











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
La polinización es de suma importancia en la agricultura, ya que es un requisito previo para el conjunto de frutas y semillas, y en la mayoría de las especies de cultivos, este servicio vital es realizado por animales, siendo los insectos los más importantes entre ellos (Khan & Yogi, 2017). Informes recientes han estimado que el 35 % del rendimiento mundial de los cultivos depende de la actividad de los polinizadores (Bailes et al., 2015; Klein et al., 2007). Es a través de las flores que las plantas han desarrollado diversas estrategias para atraer a los agentes polinizadores y asegurarse repetidas visitas que conducirán a la polinización (Simpson & Neff, 1983). La capacidad de atraer polinizadores es crucial (Bauer et al., 2017; Conner & Rush, 1996), y son varios los rasgos que influyen en la atracción, incluida la exhibición floral y el color de la flor (Bauer et al., 2017; Brunet et al. 2015; Conner & Rush, 1996; Eckhart, 1991; Mitchell et al., 2004) así como la forma, el tamaño y las recompensas florales (Whitney & Glover, 2007). Esto ha llevado a sugerir que, la selección de rasgos florales en los cultivos podría conducir a mejoras al atraer un mayor número de polinizadores (Bailes et al., 2018) y, por lo tanto, un mejor rendimiento de los cultivos (Bailes et al., 2015). El trabajo en torno a las interacciones planta-polinizador puede informarnos sobre las mejores estrategias para lograrlo (Bailes et al., 2015).
Por otra parte, la diversidad genética natural presente en forma de parientes silvestres, así como de variedades locales de cultivos, son recursos con una amplia base de rasgos valiosos que se pueden aprovechar para los programas de mejora de cultivos (Saini et al., 2020). Para ello es necesario conocer características del germoplasma existente, tanto in situ, como ex situ (Rodríguez et al., 2009). Sin embargo, existe una brecha importante entre materiales conservados, y aquellos caracterizados y evaluados (Rodríguez et al., 2009). Una manera de llevar a cabo la conservación de esta diversidad es a través de bancos de germoplasma que mantienen colecciones vegetales de especies cultivadas y parientes silvestres, estas colecciones reciben el nombre de accesiones y de acuerdo con la Organización de las Naciones Unidas para la Alimentación y la Agricultura [FAO] (2022), el término accesión hace referencia a una muestra distinta, identificable de semillas que representa un cultivar, una línea de cría o una población y que se mantiene almacenada para su conservación y uso.
Ejemplos de esta diversidad genética se encuentra en los sistemas de cultivo tradicionales de México, como el agroecosistema milpa (Montes-Hernández et al., 2005), caso concreto en Yucatán, que consiste en la siembra asociada de maíz Zea mays (Poaceae), frijol lima Phaseolus lunatus, frijol xcoolibuul Phaseolus vulgaris (Fabaceae) y calabaza Cucurbita spp. (Cucurbitaceae), junto con otros cultivos hortícolas (Canul et al., 2005; Rodríguez et al., 2009). Es importante destacar que el género Cucurbita es uno de los más importantes, y cuenta con 27 especies (Hernández, 1978; Whitaker, 1968). Las especies de este género forman el grupo conocido como calabazas, de las cuales cinco han sido domesticadas: Cucurbita pepo (calabaza de india), C. ficifolia (chilacayote), C. moschata (calabaza de castilla), C. maxima (calabaza kabocha) y C. argyrosperma (calabaza pipiana) que son importantes desde el punto de vista económico, nutricional y cultural, tanto a nivel nacional como mundial. Las partes alimenticias van desde los frutos inmaduros y maduros, semillas, flores y algunas partes vegetativas (Lira, 1995; Villanueva, 2007). Es conocido que la diversidad fenotípica dentro de las poblaciones locales de Cucurbita spp. es alta (Garzón et al., 1993; Lira, 1995; Montes-Hernández et al., 2005; Whitaker & Davis, 1962).
La mayoría de oraciones citadas en éste párrafo no están relacionadas en las referencias por favor proveer la referencia completa o casi que se eliminaría el párrafo.
