











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
Colombia is among the countries with a high number of cultivated Passiflora species because of its variety of ecological habitats (Ocampo et al., 2016). A total of nine species are cultivated from sea level to 2,800 m, and the more important ones are P. edulis f. flavicarpa Degener (yellow passion fruit), P. ligularis Juss. (sweet granadilla), P. edulis f. edulis Sims (purple passion fruit), P. maliformis L. (stone granadilla) and P. tarminiana Coppens (Banana passion fruit). The cultivated area in Colombia for these species covers about 10,000 ha (Agronet, 2019). Water is essential for passion fruit plants because it promotes good growth and increases productivity, allowing continuous and regular production with good fruit quality (Jehová et al., 2013).
The productivity and development of plants from the genus Passiflora are affected by solar radiation, temperature, sunshine and soil moisture loss. Factors such as water stress and extreme salinity conditions (> 2.5 dS m-1) limit the productive potential of passion fruit plants (Cavalcante, 2012). A frequent and adequate water supply allows continuous flowering and fruiting so long as other environmental conditions are favorable. Therefore, to achieve good yield and good fruit quality, water must be supplied in adequate amounts.
According to Rodríguez et al. (2019), the term water deficit defines an essentially ecophysiological concept related to the limitation of water in tissue that is close to being synonymous with drought stress because it refers to any limitation in the ideal operation of a plant imposed a limited availability of water.
There are two types of water stress: one that is due to a total lack of rain and another caused by poor distribution of rain throughout a plant’s growth period (Rodríguez et al., 2019); in both cases, agricultural production is seriously affected. Anthropogenic global climate changes and phenomena such as El Niño (Phenomenon of the Pacific) are typical examples that induce abnormal rainfall distribution in agricultural regions. When this phenomenon occurs in the Colombian regions of the Caribbean and the Andes, with the exception of Magdalena Medio, there are reductions in precipitation values for accumulated volume while the phenomenon lasts, near or above 20%. Regions such as Guajira, northern Cesar, the municipalities Atlantico, Bolivar, Sucre, Cordoba, the coffee triangle, Santander and Cundinamarca, in Colombia, show a highly marked decrease in precipitation, with values equal or higher to a 60% reduction in normal rainfall (Fan et al., 2017). Some of the affected regions due to drought are induced by the El Niño phenomenon, which hinders the production of the areas cultivated in passionfruits due to non-tolerance to the conditions of hydric stress. Likewise, fluctuations in the distribution of rainfall caused by global warming can increase risk from being repeatedly exposed to plant droughts (FAO, 2020).
Saline soils are defined as those adversely modified for the growth of most of these species as the result of the presence of soluble salts, exchangeable sodium or both in the root zone (Yertayeva et al., 2019). Soils affected by these salts are common in arid and semiarid regions because of low rainfall and high evaporation (Fageria et al., 2010).
Saline stress, is related to two types of effects: osmotic and ionic. The first is caused by excess salts that are biophysical in nature, with osmotic effects standing out, restricting water transport (Silveira et al., 2010). Then, a sequence of reactions modulated by hormones is quickly triggered, leading to a restriction of stomatal opening and photosynthetic assimilation of CO2. These effects predominate in the first phase of salt stress "osmotic phase", which occurs in the initial stages of plant exposure to salinity or in the presence of moderate levels of salts in contact with the root system. The same authors reported that the “osmotic phase” has a physiological response: acclimatization to stress in response to damage suffered caused by the salt stress. This is often diagnosed as symptoms of negative effects of salt stress but are actually normal plant physiological responses to overcome or acclimate to an adverse situation.
As excess saline ions accumulate in the cytosol of the cells, toxicity problems will arise (toxic or ionic phase) in plants exposed to salinity (Silveira et al., 2010). However, species differ widely in protoplasmic resistance or tissue resistance to salt stress. This ability to resist is linked mainly to the intensity of compartmentalization of saline ions in vacuoles and to the maintenance of a favorable K+/Na2+ balance in the cytosol. Currently, this has been a target for the selection of resistant cultivars from some cultures. This study aimed to determine tolerance to water and salt stress in four cultivated Passiflora L. species.
MATERIALS AND METHODS
The present study was carried out between February and September, 2016, under hydroponic conditions in a greenhouse. The city of Manizales is in the coffee triangle, in the center of the Department of Caldas, with the coordinates 5°03’23.31” N and 75°29’41.56” W, at an altitude of 2,130 m a.s.l., with an average external temperature of 18°C, average annual rainfall of 2,000 mm and average relative humidity of 78%.
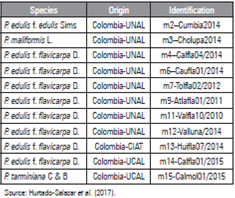
Eleven elite accessions of four Passiflora species of commercial interest were evaluated, which have been previously assessed, characterized and selected in several producing regions of Colombia based on the improvement program of Ocampo et al. (2013) and different gene banks from several institutions (Tab. 1).
Table 1 Commercial Passiflora accessions from different gene banks in Colombia selected to evaluate abiotic stresses (salinity and drought).
The experiment design was completely randomized in an array of sub-split plots, where the plot was the percentage of water saturation in relation to the requirements of the crop (100 and 33%). The subplot consisted of saturation levels of salt (sodium chloride reactive level - 99.9% purity) (EC: 1.5 and 5.5 dS m-1), and each sub-subplot contained the 11 Passiflora accessions in a completely randomized array with five replicates, where the experiment unit consisted of one plant.
