










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Ciencias de la Salud
Print version ISSN 1692-7273On-line version ISSN 2145-4507
Rev. Cienc. Salud vol.7 no.1 Bogotá Jan./Apr. 2009
Cerebral Anatomy of the Spider Monkey Ateles Geoffroyi Studied Using Magnetic Resonance Imaging. First Report: a Comparative Study with the Human Brain Homo Sapiens
Anatomía Cerebral del mono araña Ateles geoffroyi estudiada utilizando imágenes de resonancia magnética. Primer reporte: estudio comparativo con el cerebro humano Homo Sapiens
Fernando Chico-Ponce de León,
1. Laboratorio de Neuromorfología, Instituto Nacional de Psiquiatría Ramón de la Fuente, México, D. F.
2. Posgrado en Antropología, UNAM, Instituto de Investigaciones Antropológicas, México, D. F.
3. Grupo de Cronoecología y Etología Humana, Instituto Nacional de Psiquiatría Ramón de la Fuente Muñiz. México, D. F.
4. Resonancia Magnética, Hospital Ángeles del Pedregal, México D.F.
5. Laboratorio de Neuropsicología, Facultad de Medicina, UNAM, México, D. F.
6. Hospital Infantil de México Federico Gómez, México, D. F.
7. Posgrado, Facultad de Medicina, UNAM, México, D. F.
8. Hôpital de Neurologie, Nancy, Cedex, France.
9. Facultad de Psicología, UNAM, México, D. F.
10. Servicios Clínicos, Instituto Nacional de Psiquiatría Ramón de la Fuente, México, D. F.
Correspondence to: Fernando Chico Ponce de León. Laboratorio de Neuromorfología, Instituto Nacional de Psiquiatría Ramón de la Fuente Muñiz. Camino a Xochimilco 101, San Lorenzo-Huipulco, Delegación Tlalpan, C.P. 14370 e-mail: chico1204@prodigy.net.mx
Recibido: febrero 26 de 2009 Aceptado: marzo 16 de 2009
Abstract
The objective of the present qualitative study was to analyze the morphological aspects of the inner cerebral anatomy of two species of primates, using magnetic resonance images (MRI): spider monkey (A. geoffroyi) and human (H. sapiens), on the basis of a comparative study of the cerebral structures of the two species, focusing upon the brain of the spider monkey and, primarily, its limbic system. In spite of being an endemic Western hemisphere species, a fact which is by its own right interesting for research due to this animals social organization and motor functions, the spider monkey (A. geoffroyi) has hardly been studied in regard to its neuroanatomy. MRI was carried out, in one spider monkey, employing a General Electric Signa 1.5 T scanner. This investigation was carried in accordance to international regulations for the protection of animals in captivity, taking into account all protective means utilized in experimental handling, and not leaving behind any residual effects, either physiological or behavioral. From a qualitative point of view, the brains of the spider monkey and the human were found to have similar structures. In reference to shape, the most similar structures were found in the limbic system; proportionally, however, cervi cal curvature, amygdala, hippocampus, anterior commissure and the colliculi, were larger in the spider monkey than in the human.
Key words: Amygdala, Hippocampus, Limbic System.
Resumen
El objetivo del presente estudio cualitativo fue analizar los aspectos morfológicos de la anatomía cerebral interna utilizando imágenes de resonancia magnética (IRM) en dos especies de primates, El mono Araña (A. geoffroyi) y el humano (H. sapiens), tomando como base un estudio comparativo de las estructuras cerebrales de las dos especies, concentrándose primordialmente en el sistema límbico del cerebro del mono araña. Aunque es una especie común en el hemisferio occidental, es interesante para estudiar dada su organización social y funciones motoras, el mono araña (A. geoffroyi) ha sido poco estudiado en cuanto a su neuroanatomía. Las IRM fueron hechas a un mono araña utilizando un resonador General Electrics Signa 1.5 T. Esta investigación se llevo a cabo conforme a las leyes internacionales para la protección de animales en cautiverio y teniendo en cuenta todas las medidas de protección para el manejo experimental para evitar cualquier efecto residual de índole comportamental o fisiológico.
