











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
El estudio de foraminíferos planctónicos se ha realizado por décadas con ejemplares tridimensionales, obtenidos luego de la disgregación de rocas usando reactivos abrasivos (ej. peróxido de hidrógeno). Entre los distintos tipos de morfogrupos, tradicionalmente las conchillas trocoespiraladas obtenidas por estos métodos se han descrito taxonómicamente para su clasificación, teniendo en cuenta rasgos morfológicos reconocibles tanto en vistas laterales ya sea umbilicales o espirales (perpendiculares al eje de enrollamiento) como frontales (paralelas al eje de enrollamiento). Es decir que se basan en la vista tridimensional de los fósiles para su clasificación. El uso de este método de preparación ha logrado posicionar la micropaleontología con foraminíferos planctónicos como una de las herramientas más útiles para la datación de rocas en el registro geológico (Jones, 2014). Sin embargo, dependiendo del tipo de roca y sus procesos diagenéticos presenta limitaciones asociadas a la calidad y cantidad de recobro fósil obtenido dada la agresividad del método de preparación, la presencia de matriz adherida a la conchilla a pesar de la preparación, los procesos diagenéticos que afectaron la roca previamente (en especial procesos de disolución) y la fragilidad intrínseca de las conchillas ya sea por su tamaño, forma o composición. Dichas condiciones son comunes cuando se prospecta micropaleontológicamente las unidades del Cretácico en Colombia.
En nuestro caso de estudio se vieron replicadas estas condiciones, donde se dificultó la extracción de microfósiles en rocas calcáreas finogranulares y compactas de la Formación la Luna (Turoniano -Santoniano) y posteriormente con menor intensidad en rocas vulcanosedimentarias del Campaniano al Maastrichtiano, en el Valle Medio del Magdalena (VMM). La preparación con peróxido de hidrógeno al 8% enfatizó los procesos de disolución que las conchillas ya tenían, siendo este método poco efectivo. Por ende, las conchillas obtenidas se caracterizaron por tener un tamaño reducido, cámaras alargadas y frágiles rellenas de querógeno y rodeadas por una matriz impregnada de querógeno lo cual dificultaba la limpieza de los ejemplares. Dadas estas condiciones se reconocieron múltiples ventajas en abordar el estudio de las especies Clavihedbergella simplex y Clavihedbergella moremani mediante secciones delgadas. Esto, teniendo en cuenta que el género Clavihedbergella es fácilmente diferenciable de otros por la presencia de cámaras alargas con forma de dedos.
El uso de secciones delgadas para la clasificación de foraminíferos planctónicos es un método poco implementado en estudios bioestratigráficos hechos en Colombia y su uso frecuente es relativamente reciente fuera del país (Renz, 1936; Bolli, 1945; Robaszynski et al., 1984; Sykora et al., 1997; Jaff, 2021; Kalanat et al., 2021; Khan et al., 2021) en comparación con el método tradicional de extracción. El uso de secciones delgadas ha demostrado ser particularmente útil en ofrecer una vista de la muestra en condiciones de depósito con una abundancia y diversidad fósil representativa de la roca original. También permite la observación y el registro de conchillas frágiles que de otra forma muy posiblemente habrían sido dañadas durante el proceso de preparación convencional.
Este estudio logró identificar y registrar en sección delgada diversos ejemplares de las especies Clavihedbergella simplex y Clavihedbergella moremani, que sobresalen por sus contornos fuertemente lobulados, cámaras típicamente alargadas y delgadas. Se ofrecen herramientas para su identificación taxonómica en sección delgada, teniendo en cuenta la variabilidad morfológica intraespecífica. Además, se complementan las descripciones taxonómicas hechas tradicionalmente con ejemplares tridimensionales. Finalmente, se muestra el registro temporal de sus ocurrencias a lo largo del núcleo del pozo A, ubicado en la parte central del Valle Medio del Magdalena (Colombia) (Figura 1).
Marco conceptual
Tipos de corte en una conchilla de foraminífero planctónico trocoespiralado
La sistemática taxonómica de foraminíferos planctónicos se ha basado tradicionalmente en ejemplares fósiles tridimensionales. Sin embargo, en la mayoría de los casos y sin duda en el caso de las especies Clavihedbergella simplex y Clavihedbergella moremani, las características propias de cada especie pueden ser reconocidas en ejemplares bidimensionales.
