted with an electronic analytical balance (ML204, Mettler Toledo, Zurich Switzerland) with a deviation of ±0.1 mg scale in a 500 mL Erlenmeyer flask to finally add the water-NaCl solution in relation 1:10 w/v. The sample was homogenized for 5 min in VELP Scientifica stirring plate at 800 rpm. The pH was adjusted to 10, 11 and 12 with NaOH 1 N and measured with a Mettler Toledo model Seven Compact potentiometer according to the experimental design. The Erlenmeyer was placed in a shaking bath (Julabo SW22) at 25 °C at 200 rpm for 1 h. The pH was rectified each 10 min.
Experimental design
A Complete Randomized Design (CRD) was used with a factorial arrange of 32, resulting in the combination of pH (10, 11 y 12) and NaCl concentration (0, 0.25 and 0.5 M). The nine treatments E1 (pH 10, 0 M); E2 (pH 10, 0.25 M); E3 (pH 10, 0.5 M); E4 (pH 11, 0 M); E5 (pH 11, 0.25 M); E6 (pH 11, 0.5 M); E7 (pH 12, 0 M); E8 (pH 12, 0.25 M); E9 (pH 12, 0.5 M) were performed in triplicate obtaining 27 experimental units. The response variable was the protein extraction yield.
Protein analysis
Kjeldahl method .The total content of protein in the extract was determined by the Kjeldahl method (AOAC 991.22) (AOAC, 2019). The conversion factor used was 6.25 (Mbailao et al., 2014). Three temperature ramps were programed (140 °C for 15 min, 250 °C for 20 min and 420 °C for 40 min) in the protein VELP Scientific DK 6 digester to avoid overboiling of the sample. The distillation was performed in a VELP Scientific UDK 132 distillatory.
Bradford method. To use a less expensive and faster method for the quantification of protein in the extract, the Bradford method was investigated and compared with the Kjeldahl method. A Bradford test kit was used for this purpose (Sigma Aldrich). A calibration curve was constructed using BSA as standard. For it, 5 uL of the sample were mixed with 250 uL of the Bradford reactive and were incubated in a dark camera for 25 min. To measure the optical density inside the range of the calibration curve, each one of the protein samples was diluted in a ratio of 1/15, 1/20 and 1/25 with pure water. The optical density was measured in an Elisa MRX Microplate Reader from Dynex Technologies (Denkendorf, Germany) at 595 nm.
Protein extraction and precipitation yield
The comparison between the extracted protein and the original amount of protein in the leaf previous to the solubilization process was used to calculate the protein extraction yield. Also, the protein precipitation yield was calculated by comparing the remaining protein from the supernatant with the original protein concentration in the extract.
Relationship between protein analysis methods
The Pearson correlation coefficient determined the relationship between both methods. The Tukey test was used to compare the results of each treatment by different methods of analysis.
Precipitation process
The best treatment from the protein extraction process continued with the precipitation stage following the methodology of Serpa-Guerra et al. (2014) with modifications. From the extract, 1300 mL were divided into 100 mL aliquots. The pH was adjusted with a solution of HCl 1N following the experimental design and the temperature of the shaking bath configured to 100 rpm for 30 min.
The samples were put in 4 Falcon tubes of 50 mL to be centrifuged (Hermle Z206A) for 10 min at 5380 rpm to separate the supernatant from the protein (precipitate).
Experimental design
The treatments were analyzed by a CRD with factorial arrange 22 with the factors pH (4 and 5) and temperature (40 and 80 °C). Each treatment was performed twice and 6 central points were added to obtain 5 treatments: P1 (pH 4, 40 °C); P2 (pH 4, 80 °C); P3 (pH 5, 40 °C); P4 (pH 5, 80 °C); P5 (pH 4.5, 60 °C). The precipitation yield was used as a response variable.
RESULTS AND DISCUSSION
Protein extraction
The purpose of the first stage of this study was to determine the best combination of pH and NaCl concentration to obtain the maximum extraction yield from MO leaves. The original content of protein from the leaf was 26.96±0.24 g 100 g-1. Similar results were obtained by Olson et al. (2016) (27.3 g 100 g-1). This value was used to calculate the protein extraction yield. The calibration curve for the Bradford methodology had the regression equation y=0.2927X+0.4408 with a determination coefficient (R2) of 0.9954.
A significant difference was found between the treatments (P<0.05), as well as in the influence on the protein content of the pH, NaCl factors and their interaction by both analysis methods (Table 1).

According to Condo and Pazmiño (2015), the coefficient of variation (CV) in experiments carried out in the laboratory should be up to 5%, to show the reliability of the research. In the present study, both Kjeldahl and Bradford methods presented a CV of 3.23 and 3.9% respectively.
