










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. v.42 n.2 Bogotá 2016
Development and reproductive performance of Chrysoperla externa (Neuroptera: Chrysopidae) using preys from wheat crop
Preferencia, desarrollo y desempeño reproductivo de Chrysoperla externa (Neuroptera: Chrysopidae) utilizando presas provenientes de trigo
JULIANA PITWAK1,2, AYRES OLIVEIRA MENEZES JR.1,3 and MAURÍCIO URSI VENTURA1,4
1 Universidade Estadual de Londrina, Campus Universitário, Departamento de Agronomia, CCA, PO BOX 6001, 86051 970, Londrina, PR, Brazil.
2 Ms. Sc.Agronomy, jupitplant@yahoo.com.br.
3 Professor, D. Sc. in Entomology, res@uel.bray.
4 Professor, D. Sc. in Entomology. mauricioursiventura@gmail.com, corresponding author.
ABSTRACT
Biological parameters of Chrysoperla externa were assessed in laboratory. The effect of the preys on biological parameters and reproductive performance were assessed when the following food treatments were provided: Sitobion avenae in all larval instars; Rhopasosiphum padi in all larval instars; Anagasta kuehniella eggs in the first and second instars and Dichelops melacanthus eggs in the third; R. padi in the first and second instars and D. melacanthus in the third instar. All foods provided satisfactory development and reproductive performance. Changing larvae food from A. kuehniella to D. melacanthus did not affect its survival. However, changing larvae food from R. padi to D. melacanthus reduced survival. When larvae were fed with R. padi (1st and 2nd instars) and D. melacanthus (3rd instar), C. externa laid more eggs than when larvae were fed only with aphids. Other parameters either were not affected, or were little affected by the diet change. Results showed that aphis are important food sources for C. externa, and this predator may also predate and develop on D. melacanthus eggs.
Key words: Aphids. Biological control. Lacewing. Integrated pest management. Stink bug.
RESUMEN
Se evaluaron los parámetros biológicos de Chrysoperla externa en condiciones de laboratorio. El efecto de las presas sobre los parámetros biológicos y el comportamiento reproductivo fueron evaluados cuando se proporcionaron los siguientes alimentos: Sitobion avenae; Rhopasolsiphum padi (ambos en los tres estadios larvales); huevos de Anagasta kuehniella del primero al segundo estadio y huevos de Dichelops melacanthus en tercer estadio; R. padi del primero al segundo estadio y huevos de D. melacanthus en tercero. Todos los alimentos determinaron desarrollo y desempeño reproductivo satisfactorios. El cambio de presa de A. kuehniella a D. melacanthus no afectó la viabilidad de las larvas, pero sí la redujo el cambio de R. padi a D. melacanthus. Cuando las larvas se alimentaron con R. padi (primer y segundo estadio) y D. melacanthus (tercer estadio), C. externa puso más huevos que cuando se proporcionaron los áfidos. Como consecuencia del cambio de dieta, otros parámetros no fueron afectados o sólo ligeramente. Los resultados mostraron que los áfidos son importantes fuentes de alimento para C. externa y que este depredador también puede consumir y desarrollarse con huevos de D. melacanthus.
Palabras clave: Áfidos. Control biológico. Crisópidos. Manejo integrado de plagas. Chinche.
Introduction
Chrysopidae family includes some common predator species that occur in agroecosystems, and they play important role in regulating populations of phytophagous organisms. These species prey on eggs, neonate larvae, aphids, mealybugs, mites and other arthropods of small size and of easily penetrable tegument (Carvalho & Souza 2009). Propositions of using lacewings in integrated management programs include Alabama argillacea (Hubner, 1818) (Lepidoptera: Noctuidae); aphids Aphis gossypii (Glover, 1877), Schizaphis graminum (Rondani, 1852) and Rhodobium porosum (Sanderson, 1900) (Hemiptera: Aphididae); some citrus mealybugs Coccus sp (Hemiptera: Coccidea), Orthezia sp. (Hemiptera: Orthezidae) Pinnaspis sp. and Selenaspidus sp. (Hemiptera: Diaspididae); thrips Enneothrips flavens Moulton (Rodrigues et al. 2014) and Thrips tabaci Lind. (Thysanoptera: Thripidae) (Abbas et al. 2012); leafhopper Amarasca devastans Dist. (Hemiptera: Cicadellidae) (Abbas et al. 2012); and rubber tree lacebug Leptopharsa heveae Drake & Poor, 1935 (Hemiptera: Tingidae) (Carvalho & Souza 2009).