Los mejores cortes para el reconocimiento bidimensional de foraminíferos planctónicos son los cortes frontales o axiales (Figura 2). Este tipo de corte es paralelo al eje de enrollamiento de la conchilla y en él se observa con claridad: 1) la altura de la espira, 2) el ancho y profundidad del ombligo, 3) la presencia de quillas, 4) crestas umbilicales y estructuras aperturales (ocasionalmente), 5) el grosor de la conchilla, 6) la forma y tamaño de las cámaras distales y proximales donde la diferencia en tamaño es un indicador de la rapidez del crecimiento de la conchilla.
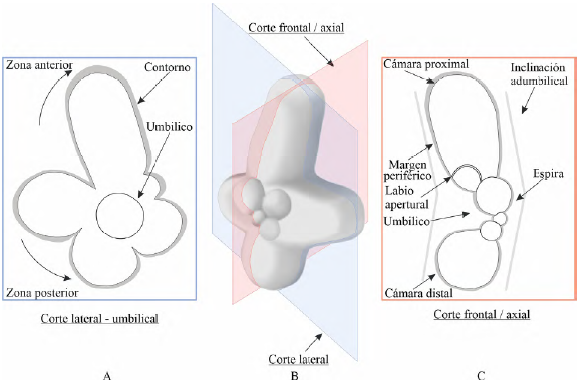
Figura 2 Tipos de corte útiles para la clasificación de foraminíferos planctónicos trocoespiralados en sección delgada y descripción de estructuras diagnósticas. A. Vista de un corte lateral del lado umbilical del ejemplar tridimensional. Las cámaras de la zona posterior y anterior se ubican respecto a la última cámara. La silueta demarcada por las cámaras de la última vuelta se ha coloreado de gris y se conoce como "contorno". B. Vista tridimensional del ejemplar seccionado por los planos de corte señalados. C. Vista de un corte frontal o axial del ejemplar. Las cámaras proximal y distal se ubican con respecto a la apertura principal. La silueta demarcada por los límites externos de la conchilla se ha resaltado con un color gris y se conoce como "margen periférico".
En segundo orden de relevancia, los cortes laterales o transversales (perpendiculares al eje de enrollamiento) ya sea de la vista espiral o umbilical, son especialmente útiles en la diferenciación de especies según la cantidad de cámaras existentes en la última vuelta, además de confirmar la rapidez de crecimiento cameral. Ocasionalmente podrán observarse las variadas presentaciones suturales tanto del lado espiral como umbilical e incluso aperturas secundarias.
Metodología
La observación cuidadosa de 109 secciones delgadas provenientes del pozo A (Valle Medio del Magdalena, Colombia) permitió reconocer algunas especies del género Clavihedbergella a lo largo de una sucesión de 465 metros del Cretácico tardío. La sucesión rocosa corresponde a la Formación la Luna (en su definición litoestratigráfica tradicional) y a una unidad vulcanosedimentaria depositada durante el Campaniano y el Maastrichtiano. Las secciones delgadas contaban con una tinción combinada de rojo de alizarin y ferricianuro de potasio. De las secciones delgadas se tomaron fotografías con un microscopio petrográfico y la escala se referenció con una barra equivalente a 100 μm en cada fotografía. Para cada sección analizada, los ejemplares de las especies Clavihedbergella simplex y Clavihedbergella moremani fueron fotografiadas en un microscopio petrográfico Motic BA310Pol (Figuras 3 a 5), luego fueron clasificadas y registradas. La clasificación de cada fósil parte de las descripciones taxonómicas realizadas de los holotipos (ejemplares tridimensionales) disponibles en la base de datos Mikrotax (Young et al., 2017). Posteriormente las descripciones taxonómicas se complementaron con observaciones de ejemplares bidimensionales (en sección delgada) obtenidos en este estudio. Finalmente, teniendo en cuenta los registros de ocurrencia para las diferentes especies, se graficó la presencia de cada especie a lo largo del núcleo del pozo A (Figura 6) y según la identificación de especies guía que permitieran el reconocimiento de biozonas, se asignaron los rangos temporales. En orden cronológico, las biozonas que contienen eventos de primer y última ocurrencia para Clavihedbergella simplex y Clavihedbergella moremani fueron: la biozona de Rotalipora cushmani definida por la última ocurrencia de Thalmanninella reicheli y la última ocurrencia de Rotalipora cushmani (Coccioni y Premoli- Silva, 2015), la biozona de Helvetoglobotruncana helvética (Dalbiez, 1955 en Sliter, 1989) definida por la primer y última ocurrencia de Helvetoglobotruncana helvetica y la biozona informal de Trinitella scotti. Dicha especie fue usada como indicador bioestratigráfico inicialmente por Masters (1977 en Arz y Molina, 2002) y posteriormente fue reconocida como el evento de foraminíferos más cercano a la sección estratotipo y punto de límite global (en inglés GSSP), en la base del Maastrichtiano por Odin y Lamaurelle (2001). En este estudio la biozona informal de Trinitella scotti se define por la primera ocurrencia de dicha especie y la primer ocurrencia de Pseudoguembelina palpebra.