The medium pH is the decisive factor in the solubility of the protein. The net charge of the protein in certain pH depends on the pKa values of the ionized groups of the protein. There are three different possibilities for the isoelectric point (pI): the net charge is 0, the net charge is positive in pH values lower than the pI and values higher than pI generate negative charge. The solubility of the molecule depends on the molecule charge. When pH is equal to pI, the solubility is minimum, but in higher and lower pH values, the solubility forms a "U shape" distribution with higher solubility in alkaline pH (Zayas, 2012). In addition to the increase in protein solubility, the alkali hydrolyzes the protein bounded with polyphenols and polysaccharides decreasing their hydrophobic characteristic. Furthermore, the cellular wall is damaged by the NaOH creating cracks and increasing the diffusion rate (Zhang et al., 2015).
On the other hand, MO leaves have an important content of phytates that could be higher than in other legumes. Phytates bind to certain minerals such as Ca+ or Mg+, and proteins, forming insoluble complexes that reduce bioavailability. This effect could also be influenced by the inhibition of digestive enzymes entrapping them or chelation of their principal substrate in the organism: calcium. Through the extraction process in an alkaline medium, the pH can alter the phytate-protein complex and with sufficient amounts of Ca+ (present in high concentrations in MO), these molecules will precipitate and could be separated from the extract before the precipitation of proteins (Rham and Jost, 1979).
In addition, the saline concentration of the medium influences the ionic strength; however, the mechanism by which it influences the protein solubility is still unknown (Zayas, 2012). Salts interact with protein-charged groups decreasing the electrostatic attraction and improving the relationship between the molecule and the solvent. When the saline concentration reaches a determinant point, the water molecules are not able to support the burden of the ions and proteins, so the less soluble solute is precipitated (protein) (McQuarrie and Simon, 1997). Since the charged groups of protein that interact with the ions of the salts depend on the pH of the medium, the effect of the saline concentration is related to the pH as could be seen by the significant interaction between both factors (P<0.05) (Table 1).
Both treatments E4 (pH 11) and E7 (pH 12) without salt were statistically equal between them. The extraction yields were 40.4 and 38.79 g 100 g-1 by the Kjeldahl method and 38.11 and 40.28 g 100 g-1 by the Bradford method, respectively (Table 2). These results are similar to those reported by Zhang et al. (2015) in alkaline extraction from tea leaves (Camellia sinensis)and 41.5 g 100 g-1 obtained by Coldebella et al. (2013) in Manihot esculenta leaves with a similar methodology.

The protein extraction yield related to the increase in pH value is explained by the increase of the negatively charged groups of the protein that generate electrostatic forces, which avoid molecules from joining and precipitating (Hou et al., 2017).
Similarly, salts in low concentrations generally improve the solubility of the protein, stabilizing it through non-specific electrostatic interactions (Perez-Jimenez et al., 2004). Nevertheless, in the conditions of this experiment with high charged proteins due to alkaline pH, the salt arouses the electrostatic repulsion by masking the charges and reducing the solubility in consequence (Dahal and Schmit, 2018).
The Bradford method is used to analyze protein content as an alternative to the Kjeldahl method. It uses Coomassie Brilliant Blue (CBB) as the colorant, which creates a complex with protein by electrostatic interaction and Van der Waal forces. This method is especially used to measure soluble protein because it is fast and cost-efficient (Palada et al., 2007). It measures the increase in absorbance of the sample after the addition of the dye and the incubation for a specific time, but if the time exceeds, precipitation of the complex is possible.
The complex is insoluble from the beginning of its formation, and the accuracy of the results could be influenced by the structure of the protein to be analyzed, and its solubility (Marshall and Williams, 1992).
Despite the high Pearson correlation coefficient (0.886) from the range (-1 to +1), only the extraction yield of treatments E4 (pH 11, 0 M NaCl), E7 (pH 12, 0 M NaCl) and E8 (pH 12, 0.25 M NaCl) analyzed by the Kjeldahl method were statistically equal compared with Bradford (Table 2); nonetheless, the other treatments had different behavior depending on the analysis method (P<0.05). It is possible that due to the high pH and the low saline concentrations, the protein was more soluble in those treatments, consequently, the Bradford test was able to quantify the majority of the protein in the sample. However, at lower pH values and higher saline concentrations, the macro-molecule is less soluble and the protein-CBB complex could not be formed.