Desde un punto de vista cualitativo, los cerebros del mono araña y el humano tenían estructuras similares. Con respecto a la forma, las estructuras más parecidas fueron encontradas en el sistema límbico, sin embargo la curvatura cervical, la amígdala, el hipocampo, la comisura anterior y el colículo fueron más grandes proporcionalmente en el mono araña que en el humano.
Palabras clave: Amígdala del Cerebelo, Hipocampo, Sistema límbico
INTRODUCTION
In Western tradition, the first documented information concerning the similarity between humans and the rest of primates goes back to [1]. Other the naturalists of Antiquity were, Caius Plinius Secundus and. The Dr. Hernández [2] mentioned the existence of New World Monkeys Urbani, [3]. In the 18 th century the works of Buffon and in the 19 th century, Charles Darwin continued the study of primates in his work [4]. In this time, [5], zoologist and paleontologist, On the Brain of Ateles paniscus and On the Relations of Man to the Lower Animals, shows the neuroanatomical interest given to non-human primates brain, as he is the first person to perform an anatomical study of the spider monkeys brain. We have been unable to find references about anatomical structures of this primate afterwards. Some experts in comparative anatomy, [5, 6] published about the diverse structures in the brain of non-human primates, such as the occipital body, the occipital lobe and the minor hippocampus (now calcar avis). Of special importance in this sense are Paul Brocas works of comparative anatomy, on what he called le grand lobe limbique of mammals in 1878. [7]. Wensceslas Papez proposal of 1937 took into account Brocas morphological suggestion in reference to the biglimbic lobe. Thus, Papez proposed that a part of the circuit of emotion lay inside this brain zone[8]. MacLean suggested that all structures relating to the survival instinct are part of thelimbic system [9]. Connolly, in the middle of the 20 th century, further reinforced the similarity of the brains of primates and helped to identify the differences in the best way possible. In his investigations, this author stressed that the great apes present complex sulci patterns exhibiting both intra- and interspecific variations [10]. Today, cerebral atlases of primates exist: rhesus macaque (M. mulatta) [11]; Paxinos et al, [12] and the vervet monkey (Cercopithecus aethiops) [13], the squirrel monkey (Saimiri sciureus) [14].
Detailed anatomical studies on the brain of the spider monkey, an endemic Western hemisphere primate, have not been carried out until now. The genus Ateles shows a high degree of morphological and neural adaptations [15]. This species presents a bigger relative mass of caudal muscles [16] and larger areas of muscle units in the vertebrae [17].
METHODS
Procedure and veterinary handling
The one spider monkey used here was not given food or water for 8 hours before initiating the study. After 8 hours, anesthesia was applied to the caged animal employing 0.70 ml of Zoletil 50 (Tiletamine), so that the monkey could be taken for a functional MRI study. The second dose of anesthetic was applied before inserting the head of the monkey into the resonance magnet. This dose was of 0.35 ml of the same drug. Afterwards, an acrylic fastener was placed around the animals head, and MRI of the brain began. After 30 minutes, the monkey was withdrawn from the scanner and reintroduced into the cage. After a recovery period of 30 minutes the monkey was given water and fruits.
This study complies with the Mexican laws for animal management and care. An adendum for this work was also aproved by the ethical comite of the National Institute of Psychiatry.
Magnetic resonance imaging
A General Electric Signa 1.5 T scanner, at 33 mT and a 34 slew rate, using a knee coil with 2 quadrature-channels for higher definition with a 20 cm field of view, and an acquisition matrix of 512 x 224 was used for MRI studies. Sixty axial slices were taken going from the medullary bulb to the highest cerebral convexity, using sequences of 3D-FSPGR volume, with a thickness of 1 mm and 0 mm of spacing, and a repetition time (TR) of 18.8 and an echo time (TE) of 4.2.
Tridimensional reconstructions were undertaken utilizing an Advantage Windows 4.0 workstation. Afterwards, a representative selection of slices in the three planes was obtained in order to carry out a qualitative and quantitative analysis (measurements with CD Viewer, GE Medical Systems) of the images.
