











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
La Amazonía colombiana con aproximadamente 45% del territorio, alberga cerca de 6500 especies vegetales y es considerada como una de las regiones con mayor diversidad del país 1. Sin embargo, la ampliación de la frontera agrícola, la ganadería extensiva, los incendios forestales y la tala selectiva para la venta de madera, constituyen los principales motores de la deforestación de bosques tropicales en el mundo, siendo que, en Colombia, se suman otras dinámicas, como la colonización y el desplazamiento de poblaciones, la minería y la siembra de cultivos ilícitos que aumentan la presión continua sobre el bosque 2. Un estudio realizado por el Sistema de Monitoreo de Bosques, del Instituto de Hidrología, Meteorología y Estudios Ambientales - IDEAM, establecieron un total de 140.356 hectáreas deforestadas en 2014 3.
Una de las principales consecuencias de la deforestación es la creación de paisajes fragmentados con tamaños y hábitats completamente transformados 4, alterando el microclima del bosque ya la vez la extinción de muchas especies, tanto a nivel local como regional 5. La fragmentación de los bosques contribuye a la pérdida de hábitats y al aislamiento de pequeñas poblaciones naturales e incrementa las probabilidades de extinción y pérdida de la biodiversidad 6. Es así que el monitoreo de grupos de organismos claves para los ecosistemas de bosques tropicales es una estrategia que ha sido utilizada para intentar entender el impacto de las alteraciones ambientales a largo plazo 7. En este sentido, la macrofauna del suelo (invertebrados mayores o iguales que 2 mm de diámetro) representa un grupo funcional que regula importantes procesos ecosistémicos en el suelo, como la descomposición de la materia orgánica, la formación de estructuras biogénicas y el reciclaje de nutrientes 8, y junto con la agregación del suelo, determinada a través de su morfología (atributos biológicos y físicos), se encuentran fuertemente relacionada con sus propiedades, como la infiltración y el almacenamiento de agua y carbono 9.
El objetivo principal de este estudio fue evaluar la composición de la comunidad de la macrofauna del suelo y su relación con la morfología del suelo en fragmentos forestales de bosques secundarios, con la finalidad de levantar informaciones que puedan auxiliar en el monitoreo de las alteraciones en los paisajes naturales con el crecimiento urbano.
Materiales y métodos
Área de estudio: El trabajo fue realizado en tres áreas de fragmentos forestales de bosque secundario denominados para este estudio como: (i) fragmento forestal de borda (FFB), cercano a la carretera; (ii) fragmento forestal conservado (FFC), con mayor diversidad de especies arbóreas; (iii) fragmento forestal de transición (FFT), localizado en un área abierta cercana de una quebrada y (iv) un área alterada de pastizal (AP). Todos los paisajes estudiados se encontraron localizados en el Centro para la Biodiversidad y el Turismo del Amazonas del Servicio Nacional de Aprendizaje - SENA Regional Amazonas, en el Municipio de Leticia, Amazonas, Colombia (Figura 1).

En el extremo sur del departamento de Amazonas, mostrando los fragmentos evaluados: fragmento forestal de borda (FFB), fragmento forestal conservado (FFC), fragmento forestal de transición (FFT) y área de pastizal (AP) en Centro para la Biodiversidad y el Turismo del Amazonas, SENA Regional Amazonas, Leticia, Amazonas, Colombia. Fuente: Google Maps, adaptado.
Autor
La vegetación predominante en los fragmentos forestales fue típica de bosque secundario en estado de regeneración, con enriquecimiento de especies arbóreas nativas de la región y el área alterada de pastizal fue cubierta por pasto de Brachiaria sp. 10 (Tabla 1).
Tabla 1 Localización geográfica y descripción general de los fragmentos forestales y área de pastizal evaluados en el Centro para la Biodiversidad y el Turismo del Amazonas, Leticia, Amazonas.