Chrysoperla externa (Hagen, 1861) (Neuroptera: Chrysopidae) occurs in superior populations in wheat crop and has been reported feeding and developing on several aphid species (Fonseca et al. 2000; Costa et al. 2002; Macedo et al. 2010; Costa et al. 2012). However, studies on the biology of this predator feeding on wheat aphids preys were not found. These studies are important since some aphid species, such as the citrus pest Toxoptera citricidus (Kirk, 1907) (Hemiptera: Aphididae), do not provide satisfactory conditions for C. externa development, despite the abundance of the predator in citrus orchards (Godoy et al. 2004). In addition, although C. externa presents several traits that may allow its use in biological control (Albuquerque et al. 1994).
In integrated pest management (IPM) programs, aphid populations may have a positive effect in attracting and maintaining natural enemies in crops (Gravena 1992). Besides, these natural enemies may keep other pest populations below damage threshold. Rhopasolsiphum padi (L.) and Sitobion avenae (Fabricius) (Hemiptera: Aphididae) are ordinary abundant preys for C. externa in wheat fields. In the field, C. externa larvae were observed also predating green belly stink bug Dichelops melacanthus (Dallas, 1851) (Hemiptera: Pentatomidae) eggs (Pitwak J., personal observation). This seed sucker is reported as a soybean secondary pest. Due to the massive use of no tillage system, and to late cropping maize (autumn cropping) in Brazil, D. melacanthus becomes a primary pest of wheat and maize, damaging the seedlings of these crops. This species is favored by crop debris, which provide shelter and food (soybean grains not collected by the combine harvester). Although D. melacanthus does not develop feeding on wheat or maize plants, it does on tropical spiderwort (Commelina benghalensis) that occurs in these fields and contributes to the high occurrence of the pest (Silva et al. 2013).
We studied the capacity of C. externa in feeding and developing on some wheat pests. To assess the effect of these pests on biological parameters, four preys or combinations were provided as food: a) S. avenae in all larval stages of C. externa; b) R. padi in all larval stages of C. externa; c) Anagasta kuehniella (Zeller, 1879) (Lepidoptera: Pyralidae) eggs [which is considered standard food for C. externa (Carvalho & Souza 2009)] in the first and second instars and D. melacanthus eggs in the third; and d) R. padi in the first and second instars and D. melacanthus in the third instar.
Material and methods
Chrysoperla externa adults were collected from a wheat field in the Universidade Estadual de Londrina farm school (23°19';S, 51°12';W) and kept in an environmental chamber (25 °C, 70 % RU and 12:12 L:D). Adult cages were made using PVC tubes (20 cm height x 10 cm diameter) covered with white bond paper. Yeast and honey (1:1), together with water, were provided as food. Eggs were collected in the rearing cages when they reached 24 h of age. Pedicel was removed using fine-tipped scissors and placed in microtitration plates sealed with laminated PVC film. Newly hatched larvae were transferred to plastic vessels (3 cm diameter x 1,7 cm height) in which preys were available.
The biology of C. externa larvae and adults were investigated in four food combinations: S. avenae (Sa) in all larval instars of the predator; R. padi (Rp) in all larval instars of the predator; A. kuehniella eggs in the first and second instar and D. melacanthus eggs in the third instar (Ak + Dm); and R. padi in the first and second instar and D. melacanthus eggs in the third instar (Rp + Dm). The treatment A. kuehniella in the two first instars, and D. melacanthus, in the third instar, were used to simulate occasional laboratory rearing and predator field release when the predator could feed on eggs. R. padi (two first intars) + D. melacanthus (third instar) is probably an ordinary situation in the field. Preys were made available ad libitum in rearing cages. Cages were observed twice a day to provide food without limitation. Aphids were reared in wheat plants cv. Alcover in greenhouse. A. kuehniella eggs not sterilized were obtained from rearing facilities in the IAPAR- Instituto Agronômico do Paraná. Seven replications with six larvae per replication were used.