Resultados obtenidos
Taxonomía
Se adopta el esquema taxonómico subordinal implementado en Mikrotax (Young et al., 2017) Familia Hedbergellidae Loeblich y Tappan, 1961 Género ClavihedbergellaBanner y Blow, 1959 Especie Clavihedbergella simplexMorrow, 1934 Especie Clavihedbergella moremaniCushman, 1931
ClavihedbergellaBanner y Blow, 1959
Corte axial: Conchilla trocoespiral de espira baja y con apariencia comprimida. En la última vuelta, la cámara distal es esférica a subglobular mientras que la cámara proximal es alargada y puede o no tener un mayor grosor hacia la periferia en comparación con la base. A mayor diferencia de tamaño entre la cámara distal y proximal, mayor será la velocidad de crecimiento de la conchilla en la vuelta determinada. La velocidad de crecimiento en la última vuelta oscila entre moderada a muy alta, y la diferencia de velocidad de crecimiento entre la penúltima y la última vuelta es alta. La pared de la conchilla tiene un grosor reducido y dependiendo de la conservación pueden verse poros reducidos con distribuciones regulares. La conservación de la estructura apertural con un labio corto es ocasional (Figura 2C).
Corte lateral umbilical / espiral: Conchilla de contorno lobulado. En la última vuelta las cámaras posteriores son globulares a subglobulares, mientras que las cámaras anteriores son elongadas en dirección radial, respecto del umbilico o el centro de la espira (excepto en Clavihedbergella subdigitata). La superficie puede verse finamente irregular o lisa (Figura 2A).
Las conchillas vistas en sección delgada pueden ser clasificadas esencialmente por la morfología y disposición de sus cámaras, pero características como el labio y las perforaciones en la conchilla no suelen ser evidentes. Las conchillas vistas en cortes axiales son reconocidas por la elongación de las cámaras a nivel de género, sin embargo, este tipo de cortes en Clavihedbergella subdigitata y Clavihedbergella subcretacea son muy similares y es necesario tener el corte lateral para identificar la especie.
Clavihedbergella simplex (Morrow, 1934)
Hastigerinella simplex.Morrow, 1934, pl 30, fig 6. Holotipo (3D)
Clavihedbergella simplex.Loeblich y Tappan, 1961, pg 285, pl 3, fig 11-14. (3D)
Clavihedbergella simplex.Eicher y Worstell, 1970, pg 317, pl 10, fig 4, 6. (3D)
Clavihedbergella simplex.Premoli-Silva y Bolli, 1973, pg 535, pl 4, fig 2-4. (3D)
Clavihedbergella simplex.Caron, 1978, pg 673, pl 8, fig 1,2; pg 678, pl 11, fig 3,4. (3D)
Clavihedbergella simplex.Miles y Orr, 1980, pg 809, pl 4, fig 8-10. (3D)
Hedbergella simplex.Bolli et al., 1989, pg 58, fig 25:15-16. (3D)
Clavihedbergella simplex.Sliter, 1999, pg 335, pl 3: 14, 18. (2D)
Muricohedbergella simplex.Premoli-Silva y Verga, 2004, pg 168, pl 98:3-4. (3D)
Muricohedbergella simplex.Premoli-Silva y Verga, 2011, pl 30: 15; pl 31:1-3 (2D)
Autor:Morrow, 1934.