240 seeds of each accession were planted, germinated in a hydroponic bed using rice husk as substrate, which was constantly kept wet until transplantation. After 15 d of germination, 54 seedlings of each accession were selected to ensure greater uniformity in the root system.
Subsequently, the plants were transferred to hydroponic beds that were 6 m long and 1.2 m wide to ensure the irrigation regime of the treatments (percentage of water saturation 100 and 33%, and salt saturation levels EC: 1.5 and 5.5 dS m-1), according to the experiment design. A conduit system was used in a vertical trellis, with 12 gauge galvanized wire, 2 m above the hydroponic greenhouse bed soil. The plants were conducted on a single stem, with weekly pruning of the side branches. When the main stems surpassed the wire at 20 cm, they were bent down, forming a loop. The secondary and tertiary branches were maintained.
The Hoagland and Arnon’s universal nutrient solution (1950), as modified by Niu et al. (2015), was used. To ensure the salinity of the treatments, each solution was brought to a value of EC: 1.5 and 5.5 dS m-1, with the addition of reagent level sodium chloride (99.9% purity). To evaluate the interaction between the percentage of water saturation and salinity, electrical conductivity was added to the different treatments (1.5 and 5.5 dS m-1) to the percentage of water of 100 and 33% for each experiment units of the evaluated accessions.
The production efficiency index (PEI) for dry matter was estimated to assess the effect of the water and saline stress. This index classified the tolerant genotypes. This ratio can be calculated as described below (Fageria et al., 2010):
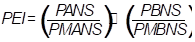
where, PEI - efficiency index of dry matter production , PANS - production of dry matter with high salinity; PMANS - average production of the experiment with high salinity; PBNS - production with low salinity; PMBNS - average production of the experiment with low salinity.
Analysis of variance was performed, and the averages were compared using the Tukey test, with a confidence level of 95 using SAS statistical package (Statistical Analysis System, 2013).
RESULTS AND DISCUSSION
According to Fageria et al. (2010) and their proposed efficiency index of dry weight production in the aerial part. When the EIP is greater than 1.0, cultivars are classified as tolerant; the range of 0.5 to 1.0 indicates moderate salinity tolerance; a value less than 0.5 means the genotypes are classified as sensitive to salinity; therefore, it is worth noting that three accessions were tolerant, and three accessions were moderately tolerant (Tab. 2).
The total dry weight per accession decreased by approximately 21.2% under the water (33% humidity) and saline (5.5 dS m-1) stress conditions, except in the tolerant accessions m15, m13 and m11, which showed a dry matter accumulation higher than other accessions evaluated under similar conditions of maximum stress (33% moisture and 5.5 dS m-1) and optimum humidity and electrical conductivity (100% and 5.5 dS m-1). The accessions m2 (P. edulis f. edulis), m12 (P. edulis f. flavicarpa), and m14 (P. edulis f. flavicarpa) obtained PEI values between 0.5 and 1.0 and were classified as moderately tolerant.
Table 2 Influence of salinity on the dry matter of the aerial part in 11 Passiflora accessions and its classification in terms of tolerance to salinity according to the Production Efficiency Index (PEI).

Cellular osmoregulation gives plants the ability to tolerate conditions with a lack of water and high salinity, with the expression of adaptive mechanisms that prevent the reduction of photosynthesis, changes in translocation and distribution of photoassimilates, and decreased yield. These events are of great importance in the normal growth and development of plants and, consequently, in crop productivity.
On the other hand, the sensitive accessions showed a 45% decrease in the accumulation of dry weight. A study on P. edulis by Gomes et al. (2012) showed a reduction in the dry matter content in the leaves, stem and roots when plants were exposed to water stress. Rodríguez et al. (2019) concluded that exposure of P. ligularis to droughts resulted in a significant decrease in leaf area, number of leaves per plant and length of the branches. However, the root-aerial ratio, based on the dry weight and specific weight of the leaves, increased as the water stress increased in the plants. The distribution pattern of dry leaves, branches, stems and roots was altered by the drought.
The negative effect of the water deficit was reflected in a growth reduction in the aerial part since a lot of assimilates were invested in the growth of the root system. Modifications in the root system occur because the root volume increases under water stress conditions, which favors water uptake and decreases the aerial part in order to reduce the rate of leaf transpiration (Rodríguez et al., 2019). Nevertheless, Hurtado-Salazar et al. (2018 and 2017) found that the accession m15 (P. tarminiana) may be able to survive with salinity up to 5.5 dS m-1 (NaCl), which is high, with the removal of toxic ions. Possibly, the excretion of excess salt was done through epidermal glands. Sodium, in addition to other inorganic ions, is used for osmotic adjustment.
CONCLUSION
According to the PEI (Efficiency Index of dry matter production) methodology, accessions m11 (P. edulis f. flavicarpa), m13 (P. edulis f. flavicarpa) and m15 (P. tarminiana) were classified as salt tolerant, accessions m2 (P. edulis f. edulis), m12 (P. edulis f. flavicarpa) and m14 (P. edulis f. flavicarpa) were classified as moderately tolerant. Studies on the tolerance mechanisms of the plants that presented potential, such as stomatal closure, expression and signaling of abscisic acid at the cellular level, osmotic adjustment, and modification of gene expression, under the levels of abiotic stress are recommended.