Toma de muestras: Los muestreos de la macrofauna del suelo fueron realizados durante los meses de mayo y octubre de 2017, y de acuerdo con el IDEAM 2017 11 se registró una precipitación mensual de 435 mm con una temperatura promedio de 27,17°C en mayo y una precipitación mensual de 137 mm con una temperatura promedio de 27,29°C en octubre (Figura 2).
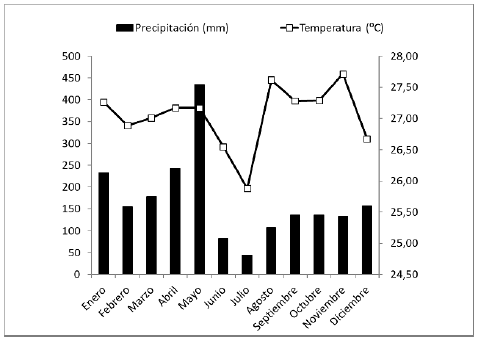
FIGURA 2 Variación de la precipitación pluvial (mm) y temperatura promedio mensual (oC) referente al año 2017, municipio de Leticia, Amazonas, Colombia. Fuente: www.ideam.gob.co (adaptado para este estudio).
Autor
El diseño experimental fue el recomendado por el protocolo detallado de muestreo de la macrofauna edáfica del proyecto Sustainable Amazonian Landscape del Centro Internacional de Agricultura Tropical (CIAT) en Palmira, Colombia 12.
La macrofauna del suelo fue evaluada con la metodología del programa de Tropical Soil Biology and Fertility (TSBF) 13, adecuado para este estudio hasta una profundidad de 20 cm. En cada una de las áreas se retiraron seis monolitos (bloques de suelo) de 25x25 cm de lado y una profundidad de 20 cm donde fueron establecidos tres estratos: hojarasca sobre el suelo y dos niveles de profundidad de 0-10 cm y 10-20 cm.
La macrofauna del suelo fue separada en el campo y conservada en frascos con alcohol al 96%. Posteriormente, en el laboratorio, los animales fueron identificados a nivel de grandes grupos taxonómicos 14 (Figura 3).

Fotos: David Preciado, Mildreth Villarreal y Sandra Celia Tapia Coral.
FIGURA 3 Secuencia del muestreo de la macrofauna del suelo, separación e identificación en campo (mayo y octubre, 2017) y proceso de identificación en el laboratorio (noviembre, 2017).
La morfología del suelo fue evaluada según la metodología descrita por Velásquez et al 15 y los muestreos del suelo fueron realizados utilizando un cuadro de metal de 10x10 cm de lado y 10 cm de profundidad en los mismos puntos donde fue muestreada la macrofauna.
El proceso de separación de los agregados del suelo fue realizado en el laboratorio, siendo que los atributos fueron separados en sus diversos componentes: (I) agregados biogénicos producidos por las lombrices, termitas, hormigas y algunas larvas de coleópteros, los cuales pueden diferenciarse por sus colores oscuros (deyecciones de lombrices principalmente), formas circulares, galerías y macroporos; (II) agregados físicos productos de compactaciones producidas por el pisoteo de animales y también, de la acción del agua, formados generalmente en suelos arcillosos y se diferencian por sus formas planas y angulares; (III) agregados producidos por las raíces de las plantas; (IV) materia orgánica (fragmentos de tallos, hojas, madera, semillas, flores, etc.); (V) suelo sin agregar, donde el material residual de la separación de los componentes es tamizado (0.5cm de diámetro); (VI) raíces de plantas y (vii) piedras 15.
Después de separados, los diversos agregados componentes del suelo fueron secados a temperatura ambiente por 72 horas y pesados en una balanza gravimétrica digital (Figura 4) 16.

Fotos: Carolina Asencio y Sandra Celia Tapia Coral.
FIGURA 4 Secuencia del muestreo de la morfología del suelo y proceso de separación de los agregados del suelo en el laboratorio (mayo y octubre 2017).
Análisis de los datos: Para evaluar el efecto de los diferentes períodos estacionales en que fueron realizados los muestreos de la macrofauna del suelo, fue realizado un análisis estadístico descriptivo seguido de la prueba t de Student con las probabilidades corregidas por la prueba de Bonferroni 17 en el programa Systat 12 18.
También, se realizó un análisis estadístico multivariado de componentes principales (ACP) con la prueba de Montecarlo 17, conforme metodología indicada en varios estudios de investigación 9,19,20,21,22. Estos análisis son realizados con el propósito de identificar los factores predominantes para las variables de las comunidades de la macrofauna del suelo (densidad ind/m2) y de la morfología del suelo (tipos de atributos) en los fragmentos forestales y en el área de pastizal. Un análisis estadístico de coinercia fue realizado para determinar correlaciones entre los vectores del análisis de componentes principales (ACP) para las variables de la macrofauna del suelo y de la morfología del suelo en las áreas estudiadas en ambos periodos de muestreo. Los análisis estadísticos multivariados fueron realizados en el software R 23 y el ACP en el paquete ADE4 24.
Resultados y discusión
Macrofauna del suelo: Fueron encontrados un total de 5332 individuos de la macrofauna del suelo distribuidos en un total de 14 grupos taxonómicos, siendo que en mayo (2329 individuos, distribuidos en 12 órdenes taxonómicas), y en octubre (3003 individuos, distribuidos en 14 órdenes taxonómicas) (Tabla 2).
TABLA 2 Abundancia de los grupos taxonómicos de la macrofauna del suelo (individuos) en mayo y octubre de 2017, en el fragmento forestal de borda (FFB), fragmento forestal conservado (FFC), fragmento forestal de transición (FFT) y área de pastizal (AP).