After emergence, adults lacewings were kept in refrigerator (9 °C) in order to reduce movements, and were weighted (mg) and separated by sex, by external genitalia observation. Couples were kept inside PVC cages (10 x 10 cm), using the method proposed by Ribeiro (1988). Yeast mixed with honey and water were provided as food. For adults, ten randomly chosen couples were used per treatment. Larvae and adults were removed from container and weight using an analytical balance (resolution 0.0001 g) (Bel Engineering, Italy) after kept in refrigerator (9 °C) in order to reduce movements. Duration and survival of larvae, prepupae (larvae cease feeding to make the cocoon) and pupae, and weight (mg) after 24 h in each stage were assessed. For adult stage, it was evaluated weight, pre-oviposition, oviposition (interval between the first and last laying), effective oviposition (number of days in which females oviposited) and post-oviposition periods; total number of eggs, and longevity and survival of eggs .
Completely randomized design was used. To verify the assumptions for the analysis of variance, tests of the variance homogeneity and normality were carried out. Thereafter, analysis of variance was carried out, and Tukey test was used to compare means. Variables percentage were transformed using the constant arc sen â(x/100). SASM - Agri (Canteri et al. 2001) software package was used.
Results and discussion
In general, results showed that preys used in the present study provided suitable nutritional quality for C. externa development and reproduction since unsuitable nutritional quality causes negative effects in the biology of the predator. Duration of the second instar was shorter than in the other ones, and Sa was the longest (Table 1). In the third instar, Ak + Dm was shorter than the others. The values reported here are quite similar to those reported when C. externa was fed on Sitotroga cerealella (Olivier, 1789) (Lepidoptera: Gelechiidae) eggs: 3.04 (± 0.02), 2.55 (± 0.08) and 3.67 (± 0.07) days, respectively (Costa et al. 2002); or A. gossypii, 3.89 (± 0.06), 2.55 (± 0.09) and 4.19 (± 0.07) (Ribeiro 1988) for the first, second and third instars, respectively. The periods found for 2nd and 3rd instars in the S. avenae treatment were similar to those found previously for the aphid Neotoxoptera (± 0.14) and 6.0 (± 0.15) days, respectively] (Costa et al. 2012).

Duration of larval stage was longer when C. externa was fed on Sa than with the remainder treatments (Table 2). The shortest period was found for Ak + Dm. Similar duration was previously reported for C. externa when reared using S. cerealella eggs (9,18 days ) (Costa et al. 2002) or the aphid A. gossypii [10.62 or 10.28 days (Costa et al. 2002; Ribeiro 1988)]. A longer duration of larval period was found for the aphid N. formosana [(14.1 days (± 0.38)] (Costa et al. 2012).

Pre-pupae period was shorter in the treatment Rp than in the other treatments (Table 2). Ak + Dm treatment showed shorter pupae period than the other ones. Total period from larvae to pupae was longer in Sa than in the other treatments (Table 2). Similar results were reported to C. externa reared on S. cerealella eggs and A. gossypii (2.68 and 3.0; 7.01 and 6.73; 19.3 and 20 days to pre-pupae, pupae and larvae to pupae, respectively) (Costa et al. 2002).
Larvae fed with Rp + Dm presented lower larval survival than Ak + Dm and Sa (Table 3). Similar result was found for pre-pupae and pupae survival among treatments. In the second instar, C. externa larvae that feed on Ak + Dm presented lower weight when compared to the other treatments. In the third instar, C. externa larvae that fed on Sa were heavier than other treatments (Table 4). However, when insects reached pupae stage, similar weights were recorded. These results were quite similar to those found when C. externa was reared feeding on Bemisia tabaci (Gennadius, 1889) (Hemiptera: Aleyrodidae) originating from milkweed (Euphorbia heterophylla L.) (Linnaeus) plants (Silva et al. 2004b). However, in the same study, when B. tabaci was reared on green collard (Brassica oleracea L.) (Linnaeus) plants, greater weights on the third instars [1.00 (± 0.15), 3.57 (± 0.22) mg, respectively)] were found when compared to that found in this study [2.50 (± 0.10)] (Table 4). Satisfactory pupae weight represents suitable ovaries development, and thus oviposition capacity. In general, females were heavier than males (Fig. 1A), which contradicts earlier results in which similar weights for males and females were found when larvae were fed with B. tabaci nymphs (Silva et al. 2004a). Adults weights are important traits associated with higher egg production (Carvalho & Souza 2009).