Analysis
A nine-year-old female adult spider monkey (A. geoffroyi) weighing 8.79 kg and in good health condition was chosen for the study. Employing a bicommissural reference, a selection of slices in three spatial dimensions, axial, coronal and sagittal, was obtained with the objectives to localize structures of the internal cerebral anatomy of the spider monkey, especially of the fifth temporal gyrus, the anterior white commissure, the gyrus of the cingulum and the great sulci, and to compare these structures afterwards to the cerebral structures of the human (H. sapiens). The measures of the structures were carried out by DICOM Viewer Program. The structures of the brain, especially the gyruses, in all extensions and directions, were precisely located in the images of the spider monkey brain using a biplanar reconstruction on the basis of a spoiled gradient recalled (SPGR) sequence. Magnetic resonance (MR) images obtained were compared to those of two female adult humans (19 and 31 years old), free of any type of pathology, and subjected to T1 and T2 MR imaging, using as reference the spatial planes of the slices of the spider monkey. Afterwards, a series of drawings based on Duvernoy [18] was made in order to analyze the MRI results more precisely. A qualitative analysis of cerebral structures of the spider monkey was carried out.
RESULTS
In this study, the structures mentioned were compared, such as the fifth temporal gyrus, the medial face of the hemisphere, the anterior white commissure, the fornix, the hypothalamus and the third ventricle, among others.
Sagittal section of the brain of the spider monkey with measurements in millimeters (Fig. 1 A, C, E, Fig. 2 D, Table 1).
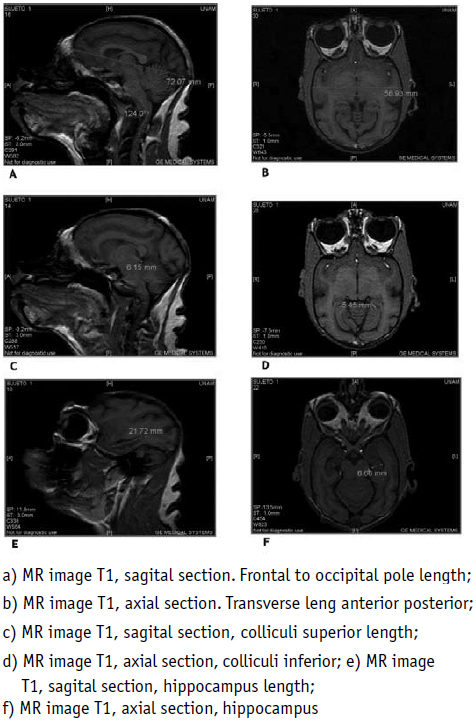
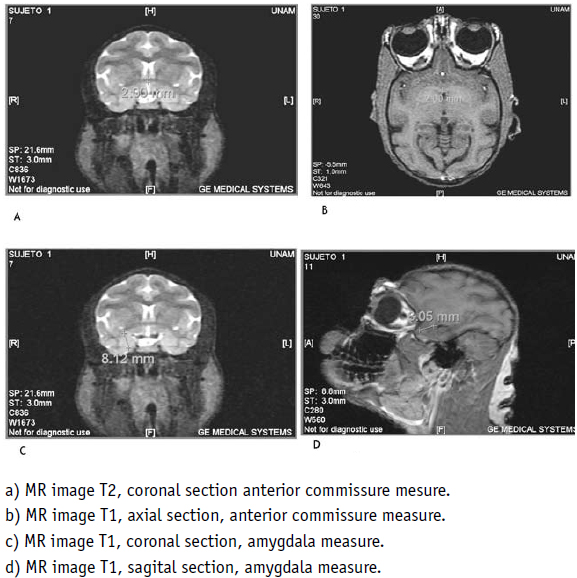
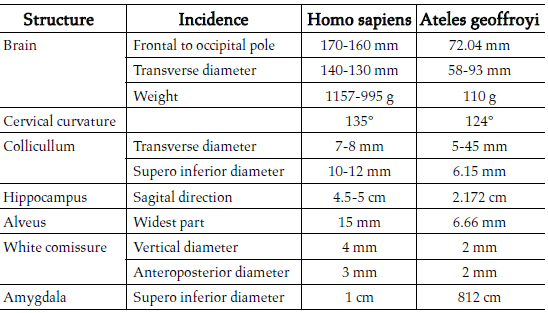
Table 1. Comparative measures between Human and Spider monkey brains
(All MR images were taken in T1):
The medial faces seen are those of the right hemisphere. In Figures 3, 4, 5, 6 and 7 similarities among the two species of primates were found, especially in regard to the morphology of the cingulate gyrus, the fornix, and the anterior white commissure, with the human one presenting greater complexity in its pattern of gyration.