Descripción: vista en un corte axial, es una conchilla claramente trocoespiral de espira baja a muy baja, con una cámara distal esférica a globular y una cámara proximal gruesa, dos a tres veces más elongada que la cámara distal (es decir, que tiene un crecimiento muy rápido) y con una terminación redondeada hacia la periferia. El umbilico que separa estas dos cámaras es usualmente amplio y superficial y rara vez se conserva la estructura apertural de un labio delgado que sobresale discretamente en la base de la cámara proximal del lado umbilical. Vista en corte lateral es una conchilla de contorno fuertemente lobulado con 4 a 5, rara vez 6 cámaras en la última vuelta. Las cámaras posteriores de la última vuelta son globulares. Hacia la parte anterior de la conchilla la penúltima cámara es ligeramente elongada, mientras que la cámara final está fuertemente alargada y tiene una terminación redondeada hacia la periferia.
Rango: según Mikrotax (Young et al., 2017) el rango es Aptiano tardío a Coniaciano tardío. Primera ocurrencia (PO): Biozona de Microhedbergella renilaevis. Última ocurrencia (UO): Biozona de Dicarinella concavata. Según los registros en el pozo A el rango es Cenomaniano temprano a Maastrichtiano temprano (Figura 6). PO: Base de la biozona de Rotalipora cushmani. UO: Biozona informal de Trinitella scotti con rango de intervalo, definido por la primera ocurrencia de Trinitella scotti y la primera ocurrencia de Pseudoguembelina palpebra.
Comentarios: la variabilidad intraespecífica está mediada por el grado de compresión de la conchilla y el grosor de las cámaras proximales, evidentes en las Figuras 3 y 4. Vista en corte axial se diferencia de las especies Clavihedbergella subdigitata y Clavihedbergella subcretacea en que las cámaras de la primera vuelta están claramente ubicadas contra uno de los márgenes periféricos o sobresale ligeramente de él. Esto puede o no darle una ligera inclinación adumbilical al margen periférico, mientras que para Clavihedbergella subdigitata el margen periférico es típicamente paralelo y para Clavihedbergella subcretacea el margen periférico puede inclinarse ligeramente hacia la espira.
La presencia de Clavihedbergella simplex inicia en la base de la Formación La Luna siendo abundante en el Miembro Salada. Su presencia es escasa a nula en gran parte de los miembros Pujamana y Galembo. Hacia el tope del Miembro Galembo presenta un incremento súbito, continua su existencia en rocas vulcanosedimentarias del Campaniano y la parte basal del Maastrichtiano. Cabe resaltar que por lo general los ejemplares registrados durante el Campaniano y Maastrichtiano tienen un tamaño reducido y cámaras que poco se ensanchan en comparación con los ejemplares vistos durante el Turoniano (Miembro Salada).
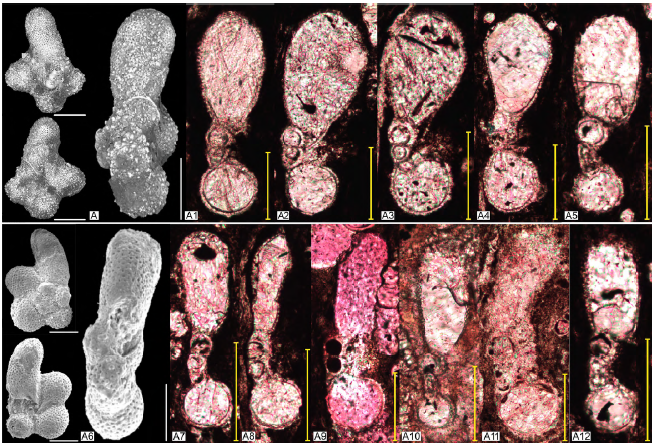
Figura 3 Ejemplares de la especie Clavihedbergella simplex. A. Hastigerinella simplex (Clavihedbergella simplex). Holotipo. Morrow, 1934 (USNM PAL 75376 en Mikrotax (Young et al., 2017). A1 - A5. Clavihedbergella simplex en corte axial. A1. Profundidad 48,53 metros. Zona Helvetoglobotruncana helvetica. A2. Profundidad 49,29 metros. Zona Helvetoglobotruncana helvetica. A3. Profundidad 64,41 metros. Zona Helvetoglobotruncana helvetica. A4. Profundidad 42,86 metros. Zona Helvetoglobotruncana helvetica. A5. Profundidad 70,63 metros. Zona Helvetoglobotruncana helvetica. A6. Muricohedbergella simplex (Clavihedbergella simplex). Premoli-Silva y Verga, 2011. Pl 98. A7-A12. Clavihedbergella simplex en corte axial. A7. Profundidad 46,21 metros. Zona Helvetoglobotruncana helvetica. A8. Profundidad 73,59 metros. Zona Helvetoglobotruncana helvetica. A9. Profundidad 313,28 metros. Zona Dicarinella asymetrica. A10. Profundidad 350,02 metros. Zona Contusotruncanaplummerae. A11. Profundidad 388,70 metros. Zona Trinitella scotti. A12. Profundidad 70,63 metros. Zona Helvetoglobotruncana helvetica. Escala: 100 μm.