In contrast, the Kjeldahl method determines the total nitrogen content and converts it to the amount of protein by a conversion factor depending on food composition (Mbailao et al., 2014), so it is more trustworthy to analyze the protein content in this case. Also, the Bradford methodology has a higher CV than the Kjeldahl one due to the calibration curve of the first one being linear in a small range between 0 ug mL-1 to 2000 ug mL-1, so it requires successive dilutions in the sample and that increase the variability (Ernst and Zor, 2010).
As a result of the extraction stage, the treatments E4 (pH 11) and E7 (pH 12) had the best extraction yields (P<0.05). As E4 used less NaOH, it may prevent the damage in the protein from extreme basic mediums (Friedman, 2010). Thus, E4 followed the precipitation stage.
Protein precipitation
There was a significant difference between treatments, and the pH factor had a statistical influence on the precipitation yield (P<0.05) (Table 3). It could be explained because the isoelectric precipitation happens when the pH of the medium is equal to the pI of the protein generating a 0 net charge (Shaw et al., 2001).

The CV was 1.21%, which confirms its reliability (<5%) (Condo and Pazmiño, 2015). Table 3 shows that the curvature is not statistically significant (P>0.05), and that implies a lineal model without the need to add extra experimental points to study the quadratic effect (Gutierrez et al., 2008). The treatments P1 (pH 4, 40 °C), P2 (pH 4, 80 °C) and P5 (pH 4.5, 60 °C) were statistically the same and presented the highest precipitation yield (P<0.05) (Table 4). The influence of the pH in the reduction of the solubility is related to the achievement of the isoelectric point of the protein. In this condition, the protein-protein interactions increase due to the drastic decline in electrostatic forces and the lesser water interaction with these macromolecules (Santamaría-Fernández et al., 2019).
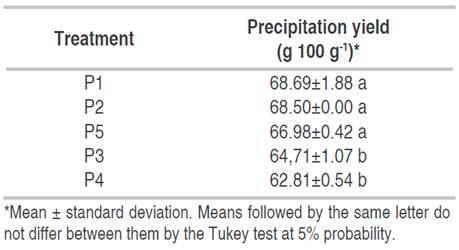
MO protein had similar behavior to other vegetable proteins, decreasing the solubility when the pH was near 4 (Kobbi et al., 2017). The precipitation yield was higher than that reported by Urribarri et al. (2004) (62.5 g 100 g-1), who used pH 4 and 50 ºC in Pennisetum purpureum Schum cv. Mott.
In general terms, temperature decreases the solubility of the protein due to conformation changes in the structure. The modification is irreversible, and it depends on the temperature and heating time. The solubility of the protein increases until reaching 40 °C and from that point, it starts to decrease. The influence of the temperature is related to the pH and the ionic strength of the medium (Zayas, 2012). This study showed that the pH changed the protein configuration, eliminating the repulsive forces and letting the greater amount of protein in the extract precipitate before it was exposed to the heat treatment, so the effect of temperature was not significant (Tables 3 and 4). Avoiding the use of temperatures while obtaining the best possible precipitation yield prevents the protein from losing heat-labile amino acids or damaging other of its functional properties (Belhadj-Slimen et al., 2016).
Considering the average of both stages (extraction and precipitation), the maximum obtained yield was 26.93±0.22 g 100 g-1 from the original content of the leaf protein, which represents about 7.26±0.19 g of isolated protein from 100 g of the leaf. This result is similar to those obtained by Nissinen et al. (2008) who recovered 24-26 g 100 g-1 of Phleum pratense leaf protein and to the Edwards et al. (1975) results, who reported 26.1 g 100 g-1 in Medicago sativa. As well as the results from Chiesa and Gnansounou (2011) with alfalfa (Medicago sativa) protein (8.53 g 100 g-1 from dry leaf). Moreover, Edwards et al. (1975) obtained in the same crop a yield of 8.7 g 100 g-1 from the dry leaf.
CONCLUSIONS
The best condition for Moringa oleifera Lam. leaf protein extraction was pH 11 or 12 without NaCl, which allowed obtaining the highest yield with a significant difference concerning the other treatments. NaCl negatively influenced the solubility of proteins in an alkaline medium. Ranges of pH between 4 and 4.5 are close to the MO leaf protein isoelectric point and have maximized the precipitated protein content with the highest yield. The temperature did not have a significant effect on this process. After both stages (extraction and precipitation), it was possible to obtain up to 26.93±0.23 g 100 g-1 of the protein content of the original leaf, which meant 7.26±0.19 g 100 g-1 of the dry leaves. This yield was similar to that of the alfalfa concentrates that are generally available on the market. Optimizing the process of extraction and precipitation of protein from MO leaf offers a new option for plant protein that could become a strategy to combat malnutrition in developing countries.