Concerning differences, strong interthalamic adhesions in the brain of the spider monkey were found, a cerebral structure which usually is very slim in the human. A small frontal lobe, as well as a more pronounced cervical curvature, was also determined. The cervical curvature in the human has a 135° angle, according to Testut and Latarjet, while the angle of the same curvature in the spider monkey was 124° (Fig. 1 A). The largest axis of the colliculi in the human measures 10-12 mm, from the front to the outside, and the transverse axis just 7-8 mm, whereas in the spider monkey the superoinferior diameter of the superior colliculi was 6.15 mm and their transverse diameter 5.45 mm (Fig. 1 C, D).
The anteroposterior diameter from the frontal pole to the occipital pole of a human brain measures 17 cm in a male and 16 cm in a female person. The transverse diameter in a male human brain measures 14 cm and 13 cm in a female brain. In the spider monkey, the anteroposterior diameter measured 72.07 mm and the transverse diameter, perpendicular at the bicommissural line, 58.93 mm (Fig. 1 A, B).
The retrocommissural hippocampus, as regards Ammons horn, the dentate gyrus and the subiculum, measures, in the human, in the sagittal direction, between 4.5 and 5 cm, whereas, in the spider monkey, according to our measurements, 21.72 mm (Fig. 1 E). The widest part of the alveus measures 15 mm in the human [19] and in the spider monkey 6.66 mm (Fig. 1 F).
One may see there the corresponding hypointensity of Ammons horn and the dentate gyrus, in lengthened form, from the back side to the front and from the top to the bottom, with the extreme anterior end folded upon itself (Fig. 5, No 2), and the amygdala located in front of it (Fig. 5, No 1), always with an irregular spherical form. Between this structure and the ventral striatum, lies the anterior commissure. In the human brain, the vertical diameter of this cylindrical structure is 4 mm and the anteroposterior diameter 3 mm [20]. The MRI analysis disclosed that both diameters of this structure in the brain of the spider monkey were 2 mm (Fig. 2 A, B), and one could also observe the internal capsule and the caudate nucleus above the temporal recess of the lateral ventricle (Fig. 5 No. 6, 7).
The human brain presented a similar structure, with the retrocommissural hippocampus, where Ammons horn and the dentate gyrus are folded upon themselves, in its anterior part (Fig. 6, 7, No. 2). Above and in front of it, the amygdala was identified, than in the monkey brain (Fig. 5, No. 2). In the human brain, the amygdala measures 1 cm in diameter [19]. The one measured in the spider monkey of the present study had a superoinferior diameter measuring 8.12 mm, in a coronal section, and 8.06 mm, in a sagittal section (Fig. 2 C, D). With measurements, Similarities the brain of another primate was found, the rhesus monkey and may be confirmed perusing Saleem and Logothetiss [11] and Paxinos et al [12] atlases.
In both primates, the human and the spider monkey, the temporal recess of the lateral ventricle (Fig. 5, 6, No. 11) lay next to the alveus and the fimbria of the hippocampus, the posterior part of Ammon´s horn and the end of the dentate gyrus at the ventricular atrium (same Fig. (sF) 5, 6, No. 12) (see in particular Fig. 7, No. 12 of Duvernoy). The same disposition could be seen in the case of the anterior white commissure, the striatum, the internal capsule and the tail of the caudate nucleus (Fig. 5, 6 No. 4, 5, 6, 7).
Figure 3. Spider monkey
Figure 4. Human, female, 31 years of age MR image T1, without contrast, midsagittal section, indicating the structures mentioned in figure 3


Figure 6. Human, female, 19 years of age MR image T1, without contrast, sagittal longitudinal section of the right retrocommissural hippocampus
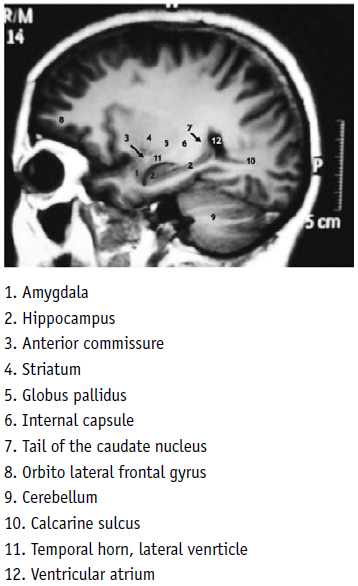
Figure 7. Human Drawing, based on Duvernoy 1998:167,97F, illustrating the similarity to the hippocampus area of the psider monkey (fig 5) (same structures as shown in fig 6)

Axial section
In the axial section, similar differences were observed, when comparing the spider monkey and the human on a strict bicommissural line.