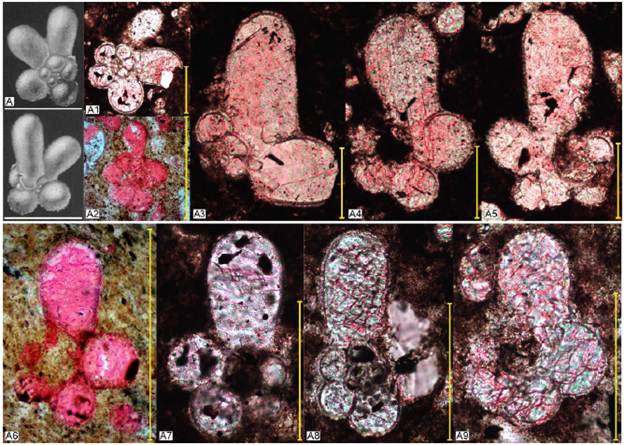
Figura 4 Ejemplares de la especie Clavihedbergella simplex. A. Clavihedbergella simplex.Loeblich y Tappan, 1961. Pg 285, pl 3, fig 12-14. A1-A9. Clavihedbergella simplex. A1. Profundidad 76,24 metros. Zona Helvetoglobotruncana helvetica. A2. Profundidad 358,71 metros. Zona Globotruncanella havanensis. A3, A4, A5. Profundidad 61,49 metros. Zona Helvetoglobotruncana helvetica. A6. Profundidad 423,09 metros. Zona Trinitella scotti. A7, A8, A9. Profundidad 76,24 metros. Zona Helvetoglobotruncana helvetica. Escala 100 μm.
Clavihedbergella moremani (Cushman, 1931)
Hastigerinella moremani.Cushman, 1931, pg 85, pl 11, fig 1-3. Holotipo. (3D)
Clavihedbergella moremani.Pessagno, 1967, pg 393, pl 53, fig 5. (3D)
Clavihedbergella moremani.Eicher y Worstell, 1970, pg 317, pl 10, fig 5. (3D)
Clavihedbergella moremani.Premoli-Silva y Bolli, 1973, pg 539, pl 6, fig 1. (3D)
Clavihedbergella moremani.Miles y Orr, 1980, pg 809, pl 4, fig 4-7. (3D)
Clavihedbergella moremani.Leckie, 1984, pg 611, pl 8, fig 11-12. (3D)
Autor:Cushman, 1931.
Descripción: vista en un corte axial, es una conchilla claramente trocoespiral de espira muy baja, con la última vuelta planispiral. La cámara distal es esférica a globular y ligeramente alargada, mientras la cámara proximal es tres o cuatro veces más larga (indicando un crecimiento muy rápido). La cámara proximal tiene una base medianamente ancha, una parte media más estrecha y una terminación muy amplia y redondeada. El umbilico es estrecho y superficial. Observando ejemplares con buena preservación, la pared de la conchilla se reconoce macroperforada (Figura 5). No se observó la conservación de estructuras aperturales. En vista lateral es una conchilla fuertemente lobulada, con 5 a 6 cámaras en la última vuelta. Las cámaras posteriores (cámaras iniciales de la última vuelta) son globulares, mientras que en sentido proximal (cámaras finales de la última vuelta) las cámaras son más elongadas y clavadas. Las últimas cámaras son muy alargadas y tienen una terminación redondeada y expandida.