La abundancia total de individuos registrados en los dos períodos de muestreo no presento diferencia significativa [t=-0,75; GL=71; p(Bonferroni)=0,45], siendo en promedio, relativamente menor en mayo (x=40,6) que en octubre (x=46,9). Cuando analizamos la abundancia total de individuos en los fragmentos forestales y en el área de pastizal, entre los dos períodos de muestreo, fue encontrado una diferencia significativa únicamente para el pastizal [t=2,19; GL=17; p(Bonferroni)= 0,04], siendo en promedio, mayor en mayo (APjt=24,66) que en octubre (AP3t12,55). Para los fragmentos forestales no fueron encontrados diferencias significativas.
La riqueza total de los grupos taxonómicos registrados en los dos períodos de muestreo, no tuvo diferencia significativa [t=1,51; GL=71; p(Bonferroni)= 0,13], siendo que el promedio fue relativamente mayor en mayo (x=4,36) que en octubre (¿c=3,87). La riqueza de grupos taxonómicos de la macrofauna del suelo observada en este estudio (14 grupos taxonómicos) fue similar al reportado por Marín et al 25, que encontraron 15 grupos en sistemas de labranzas en el Valle del Cauca. Entre tanto, Botina et al 26 reportó 10 grupos en sistemas de labranza de papa en el departamento de Pasto y Suarez et al 21 reportaron 10 grupos en sistemas agroforestales en el Caquetá, Amazonía colombiana. No en tanto, Duran et al 27 reportaron hasta 21 grupos taxonómicos de la macrofauna del suelo en sistemas agroforestales con diferentes composiciones también en el Caquetá, Amazonia colombiana. Los estudios realizados en Colombia fueron enfocados en sistemas de chagras, agroforestales y cultivos con diferentes arreglos forestales en su composición 19,21,27. En la parte sur de la Amazonía colombiana aún son escasos los estudios de la macrofauna del suelo 16, siendo el presente estudio uno de los pocos realizados en áreas de fragmentos forestales de bosques secundarios.
Con relación a la distribución vertical de la densidad de la macrofauna del suelo, los datos demostraron que en el mes de mayo hubo una distribución de invertebrados predominante en la capa de hojarasca y sobre todo, en el primer horizonte del suelo (0-10 cm) en todos los fragmentos forestales y el área de pastizal (Figura 5). Indicando que durante el periodo más húmedo los primeros horizontes del suelo tienen una mayor contribución como fuente de energía y nutrientes 28.