The higher survival in Ak + DM than than Rp + Dm (Table 3) suggests that the predator adapted itself more quickly to feed on Dm eggs when they had been previously fed on A. kuehniella eggs. Otherwise, the high longevity in the treatment Ak + Dm suggests suitable nutritional value of Dm eggs. High survival of C. externa larvae when feeding on several preys was also previously reported (Ribeiro 1988; Costa et al. 2002). Predators, in general, can consume more than one kind of prey during their development, which contributes to fulfill nutritional requirements for C. externa development (Fonseca et al. 2000; Ecole et al. 2002).
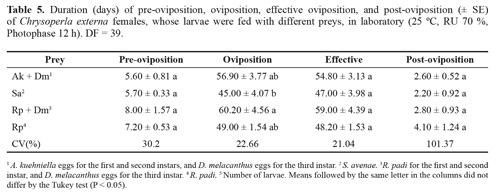
Longer oviposition period was found for larvae fed with Rp + Dm than for those fed with Sa (Table 5). Similar pre, effective and post-oviposition periods were observed among treatments. Higher longevity was recorded for males reared on Rp + Dm than on the other treatments (Table 6). Females reared on Rp + Dm also showed higher longevity than on Sa treatment. In previous studies, adults longevity (males and females recorded together) varied quiet similarly to the present results, ranging from 34.96 (± 7.60) to 69.75 (± 4.21), depending on the host in which the prey was developed (Silva et al. 2004a).

The number of eggs produced by females whose larvae were fed with Rp + Dm was higher than those fed with Sa (Fig. 1B). In Sa treatment, higher males than females were observed (Fig. 1A) what can explain this result. In general, the number of eggs reported here [ranging from 763.30 (± 121.88) to 1269.70 (± 202.77)] are higher than those previously recorded for C. externa reared on B. tabaci [from 293.83 (± 97.08) to 592.08 (± 62.96)] (Silva et al. 2004a) and 711.8 (± 10.57) (Auad et al. 2005) ] and A. gossypii [428.5 (± 85.2)] (Macedo et al. 2010). Similar egg survival rate was found among treatments [72.20 (± 6.68] (Fig. 1C). Higher mean values were previously observed for C. externa larvae fed with A. kuehniella and A. argillacea eggs (89.15 and 86.62 %, respectively) (Ribeiro 1988).
In general, the biological parameters recorded in this study are close to those previously reported for C. externa. The high performance of the predator when feeding on wheat aphid species confirms the importance of these species to provide food for superior populations of C. externa that have been found in fields. This situation probably occurs in other crops in which aphids develop early, and may be the basis for C. externa population growth, such as maize and cotton (Gravena 1992). This fact reinforces the need to avoid spraying insecticides or even using selective products to manage aphids. In addition, other strategies may be eventually tested in further studies in order to improve and maintain its populations in the field.
The performance obtained when Rp + Dm was provided as food shows that C. externa may develop using stink bug eggs as alternative food source, and may control itin the beginning of the pest cycle development. It was not found previous reports on predation of Pentatomidae eggs by C. externa. Although C. externa reduces its survival when it changes the diet from aphid to D. melacanthus eggs (Table 2), a greater fecundity (Fig. 1B) apparently compensates this reduction. The quick adaptation of C. externa from A. kuehniella eggs to D. melacanthus eggs suggests that insects reared in laboratory could be tested in further studies for the control of stink bug and other preys in the field. The capacity to feed on different aphid and D. melacanthus eggs is also a very favorable trait which suggests effectiveness of C. externa to control plant feeding insects in the field where population density of preys is seasonal (Carvalho & Souza 2009). Besides aphids and D. melacanthus eggs, pollen grains and honeydew may be other alternative food sources, and future studies could investigate this hypothesis.