In the images of the spider monkey, the ventral part of the striatum could be clearly identified (Fig. 8 A [T2], B [T1] and C, No. 6), as could also be the anterior white commissure (same Fig. No. 5), the inferior part of the internal capsule (sF No. 7), the interthalamic adhesion (sF No. 11), the fornix (sF No. 1), the posterior columns of the fornix (sF No. 12), the disposition of the opercula covering the insula (sF No. 18), the olfactory structures (sF No. 4) and the occipital recesses of the lateral ventricles (sF No. 17). This section type permitted observing the superior part of the mesencephalon, setting the profile of the cerebral peduncles and colliculi in evidence.
The calcarine sulcus was easy to identify in the spider monkey (sF No. 16). On the other hand, employing the same section incidence, one may only with difficulty visualize this sulcus in man.
Figure 8. Spider monkey
In the bicommissural plane, in the human (fig. 9 [T1]), one could notice straight away the big frontal lobe as the main difference, which in the spider monkey was small, and almost exclusively limited by the straight gyrus and the olfactory sulcus. We found in the spider monkey that there was a simpler pattern of gyration (Fig 8 No. 4, 10, 13, 18). The central cerebral structures were similar in the ventral part of the striatum (sF No. 6), the thalamus, the internal capsule (sF No. 7), the anterior and posterior columns of the fornix (sF No. 1 and 12), the pineal body and the cerebellar culmen (sF No. 3), as well. The anterior and posterior white commissures (sF No. 5, 8), the ventricular atrium (sF No. 17) and the large sulci were also similar.
The gyration of the insula was simple in the non-human primates, in comparison to that of the human. In the three primates, Homo sapins, Macaca rhesus and Ateles geoffroyi the insula was covered by the cerebral opercula, although in the non-human primates these were thinner [Saleem & Loghothetis 11; Paxinos et al 12].
Figure 9. Human, female, 19 years of age MR image T1, axial section, bicommissural plane

Figure 10. Spider monkey
The axial sections at the level of the nucleus of amygdala and the anterior part of the hippocampus
The structures of the temporal mesiobasal region were identified without problem in the MRI of the spider monkey (Fig. 10). The section passed at the level of the optic chiasm (sF No. 8), al level of the entrance of the optic nerves to the cranium and, at a more posterior, the optic chiasm (sF No. 1). One could also identify the upper part of the cerebral peduncle (sF No. 2) and its relation to the optic tract. From the outside, both the amygdala and the hippocampus (sF No. 6, 7) could be easily identified, as well as the temporal recess of the lateral ventricle (sF No. 3). The temporal pole advanced a little in front of these structures. The temporal lobe extended to a section of the occipital lobe (sF No. 13). In the posterior part of the mesencephalon, the inferior part of the mesencephalic ventricle (sF No. 9), the superior colliculi and, immediately on the back side, the cerebellar culmen (sF No. 14).
In the human (Fig. 11, 12), in a section that passed over the chiasm (sF No. 8) and the hypophyseal infundibulum (sF No. 11), at the level of the cerebral peduncles (sF No. 5), one could also observe the mesiobasal structures, the amygdala (sF No. 6) and, on the back side, in the fifth temporal gyrus, the retrocommissural hippocampus folded upon itself, with the structures of Ammons horn and the dentate gyrus (sF No. 7), as well as the temporal recess of the lateral ventricle in its anterior part (sF No. 3). In a similar manner, one could define the temporal pole (sF No. 10) and the cerebellar culmen (sF No. 14). The upper part of the fourth ventricle was evident towards the end of the mesencephalic aqueduct (sF No. 9).
Figure 11. Human, female, 19 years old MR image T1, showing the same structures in the areas of the amygdala and the hippocampus and the similarity to the spider monkey (fig. 10).