FIGURA 5 Distribución vertical de la macrofauna del suelo (%) en el fragmento forestal de borda (FFB), fragmento forestal conservado (FFC), fragmento forestal de transición (FFT) y el área de pastizal (AP) en los dos periodos de muestreo en mayo (izquierda) y octubre (derecha) de 2017.
El segundo horizonte del suelo (10-20 cm) presentó una densidad muy baja de invertebrados en todas las áreas y en los dos periodos de muestreo, lo que puede indicar que por debajo de los 10 cm de profundidad hay una menor contribución de la actividad de la macrofauna del suelo en este horizonte.
Resultados similares a los observados en este estudio sobre la distribución vertical de la densidad de la macrofauna del suelo, fueron reportados para la Amazonía colombiana 27 y en plantíos forestales de la Amazonía peruana 29, con una mayor actividad de la macrofauna reflejada en los primeros horizontes del suelo y en la capa de hojarasca.
En octubre, durante el periodo de menor humedad, la macrofauna del suelo tuvo una mayor predominancia en el fragmento forestal conservado (FFC) y el fragmento forestal de borda (FFB) (Figura 5). Sin embargo, el fragmento forestal de transición (FFT) y el área de pastizal (AP) tuvieron una disminución considerable de la macrofauna del suelo en este período de muestreo (Figura 5), lo que puede indicar que la macrofauna fue afectada por las menores precipitaciones en este período, principalmente en el pastizal por la falta de cobertura del suelo. Resultados similares fueron reportados en otros estudios donde la comunidad de la macrofauna del suelo generalmente fue afectada en periodos con menores humidad 20,21,27.
En el fragmento forestal conservado (FFC) fue observada la mayor densidad de la macrofauna del suelo en los dos períodos de muestreos, lo que puede estar relacionado con la mayor diversidad vegetal de especies arbóreas en este fragmento 10. Otros autores 28,30 reportaron que la diversificación de plantas ofrece una mayor diversidad de microhábitats y de esta manera contribuyen a una mayor diversidad y densidad biológica del suelo.
Las lombrices de tierra (Oligochaeta) y los grupos de insectos sociales, hormigas (Formicidade) y termitas (Isoptera), fueron los más predominantes en el FFC y FFT (Tabla 2). Estos tres grupos de la macrofauna del suelo son clasificados como "ingenieros del ecosistema" 8 porque producen estructuras biológicas originadas de sus actividades, que promueven alteraciones en los atributos micromorfológicos y físicos del suelo, como la infiltración de agua y la aireación del suelo por el aumento de la porosidad. Sin embargo, estos grupos son muy sensibles a las variaciones del uso de la tierra y de la cobertura del suelo y, por lo tanto, pueden ser estudiados como indicadores de recuperación del suelo de bosques forestales 19,31.
Las termitas son organismos oportunistas y resistentes a las perturbaciones inducidas, que indican hábitats menos conservados o con algún nivel de degradación 32. En los fragmentos forestales los géneros de termitas (Isoptera) predominantes fueron: Anoplotermes sp. Cylindrotermes sp. y Nasusitermes sp.
La lombriz exótica Pontoscolex corethrurus fue predominante en todos los fragmentos forestales e inclusive en el área de pastizal. Otros estudios, también ya mostraron la predominancia de P. corethrurus en ambientes con un cierto grado de perturbación o manejo del suelo 29,33 y por lo tanto, esta lombriz es resistente y frecuente en ambientes donde el grado de perturbación es intensivo como es el caso de los pastizales.
En el segundo período de muestreo (octubre) las hormigas (Formicidae) fueron muy presentes en los fragmentos forestales, principalmente en el bosque con mayor diversidad de árboles (FFC), y poco frecuentes en el área de pastizal (Tabla 2). Las condiciones climáticas y el alimento disponible debido a la cobertura de hojarasca del suelo 20 en los fragmentos forestales, probablemente determinó una mayor actividad de las hormigas, indicando el estado de degradación del pastizal para los organismos edáficos, más afectados debido a la falta de cobertura del suelo 33.
El análisis de componentes principales (ACP) presentó una diferencia significativa (p = 0,002) en la separación de los fragmentos forestales y el área de pastizal en función de los indicadores de la macrofauna del suelo. El primer factor explicó el 22% de la variabilidad de los datos y separó principalmente el FFC de las demás áreas estudiadas. El segundo factor explicó el 18% y separó el FFT y el AP del FFC y el FFB (Figura 6).

(a) circulo de correlación de las variables: ARAN (Araneae), BLAT (Blattodea), CHIL (Chilopoda), COLE (Coleoptera), DERM (Dermaptera), DIPL (Diplopoda), FOR (Formicidae), GAST (Gasteropoda), ISOP (Isopoda), ISOPT (Isoptera), LARV (larvas), OLIGO (Oligochaeta). (b) Proyección de los fragmentos forestales: FFB (fragmento forestal de borda), FFC (fragmento forestal conservado), FFT (fragmento forestal de transición) y AP (área de pastizal) en función de los indicadores. Autor
FIGURA 6 Análisis de componentes principales (PCA) de los indicadores de la macrofauna del suelo.
El ACP también presento diferencia significativa (p = 0,001) de las variables de la macrofauna del suelo entre los dos periodos de muestreo (mayo y octubre) (Figura 7), indicando que la comunidad de la macrofauna del suelo es sensible a los cambios de precipitación en una corta escala de tiempo, como ya fue observado en otros estudios 20,21,32.