Existen antecedentes de trabajos realizados con polinizadores y visitantes florales en Cucurbitaceae, tales como los estudios de McGrady et al. (2019) en donde las tasas de visitas de abejas silvestres en agroecosistemas comerciales de Cucurbita influyeron positivamente en las variables de rendimiento, así como también una mayor densidad de flores se relaciona con un mayor número de visitas. Por otra parte, Mazzei et al. (2020) identificaron visitantes polinizadores y no polinizadores en flores del género Cucurbita y como la presencia de éstos últimos influía en las visitas de polinizadores, siendo positiva negativa o neutra dependiendo de la familia. También, Delgado-Carrillo et al. (2018) efectuaron estudios sobre los servicios de polinización en C. moschata y obtuvieron datos de polinizadores asociados al cultivo, con respecto a la variación temporal e identificaron que la predominancia de ciertos polinizadores cambiaba de acuerdo con la variación estacional. Piedade et al. (2016) realizaron estudios en los atributos florales de híbridos de melón y determinaron cuáles influían en la atracción para polinizadores; encontraron diferencias entre los híbridos en cuanto a los atributos, en el sexo de la flor y en la frecuencia de visitas.
Si se considera la importancia de los tipos de visitantes en los cultivos y en cómo los rasgos florales pueden determinar la capacidad de atracción y la diversidad existente entre el género Cucurbita, es necesario ampliar la información de lo que ocurre en las variedades o accesiones locales de cada región, en este caso para la península de Yucatán. Por tanto, el presente trabajo tiene como objetivo evaluar las características florales en accesiones de calabaza pertenecientes a la península de Yucatán relacionadas con la atracción de insectos visitantes.
Materiales y métodos
Material vegetal y condiciones de cultivo.
El trabajo experimental se desarrolló en un huerto familiar ubicado en el municipio de Baca, Yucatán México (21°06′21″N, 89°24′11″O, 14 m s.n.m.). Se utilizaron semillas de cinco accesiones de calabaza Cucurbita argyrosperma Huber (Cucurbitaceae) procedentes de productores en municipios del estado de Yucatán y Campeche, las cuales son conservadas en banco de germoplasma en el Instituto Tecnológico de Conkal, Yucatán. Las accesiones RRS004 y RRS006 de Mérida (Yucatán), RRS005 procedente de Peto (Yucatán) y RRS008 al igual que la RRS010 con origen en Hopelchén (Campeche) y Quetzal (Campeche), respectivamente. Se sembraron tres semillas de cada accesión por poceta a una distancia de 0,60 m entre plantas y 1,80 m entre líneas, en un diseño de bloques al azar con 60 plantas por línea, con un total de 5 líneas; para el riego se utilizó un sistema por goteo. Las pocetas antes de la siembra fueron abonadas con bagazo de henequén, y las actividades restantes de manejo se realizaron de acuerdo con las prácticas locales.
Evaluación de rasgos florales.
En este estudio se incluyeron aquellos caracteres florales involucrados en la atracción de polinizadores (Bailes et al., 2018; Bauer et al., 2017; Unni et al., 2021), las mediciones se realizaron solo en las flores estaminadas, debido a las flores pistiladas se presentaron en cantidades bajas. Las variables evaluadas para rasgos florales fueron las siguientes: diámetro de corola, altura de la flor, área floral, producción total de polen, viabilidad de polen, producción néctar, contenido de azúcar en néctar. Por otra parte, para los visitantes las variables evaluadas fueron: el número de visitantes, el tipo de visita y la identificación del insecto visitante hasta nivel género. Todas las variables medidas se realizaron en las mismas plantas en diferentes flores (por la vida corta de la flor) y durante el período de floración máxima (30 días después de la siembra)͙.