Figure 12. Human, female Drawing, based on Duvernoy 1998:197,105E, illustrating the similarity in the area of the amygdale and the hippocampus to the spider monkey (fig. 10).

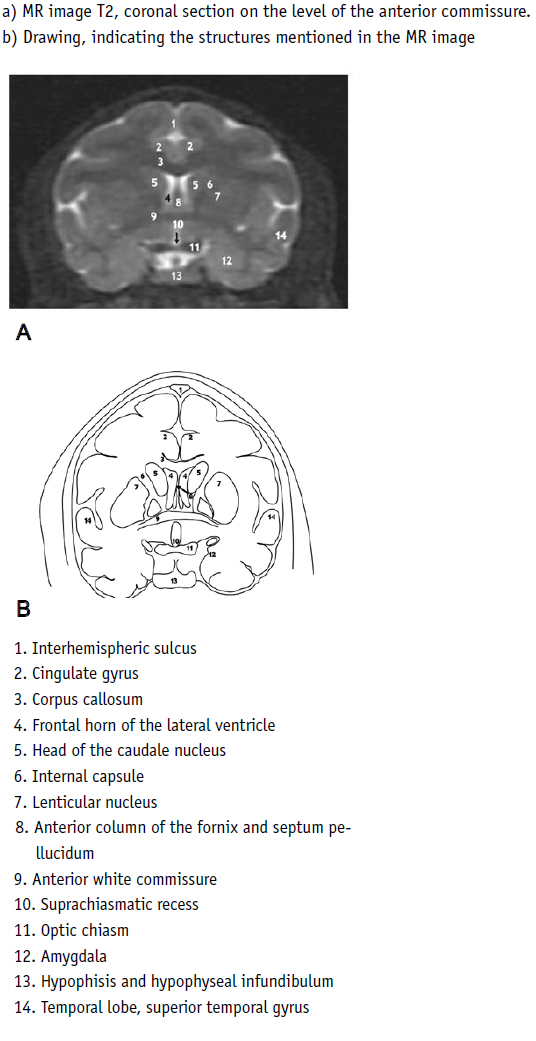
Coronal section:
Two of these sections of the spider monkey (Fig. 13, 14 [T2]) were analyzed. The anterior section lay at the level of the anterior white commissure (Fig. 13 No. 9), which was found to reside in the posterior part of the anterior columns of the fornix (sF No. 8), at the level of the frontal recesses of the lateral ventricles (sF No. 4), and the anterior part of the head of the caudate nucleus (sF No. 5). The corpus callosum was found above these structures (sF No. 3), and one could see the anterior arm of the internal capsule (sF No. 6) nested between the caudate nucleus and the lenticular nucleus (sF No. 7). Above the corpus callosum, the cingulate gyruses could be identified (sF No. 2). Below the anterior white commissure, one found the suprachiasmatic recess (sF No. 10), below it, the optic chiasm (sF No. 11) and, at one side and from below, the amygdala (sF No. 12) could be determined. The insula was covered by the opercular system.
In the posterior coronal cut (Fig 14), the splenium of the corpus callosum (sF No. 3), the gyruses and the sulcus of the cingulum (sF No. 2) could be clearly identified. Below and from the outside of the splenium, one could observe the ventricular atrium (sF No. 4) and, inside it, the posterior columns of the fornix, parallel to the posterior part of the alveus. The parietal areas could be seen in the marginal part of the lateral sulcus (sF No. 12). Below the splenium, one could determine the deep venous confluence (sF No. 8) and, below this, the cuadrigeminal cystern spaces and the pons (sF No. 6), as well as a part of the fourth ventricle (sF No. 14), and the cerebellum with the middle cerebellar peduncles (No. 10), a voluminous flocculonodular lobe (sF No. 7) and the superior part of the bulb (sF No. 11).
In the human (Fig. 15), in the coronal section, at the level of the anterior white commissure, one could see all the structures as in the spider monkey but varying with respect to a more complex anatomical pattern.
DISCUSSION
Limits of the structures mentioned in this work; amygdala, hypocampus and the anterior white commisure, where based on the atlas of [11, 12] in rhesus monkey primate species.
This authors work confirms that humans possess a highly developed frontal lobe, in contrast to the spider monkey, whose frontal lobe is generally of a simpler and smaller configuration.