(a) Círculo de correlación de las variables de los indicadores de la macrofauna del suelo (b) Proyección de los periodos de muestreo mayo (MAY) y octubre (OCT) en función de los indicadores de la macrofauna del suelo.
FIGURA 7 Análisis de componentes principales (PCA)
Morfología del suelo. En el muestreo realizado en mayo, la composición de agregados de la materia orgánica del suelo (MO) (%) fue la más abundante en el FFC. Los agregados biológicos (AB) fueron relativamente similares en todos los fragmentos forestales muestreados y el área de pastizal en los dos periodos de muestreo. Sin embargo, en octubre se encontró una mayor cantidad de AB en el FFC y FFB; y mayores proporciones de los agregados físicos (AF) en todos los fragmentos forestales y el AP (Figura 8).
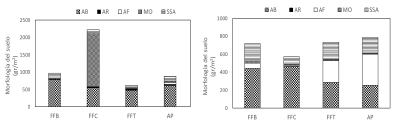
FIGURA 8 Composición de la morfología del suelo (grm2) en agregados biológicos (AB), agregados de raíz (AR),agregados físicos(AF), materia orgánica (MO) y suelo sin agregar (SSA) en el fragmento forestal de borda (FFB), fragmento forestal conservado (FFC), fragmento forestal de transición (FFT) y el área de pastizal (AP) en ambos periodos de muestreo, mayo (izquierda) y octubre (derecha).
Al analizar las variables de la morfología del suelo por medio del ACP, fue explicada un total de 58.2% de la variación en los dos primeros factores. Sin embargo, no hubo diferencia significativa entre las áreas de estudio evaluadas. Entre tanto, cuando analizamos las variables de la morfología del suelo entre los dos periodos de muestreo (mayo y octubre) encontramos una diferencia significativa (p=0.001) (Figura 9).

(a) Círculo de correlación de las variables de los indicadores de la morfología del suelo de agregados biológicos (AB), agregados de raíz (AR), agregados físicos (AF), materia orgánica (MO) y suelo sin agregar (SSA) y (b) Proyección de los periodos de muestreo mayo (MAY) y octubre (OCT) en función de los indicadores de la morfología del suelo. Autor
FIGURA 9 Análisis de componentes principales (PCA)
El análisis estadístico de coinercia no detecto correlaciones significativas entre las variables de la macrofauna del suelo y de la morfología del suelo, en los fragmentos forestales y en el área de pastizal. Sin embargo, nuestros datos demostraron diferencias na composición de la macrofauna entre los dos periodos estacionales de muestreo. El estudio de Duran et al 27 mencionó que la estructura de la comunidad de la macrofauna del suelo varía de acuerdo con el periodo de muestreo, siendo que las mínimas precipitaciones en sistemas agroforestales, resultan en una abundante capa de hojarasca en el suelo, que proporciona refugio y alimento para los macroinvertebrados. También, la densidad y diversidad de la macrofauna es afectada entre otros factores, por la composición de los sistemas 21. Sin embargo, el FFC fue el que presento mayores densidades de la macrofauna del suelo en ambos periodos de muestreo, y mayores cantidades de agregados biológicos, siendo estos AB relacionados con una mayor actividad biológica y, sobre todo, con la acción de los ingenieros del ecosistema 31.
Consideraciones finales. La macrofauna del suelo fue predominante en los primeros horizontes del suelo: hojarasca y 0-10 cm de profundidad en todos los fragmentos forestales evaluados, siendo que los grupos de insectos sociales, Isoptera y Formicidae, fueron los más predominantes en el FFC. Este fragmento presentó una mayor diversidad de composición de árboles y la mayor densidad de la macrofauna del suelo en ambos periodos de muestreo. Entre tanto, la diversidad de grupos de la macrofauna del suelo fue relativamente baja en los fragmentos de muestreo, lo que puede indicar que estas áreas tienen un cierto grado de perturbación. Por otro lado, estas áreas están en estado de regeneración forestal y, por lo tanto, son laboratorios permanentes para el desarrollo de investigaciones y prácticas de campo de los programas ambientales del centro del SENA Regional Amazonas. Resaltamos así, la importancia de los fragmentos forestales como fundamentales para el mantenimiento de la biodiversidad y de los servicios ecosistémicos del suelo, particularmente los fragmentos forestales urbanos, cada vez más afectados por el crecimiento de la ciudad de Leticia.