In wheat field crops, aphids are controlled by natural occurrence of Aphidiinae (Hymenoptera: Braconidae) parasitoids. However, misguided spraying insecticides have been currently carried out to control them. Moreover, pesticides currently registered to wheat crop do not include selective insecticides to natural enemies (IAPAR 2013). Predation on Lepidoptera eggs by C. externa has also been previously reported [Alabamma argilacea (Hubner, 1818) (Lepidoptera: Noctuidae) (Carvalho et al. 1988)]. This suggest that judicious management of aphids in early stages of wheat crop are fundamental for the establishment of C. externa populations, not only for aphid control, but also for D. melacanthus control, and other Lepidoptera pests that may occasionally occur, such as Spodoptera frugiperda (J.E. Smith and Pseudaletia sequax (Franclemont, 1951) (Lepidoptera: Noctuidae). Laboratory rearing and liberation of Chrysoperla carnea (Stephens, 1836) (Neuroptera: Chrysopidae) for the control of Heliothis virescens (Fabricius, 1781) (Lepidoptera: Noctuidae) and Helicoverpa zea (Boddie, 1850) (Lepidoptera: Noctuidae) was previously successfully achieved (Macedo et al. 2010). However, providing conditions for C. externa development and conservation by feeding on aphis is probably an easier, cheaper and safer way to reduce some pests of wheat crop.
Conclusions
Results reported here evidenced that aphid preys S. avenae and R. padi present in wheat crop provided satisfactory resources for C. externa development and reproduction. D. melacanthus eggs also provided suitable performance of C. externa. Changing food (R. padi to D. melacanthus eggs) in the third instar reduced C. externa larval survival. Other parameters were not affected or were little affected by the diet change. However, when they were fed with A. kuehniella eggs in the first two instars, the food change (D. metacanthus eggs) did not affect the biological parameters. This implies that preserving C. externa populations which developed feeding on aphids may reduce not only aphids but also D. melacanthus and eventually Lepidoptera pests in wheat crop.
Literature cited
ABBAS, Q.; ARIF, M. J.; GOGI, M. D.; ABBAS, S. K.; KARAR, H. 2012. Performance of imidacloprid, thiomethoxam, acetamaprid and a biocontrol agent (Chrysoperla carnea) against whitefly, jassid and thrips on different cotton cultivars. Word Journal of Zoology 7:141-146. [ Links ]
ALBUQUERQUE, G. S.; TAUBER, C. A.; TAUBER, M. J. 1994. Chrysoperla externa (Neuroptera: Chrysopidae): life history and potential for biological control in Central and South America. Biological Control 4: 8-13. [ Links ]
AUAD, A. M.; CARVALHO, C. F.; SOUZA, B.; TREVIZANI, R.; MAGALHÃES, C. M. F. R. 2005. Desenvolvimento das fases imaturas, aspectos reprodutivos e potencial de predação de Chrysoperla externa (Hagen) alimentada com ninfas de Bemisia tabaci (Gennadius) biótipo B em tomateiro. Acta Scientiarum Agronomy 27 (2): 327-334. [ Links ]
CANTERI, M. G.; ALTHAUS, R. A.; VIRGENS FILHO, J. S.; GIGLIOTI, E. A.; GODOY, C. V. 2001. SASM – Agri: Sistema para análise e separação de médias em experimentos agrícolas pelos métodos Scott – Knott, Tukey e Duncan. Revista Brasileira de Agrocomputação 1: 18-24. [ Links ]
CARVALHO, C. F.; SOUZA, B.; SANTOS, T. M. 1998. Predation capacity and reproduction potential of Chrysoperla externa (Hagen) (Neuroptera,Chrysopidae) fed on Alabama argillacea (Hübner) eggs. Acta Zoologica Fennica 209: 83-86. [ Links ]
CARVALHO, C. F.; SOUZA, B. 2009. Métodos de criação e produção de crisopídeos. pp. 107-113. In: Bueno, V. H. P. (Ed.). Controle Biológico de Pragas. 2nd ed. Editora UFLA. Lavras. Brasil. 429 p. [ Links ]