Evidence is provided for the fact that the limbic structures the fifth temporal gyrus, the amygdala, the hippocampus, the anterior white commissure, the ventral striatium and the perihippocampal area, are similar in form in the two primates, the human and the spider monkey. The limbic lobe was similar in the two primates.
The constant sulci, such as the lateral, the central, the parieto-occipital, the calcarine and the cingulated, were also similar in the two primates. The simple pattern of gyration is similar to the one observable in the human fetus. One could also notice that the anterior white commissure was thicker in the non-human primates.

It is well known that the similarities between the primates are not only anatomic but cytoarchitectonic and neuroendocrinological as well as from the neurobiochemical points of view [11, 21, 22]. To this day, the most studied non-human primate has been the rhesus macaque. A detailed anatomic study of the brain structures of the spider monkey enriches general knowledge in this field of research.
It is also well known that, today, an approximation to cerebral anatomy and physiology has become less complicated, both in the human and in the primates, thanks to MRI. This work confirms these observations in regard to the spider monkey. The clearness of the images and the possibility of transpolation of information from the rhesus macaque as well as from the human to the cerebral anatomy of the spider monkey will make complex atlas elaboration easier in the near future, and increase its usefulness in biological investigation, in general. This approximation may also be used to suggest concepts explaining the species-specific differences of behavior in relation to the development of diverse structures and add support to the argument that the development of those structures favors the diverse adaptations observed. As the amygdala, hypocampus and the anterior white commisure are basically related to emotional behavior, and considering that the spider monkey has a fusion-fission social organization system, these structures have a fundamental participation in the daily life of the species, as during the noon fusion animals must invest in activity to establish fraternal relations with other members of the group in order to achieve comfort and thermoregulation through the night, a characteristic of most of diurnal primates [23, 24]. We suggest, as does Sherwood and cols. [25], it is necessary to perfom more extensive on the relation between structure and function, as well as to enlarge the spider monkey study sample. Some authors stress, though, that even if the brains are similar when macroscopically compared, they may vary considerably in cytoarchitecture, electrophysiology and chemical functioning [Holloway, 26].
Nonetheless, a macroscopical comparison does provide valuable information concerning the neurological stage of development, the similarities and morphological differences in the brains of the two primates and the probable morphological adaptations of each species.
Today, in this particular field of neuroscience, there is a tendency towards providing more tools for the understanding of brain structures and their function. Therefore, besides the objectives included in this work, there are increasing attempts to work in scientific, medical and cerebral areas relating to anthropological, neurophysiologic and pharmacological perspectives of viewing the spider monkey. This same approximation may also suggest elements for a correlation between the development of diverse cerebral structures and its relation to the behavior of the primates. Lines of investigation based on the high degree of telencephalization of the non-human primates are proposed, which allow us to infer a very complex social organization and cognitive and behavioral activities in these animals. Such lines of research allow carrying out comparative species-specific studies of the primates. The cortical functional areas in the spider monkey, such as the area responsible for motility and sensitivity of the gripping tail, still have to be identified.
Figure 15. Drawing based on Duvernoy, 1998, coronal section, illustrating the similarity on the level of the anterior white commissure to the spider monkey (fig. 14).

ACKNOWLEDGMENTS
This project were approved by ethical comitte of Instituto Nacional de Psiquiatría Ramón de la Fuente Muñiz. No animal were injured as a result of this research. The author aknowledments to Virbac Labotatories, for Zoletil 50 anesthesia donation.