Rango: según Mikrotax (Young et al., 2017) el rango es Turoniano medio a Coniaciano tardío. PO: Biozona de Marginotruncana schneegansi. UO: Biozona de Dicarinella concavata. Según los registros en el pozo A el rango es Turoniano temprano (Figura 6). PO: Biozona de Helvetoglobotruncana helvetica. UO: Biozona de Helvetoglobotruncana helvetica.
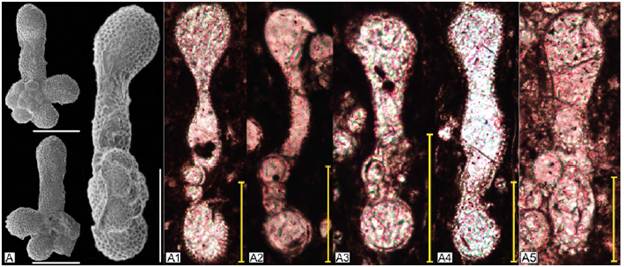
Figura 5 Ejemplares de la especie Clavihedbergella moremani. A. Hastigerinella moremani (Clavihedbergella moremani). Holotipo. Pessagno, 1967 en Mikrotax (Young et al., 2017). A1-A4. Clavihedbergella moremani en corte axial. A1, A3, A4. Profundidad 48,9 metros. Zona Helvetoglobotruncana helvetica. A2. Profundidad 51,8 metros. Zona Helvetoglobotruncana helvetica. A5. Ejemplar de transición entre Clavihedbergella simplex a Clavihedbergella moremani en corte axial. Profundidad 24,4 metros. Zona Rotalipora cushmani. Escala 100 μm.
Comentarios: este estudio publica los primeros cortes axiales en sección delgada de Clavihedbergella moremani en Colombia. En las secciones delgadas estudiadas sólo se observaron cortes axiales de esta especie y en ese sentido se conservan las descripciones en ejemplares tridimensionales para las características evidentes únicamente en cortes laterales.
La presencia de la especie Clavihedbergella moremani inicia y termina en el Miembro Salada de la Formación La Luna, abarcando un espesor estratigráfico reducido (~27 metros).
Discusión de resultados
La variabilidad morfológica de Clavihedbergella simplex se observa en el grado de compresión de la conchilla vista en un corte axial y el grosor de las cámaras más alargadas (Figura 3). Los rasgos morfológicos de esta especie son tan particulares que en varios casos podría clasificarse en sección delgada aún con ejemplares incompletos (Figura 4A3).
Las conchillas de Clavihedbergella simplex vistas en rocas del Turoniano son más grandes y sus cámaras son infladas hacia la periferia, mientras que aquellas conchillas del Santoniano tardío al Maastrichtiano temprano son comparativamente más pequeñas, comprimidas y con cámaras de un grosor uniforme (Figura 3). Esta información es relevante por dos motivos: 1) Usualmente el rango temporal para esta especie es Aptiano tardío a Santoniano (Young et al., 2017), pero en estas rocas la especie tiene un rango registrado de Turoniano a Maastrichtiano temprano. 2) La variabilidad morfológica de la especie durante el Santoniano - Maastrichtiano temprano probablemente obedecería a condiciones ambientales menos favorables para su desarrollo en comparación con aquellas presentes durante el Turoniano. Respecto a la mayor amplitud del rango temporal de Clavihedbergella simplex en estas rocas, se sugiere que la actividad volcánica registrada durante el Campaniano en esta parte de la sección, junto con el contexto regional de cuenca intracontinental para este momento (Villamil et al., 1999) podrían ser factores que favorecieron condiciones anóxicas a nivel local, y por ende permitieron la preservación de este grupo fósil más allá del Santoniano. Estudios adicionales en diferentes partes de la cuenca serían necesarios para corroborar dichas hipótesis.