Se realizaron muestreos en campo para evaluar rasgos de tamaño (Fig. 1). Se tomaron fotografías de una flor por planta en las vistas lateral y cenital con una cámara digital (Nikon D5100) y se utilizaron las imágenes laterales para medir la altura de la flor y las cenitales para el diámetro de la corola. Adicionalmente se midió el área floral de igual manera con fotografías de los pétalos, cada imagen incluía una escala y se utilizó el software ImageJ, con la herramienta seleccionar área y analizar (versión 1.52a, National Institute of Health, EE. UU.), así como también la exhibición floral que es igual al número de flores abiertas en una planta (Bauer et al., 2017) en 15 plantas por accesión (Ferreira & Torezan, 2013).
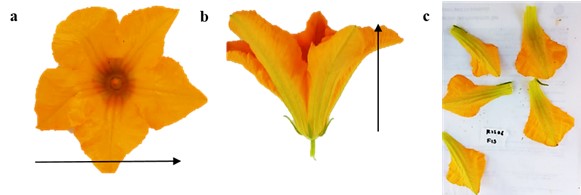
Figura 1 Vista cenital (a) y lateral (b) de flor estaminada de Cucurbita sp.; medición del área en flor estaminada de Cucurbita sp. (c).
Para determinar la producción total de polen, se recogieron flores estaminadas un día antes de la antesis. Después de la recolecta, la corola se retiró con una cuchilla y la base de la flor con anteras se colocó en viales individuales de 12 mL (Stanghellini et al., 2002). Cada vial contenía 10 mL de suspensión de agua con una gota de detergente, luego la antera se presionó con el apoyo de una aguja para liberar todos los granos de polen y se eliminaron los residuos visibles con un par de pinzas finas. Mientras se agitaba suavemente la suspensión, utilizando una micropipeta se tomaron 0,01 mL para colocarlos en un portaobjetos marcado con líneas. Suavemente se colocó un cubreobjetos de tal manera que el líquido no fluyera ni se esparciera fuera, el portaobjetos se llevó bajo un microscopio (Digital Usb 1000 X, GLOptics) para el conteo (Shivanna & Tandon, 2014). Para estimar la cantidad total de granos de polen se utilizó la siguiente fórmula:
Para la viabilidad de polen se utilizó el método de tinción con cloruro de 2, 3, 5-trifenil tetrazolio (CTT) al 1 % añadiendo 0,2 g de CTT y 12 g de sacarosa, disueltos en 20 mL de agua destilada. Se dejaron caer dos gotas de la mezcla en un portaobjetos y se espolvoreó el polen para luego colocar un cubreobjetos. Se llevaron las muestras a microscopio para realizar los recuentos de viabilidad del polen después de 30-40 min de incubación en un lugar oscuro. Los granos de polen teñidos de rojo brillante se contaron como viables. La viabilidad se expresó en porcentaje (%) utilizando la siguiente fórmula (Vijayakumar et al., 2018):
Para estimar el volumen y el contenido de azúcares totales en el néctar se protegieron las flores estaminadas una noche antes de la antesis para evitar que los visitantes tomen el néctar antes de la estimación. Poco después de la antesis, se retiraron las bolsas de las flores seleccionadas para recolectar néctar. Se recolectó el néctar insertando suavemente un tubo capilar de 15 μL en el nectario durante tiempo suficiente para el movimiento del néctar en el tubo capilar. La cantidad de néctar se estimó midiendo la longitud del tubo lleno de néctar y se calculó su proporción sobre la base de la longitud total del tubo capilar. Este procedimiento se realizó en 15 plantas por accesión con una flor por planta, con un total de 15 flores por accesión. Luego se dispensó el néctar del tubo capilar sobre la superficie del refractómetro portátil calibrado y se realizó la lectura que indica la concentración de azúcares en porcentaje (Shivanna & Tandon, 2014).
Evaluación de visitantes florales.