REFERENCES
1. Aristóteles. Historia de los animales. Editorial Gredos, Madrid. 1978. [ Links ]
2. Hernández F. Obras completas. Historia natural de Cayo Plinio Segundo, tomo IV, volumen I. Universidad Nacional Autónoma de México: México. 1996. [ Links ]
3. Urbani B. Nuevo Mundo, nuevos monos: sobre primates neotropicales en los siglos XV y XVI Bol Asoc Primatol Esp. 2001 8/1. [ Links ]
4. Platas D. El uso de los modelos de primates para explicar la evolución de la organización social en la evolución humana. Una crítica epistemológica. Tesis de maestría. Instituto de Investigaciones Antropológicas- Facultad de Filosofía y Letras, Universidad Nacional Autónoma de México, Ciudad de México. 2006. [ Links ]
5. Huxley TH. On the brain of Ateles paniscus. Zool Soc London. Scient Mem II, London, 1861, pp 493:508. [ Links ]
6. Gratiolet LP. Mémoire sur les plis cérébraux de lhomme et des primates. Bertrand: Paris. 1854. [ Links ]
7. Broca P. Anatomie comparée des circonvolutions cérébrales. Le grand lobe limbique et la scissure limbique dans la série des mammifères. Rev Anthropol. 1878.1, 2:385-498. [ Links ]
8. Papez W. A proposed mechanism of emotion. Arch Neurol Psychiat. 1937 38:725-743. [ Links ]
9. MacLean PD. Contrasting functions of limbic and neocortical systems of the brain and their relevance to psychophysiological aspects of medicine. Am J Med. 1958, 25/4:611-26. [ Links ]
10. Conolly CJ. External morphology of the primate brain. Charles C Thomas: Springfield. 1950. [ Links ]
11. Saleem KS, Logothetis NK. A combined MRI and histology atlas of the rhesus monkey brain, Elsevier: San Diego California. 2007. [ Links ]
12. Paxinos G, Xu-Feng H, and Arthur WT. The rhesus monkey brain in stereotaxic coordinates. Academic Press: San Diego California. 2000. [ Links ]
13. Contreras C, Mexicano G, Guzmán-Flores C. Astereotaxic brain atlas of the green monkey (Cercopithecus aethiops). Universidad Nacional Autónoma de México. Bol Estud Méd Biol 1981.31/7-8:383-428. [ Links ]
14. Gergen JA, MacLean PD. Links in the limbic system: photic activation of limbic cortical areas in the squirrel monkey. Ann N Y Acad Sci. 1964, 10/117:69-87. [ Links ]
15. Rosenberg AL. Tale of tails: parallelism and prehensility. Am J Phys Anthropol. 1983, 60:103-107. [ Links ]
16. Lemelin P. Comparative and functional myology of the prehensile tail in new world monkeys (Alouatta, Ateles & Saimiri). J Morphol. 1995, 224:351-368. [ Links ]
17. German R. Functional morphology of new world monkeys tails. Am J Phys Anthropol. 1981, 54:224. [ Links ]
18. Duvernoy H. The human brain. Springer-Verlag: Wien, New York: 354 pp. 1991. [ Links ]
19. Bricout J. Anatomie descriptive du système nerveux central, Fascicule II, Librairie Maloine: Paris. 1965. [ Links ]
20. Testut L, Latarjet A. Tratado de anatomía humana, tomo II, 2ª ed, Salvat Editores: Barcelona. 1976. [ Links ]
21. Semendeferi K, Damasio H, Frank R, Van Hoesen GW. The evolution of the frontal lobes: A volumetric analysis based on three-dimensional reconstructions of magnetic resonance scans of human and ape brains. J Human Evol. 1997, 32/4:375-388. [ Links ]
22. Chiarelli B. Pheromonal communication and socialization. In: Evolutionary anatomy of the primate cerebral cortex, D Falk & K Gibson (eds), Cambridge University Press: Cambridge, pp. 165-176. 2001. [ Links ]
23. Anderson JR. Sleep-related behavioural adaptations in free-ranging anthropoid primates. Sleep Medicine Reviews. 2000, 4(4):355-373. [ Links ]
24. Muñoz-Delgado J, Corsi-Cabrera M. Chronoecology of neotropical primates: the spider monkey Ateles geoffroyi. In Frank Columbus (ed.). Trends in Chronobiology (en prensa). [ Links ]
25. Sherwood Ch, Cranfield M, Mehlman P, Lilly A, Garbe AL, Whittier CH, Nutter F, Rein Th, Bruner H, Holloway R, Tang Ch, Naidich Th, Delman B, Steklis D, Erwin J, Hof P. Brain Structure Variation in Great Apes, With Attention to the Mountain Gorilla (Gorilla Beringei Beringei). American Journal of Primatology. 2004, 63:149-164. [ Links ]
26. Holloway RL. Evolution of the human brain. In: Handbook of human symbolic evolution. A Lock & C Peters (eds.), Oxford University Press: New York, pp 41-49. 1996. [ Links ]