Continuando, la presenciade laespecie Clavihedbergella simplex en las rocas estudiadas del pozo A (Figura 6) tiene sus inicios en la base de la Formación La Luna, en el Miembro Salada (Turoniano) y suele tener mayor abundancia en estratos con un alto contenido de materia orgánica. La presencia de materia orgánica indicaría bajos niveles de oxígeno en los sedimentos de fondo, y es en estas condiciones que se observa mayor abundancia de Clavihedbergella simplex. Las unidades con mayor abundancia de esta especie fueron el Miembro Salada, el Miembro Galembo y en menor proporción las rocas vulcanosedimentarias del Campaniano con procesos intensos de fosfatización. Esto corroboraría que los episodios de deficiencia de oxígeno en el mar se relacionan con el favorecimiento de morfologías con cámaras alargadas (Kopaevich y Gorbachik, 2017). Se cree que dichas condiciones de anoxia fueron más intensas durante el Turoniano en medio de un evento anóxico oceánico (EAO2) (Monteiro et al., 2012) que durante el Campaniano en medio de eventos volcánicos locales (denotado por la presencia de tobas) y fenómenos de surgencia (fosfatización intensa), lo cual puede explicar que las conchillas estén más desarrolladas durante el Turoniano que durante el Santoniano hasta la base del Maastrichtiano.
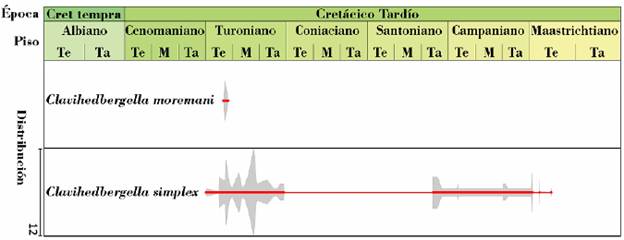
Figura 6 Rango temporal para las especies estudiadas. En la fila superior se detalla la época, piso/edad y en las filas posteriores se representa gráficamente la distribución y abundancia absoluta de las especies Clavihedbergella simplex y Clavihedbergella moremani vistas en secciones delgadas. La línea roja y gruesa indica el registro certero de la especie, mientras que la línea roja y delgada indica una presencia asumida de la especie. La abundancia de cada especie se representa con una curva cerrada de color gris cuya amplitud tiene como referencia una escala (esquina inferior izquierda) cuyo máximo valor equivale a 12 ejemplares y el mínimo equivale a 0 (una línea simple). La especie Clavihedbergella simplex tiene su primer registro en 30,61 metros y su último registro en 474,9 metros. La especie Clavihedbergella moremani tiene su primer registro en 24,42 metros y su último registro en 51,79 metros. Las profundidades en este estudio son propias y están invertidas respecto a la superficie. La figura no ha sido representada a escala.
Por su parte, los registros de Clavihedbergella moremani en el pozo A inician y finalizan en la parte inferior del Miembro Salada (Turoniano temprano) y es particularmente notorio que esta especie tiene un rango más reducido que la biozona que lo contiene, por lo cual se sugiere realizar estudios regionales que permitan estimar su utilidad como un fósil guía y su uso para correlacionar secciones estratigráficas del Miembro Salada de la Formación La Luna a lo largo de la cuenca.
Conclusiones
Las especies de foraminíferos planctónicos Clavihedbergella simplex y Clavihedbergella moremani fueron descritas en cortes de sección delgada, y su presencia registrada a lo largo de rocas Cretácicas perforadas en el pozo A, ubicado en el Valle Medio del Magdalena.
La especie Clavihedbergella simplex presenta una gran variabilidad intraespecífica y es fácilmente reconocible en cortes de sección delgada. Tiene su primera ocurrencia en la base de la Formación La Luna - Miembro Salada (Turoniano) y la base de la biozona de Rotalipora cushmani. Su última ocurrencia se registró en rocas fosfatizadas de edad Maastrichtiano temprano, en la biozona informal de Trinitella scotti. El inusual registro de esta especie en rocas más recientes que el Santoniano probablemente se deba a fenómenos locales, donde su mayor abundancia ocurre en rocas del Miembro Salada, y en menor medida, en rocas del Miembro Galembo y rocas vulcanosedimentarias de edad Campaniano. A su vez, dicha especie muestra una variabilidad morfológica asociada probablemente a los niveles de anoxia en el ambiente.
Se presenta el primer corte en sección delgada de la especie Clavihedbergella moremani en Colombia (Miembro Salada). Esta especie tiene un rango bioestratigráfico muy corto, está incluida en la biozona de Helvetoglobotruncana helvetica (Turoniano temprano) y podría ser de utilidad para el estudio bioestratigráfico de la Formación La Luna.