COSTA, M. B.; BEZERRA, C. S.; SOUZA, B.; SOARES, C. S. A. 2012. Development and reproduction of Chrysoperla externa (Neuroptera: Chrysopidae) fed with Neotoxoptera formosana (Hemiptera: Aphididae). Revista Colombiana de Entomología 38 (2): 187-190. [ Links ]
COSTA, R. I. F.; ECOLE, C. C.; SOARES, J. J.; MACEDO, L. P. M. 2002. Duração e viabilidade das fases pré-imaginais de Chrysoperla externa (Hagen) alimentadas com Aphis gossypii Glover e Sitotroga cerealella (Olivier). Acta Scientiarum 24 (2): 353-357. [ Links ]
ECOLE, C. C.; SILVA, R. A.; LOUZADA, J. N. C.; MORAES, J. C.; BARBOSA, L. R.; AMBROGI, B. G. 2002. Predação de ovos, larvas e pupas do bicho-mineiro-do-cafeeiro, Leucoptera coffeellum (Guérim-Méneville & Perrollet, 1842) (Lepidoptera: Lyonetiidae) por Chrysoperla externa (Hagen, 1861) (Neuroptera: Chrysopidae). Ciência e Agrotecnologia 26 (2): 318-324. [ Links ]
FONSECA, A. R.; CARVALHO, C. F.; SOUZA, B. 2000. Resposta funcional de Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae) alimentada com Schizaphis graminum (Rondani) (Hemiptera: Aphididae). Anais da Sociedade Entomológica do Brasil 29 (2): 309-317. [ Links ]
GODOY, M. S.; CARVALHO, G. A.; MORAES, J. C.; COSME, L. V.; GOUSSAIN, M. M.; CARVALHO, C. F.; MORAIS, A. A. 2004. Seletividade de seis inseticidas utilizados em citros a pupas e adultos de Chrysoperla externa (Hagen) (Neuroptera: Chrysopidae). Neotropical Entomology 33 (3): 359-364. [ Links ]
GRAVENA, S. 1992. Controle biológico e manejo integrado. Pesquisa Agropecuária Brasileira 27: 281-299. [ Links ]
IAPAR. 2013. Reunião da Comissão Brasileira de Pesquisa de Trigo e Triticale. Informações técnicas para trigo e triticale – safra 2013/Reunião da Comissão Brasileira de Pesquisa de Trigo e Triticale, Londrina, PR, 29 de julho a 2 de agosto de 2012. – Londrina, PR : Instituto Agronômico do Paraná (IAPAR).220 p. Available In: http://www.iapar.br/arquivos/File/zip_pdf/ TrigoeTriticale2013.pdf. [Review date: 16 July 2014] [ Links ].
MACEDO, L. P. M.; PESSOA, L. G. A.; SOUZA, B.; LOUREIRO, E. S. 2010. Aspectos biológicos e comportamentais de Chrysoperla externa (Hagen, 1861) em algodoeiro. Semina: Ciências Agrárias 31 (1): 1219-1228. [ Links ]
RIBEIRO, M. J. 1988. Biologia de Chrysoperla externa (Hagen, 1861) (Neuroptera:Chrysopidae) alimentada com diferentes dietas. Dissertação (Mestrado em Fitossanidade) Escola Superior de Agricultura de Lavras, Lavras, 171 p. [ Links ]
RODRIGUES, C. A.; BATTEL, A. P. G. B.; MARTI, N. M. 2014. Dynamics and Predation Efficiency of Chrysoperla externa (Neuroptera: Chrysopidae) on Enneothrips flavens (Thysanoptera: Thripidae). Florida Entomologist 97: 653-658. [ Links ]
SILVA, J. J.; VENTURA, M. U.; SILVA, F. A.; PANIZZI, A. R. 2013. Population Dynamics of Dichelops melacanthus (Dallas) (Heteroptera: Pentatomidae) on Host Plants. Neotropical Entomology 42 (2): 141-145. [ Links ]
SILVA, C. G.; AUAD, A. M.; SOUZA, B.; CARVALHO, C. F.; BONANI, J. B. 2004A. Aspectos biológicos de Chrysoperla externa (Hagen, 1861) (Neuroptera: Chrysopidae) alimentada com Bemisia tabaci (Gennadius, 1889) biótipo B (Hemiptera: Aleyrodidae) criada em três hospedeiros. Ciência e Agrotecnologia 28 (2): 243-250. [ Links ]
SILVA, C. G.; SOUZA, B.; AUAD, A. M.; BONANI, J. P.; TORRES, L. C.; CARVALHO, C. F.; ECOLE, C. C. 2004B. Desenvolvimento das fases imaturas de Chrysoperla externa alimentadas com ninfas de Bemisia tabaci criadas em três hospedeiros. Pesquisa Agropecuária Brasileira 39 (11): 1065- 1070. [ Links ]
Received: 22-Oct-2014
Accepted: 22-Jun-2016