Para las visitas florales, un observador pasó tres minutos por flor cada hora en 15 flores estaminadas en un horario de 07:00 am a 10:00 am (No se incluyeron las flores pistiladas debido a la baja cantidad de producción). Se contó el número de visitantes. Se utilizaron succionadores bucales y frascos con etanol al 70 % para captura, colecta y posterior identificación (Enríquez et al., 2015). El observador mantuvo distancia con respecto a los visitantes florales en las flores y solo había acercamiento en un corto tiempo para la colecta, para inmediatamente retirarse. También se registró el tipo de visita (legítima o ilegítima). Una visita legítima se definió como cualquier contacto físico entre el cuerpo de un visitante y la antera, mientras que una visita ilegítima se consideraba como cualquier otro evento sin contacto físico con las anteras (Gélvez et al., 2018). Para la identificación de los insectos se realizó hasta nivel de género con las guías de Krysan (1986), Mackay y Mackay (1989), Ayala (1999), Brothers y Finnamore (1993), Michener et al. (1994), y Soto y Retana (2003).
Análisis de datos.
Las variables de morfología floral como diámetro de la corola, altura y área floral se analizaron con un análisis de varianza (ANOVA, p ≤ 0,05) para cada variable (sólo para el área floral los datos fueron transformados con logaritmo natural) para posteriormente realizar comparación de medias con la prueba de Tukey (p ≤ 0,05). Estos análisis se realizaron con el paquete estadístico InfoStat. Para las variables de recompensas florales y exhibición floral los datos fueron transformados: exhibición floral y producción total de polen a raíz cuadrada, viabilidad de polen y contenido de azúcar en néctar con el arcoseno de la raíz cuadrada y producción de néctar con logaritmo natural. Posteriormente, se analizaron con análisis de varianza (ANOVA, p ≤ 0,05) para cada variable y luego se aplicó una prueba de Tukey (p ≤ 0,05). Estos análisis se realizaron con el paquete estadístico InfoStat. Los datos obtenidos de visitantes florales, tales como la riqueza y abundancia de géneros entre accesiones, riqueza y abundancia de visitantes legítimos e ilegítimos entre accesiones, así como proporción de individuos por género entre accesiones se analizaron con modelos lineales mixtos generalizados (MLMG), en los cuales los efectos fijos fueron las accesiones y los aleatorios fueron las plantas. Para todas las variables de respuesta, se utilizó la distribución de Poisson y se compararon las medias con el test de Bonferroni secuencial utilizando el software IBM SPSS Statistics.
Se realizó un análisis de componentes principales para evaluar la proximidad entre las accesiones respecto a los rasgos florales y las variables morfológicas. Este análisis se realizó en el programa Infostat. Asimismo, se realizó un análisis de correspondencia canónica y un análisis de redundancia para evaluar asociaciones entre los rasgos florales con los visitantes florales totales y con los visitantes legítimos, respectivamente. El criterio de selección de los análisis se basó en la longitud de gradiente del eje 1. La longitud de gradiente se calculó mediante un análisis de correspondencia distendido. La significancia de cada variable explicativa (rasgos florales), así como del primer y los cuatro ejes, se examinaron a través de la prueba de permutaciones aleatorias de Monte Carlo (499 permutaciones, P < 0,05). Estos análisis se desarrollaron usando el programa Canoco 4.5.
Resultados
Morfología y exhibición floral.
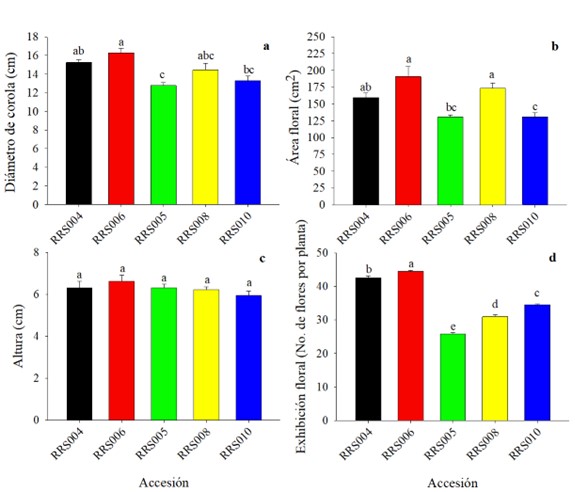
Se encontraron diferencias estadísticamente significativas (p ≤ 0,05) entre las accesiones de Cucurbita argyrosperma (Fig. 2) para las variables diámetro, área y exhibición floral. En la altura de flor no se registraron diferencias significativas (F = 1,07, gl = 4, p = 0,38) (Fig. 2c). La accesión RRS006 es la que presentó el mayor diámetro con 16,28 ± 0,4925 cm y RRS005 el menor con 12,77 ± 0,3413 cm (F = 8,08, gl = 4, p = 0,0001) (Fig. 2a). En área floral las accesiones RRS006 y RRS008 también presentaron diferencias estadísticamente significativas (F = 10,.43, gl = 4, p = 0,0001) y los valores más altos con 191 ± 14,66 cm2 y 173,19 ± 7,71 cm2, respectivamente. Por el contrario, RRS005 fue la que presentó menor área floral con 130,72 ± 2,98 cm2 (Fig. 2b). Para el número total de flores producidas por planta el mayor número se registró en la accesión RRS006 con 44,47 ± 0,2363 flores y el menor en RRS005 con 25,87 ± 0,3217 (F=2.43, gl = 4, p = 0,029) (Fig. 2d).
Recompensas florales.
Producción y viabilidad de polen en las flores. Para la producción total de polen entre las accesiones de C. argyrosperma. se registraron diferencias estadísticamente significativas (F = 33,50, gl = 4, p= 0,0001) (Fig. 3a). Los valores oscilaron entre 381.264 y 182.592 granos de polen por flor, la mayor producción de polen se obtuvo en la accesión RRS006. Para la viabilidad del polen también se presentaron diferencias significativas (p ≤ 0,05) entre las accesiones (F = 8,71, gl = 4, p = 0,0001) (Fig. 3b). Los valores más altos se registraron en RRS004 (46 %) y RRS005 (43 %) seguido de RRS008 (36 %), el valor más bajo se registró en la accesión RRS006 con menos del 30 %
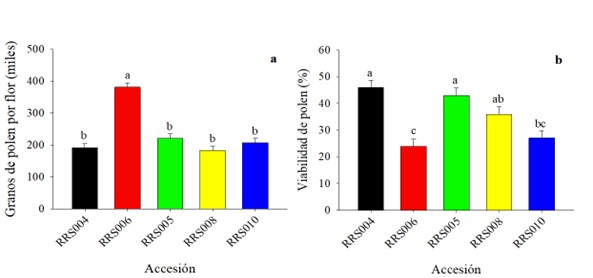
Figura 3 Producción y viabilidad de polen en flores estaminadas de accesiones de calabaza (Cucurbita argyrosperma). Los datos son medias (± error estándar), los valores con literal diferente son estadísticamente diferentes (Tukey, p ≤ 0,05).
Producción total de néctar acumulado. La producción de néctar presentó diferencias estadísticamente significativas entre las accesiones (F = 7,91, gl = 4, p = 0,0001). Los valores oscilaron entre 64,31 ± 1,92 µL y 25,39 ± 1,08 µL (Fig. 4a). En la producción de néctar por hora se registraron valores bajos al inicio y al final (8,77 ± 0,8212 µL, 8,06 ± 0,8200 µL, respectivamente) y valores altos (19,88 ± 0,8189 µL, 15,63 ± 0,8192 µL) en las horas intermedias (Fig. 4b).
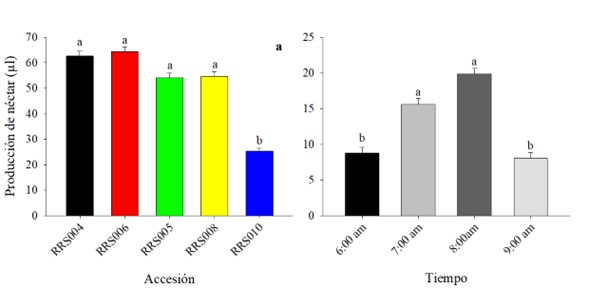
Figura 4 Producción total de néctar acumulado de flores estaminadas en accesiones de calabaza (Cucurbita argyrosperma). Los datos son medias (± error estándar), los valores con literal diferente son estadísticamente diferentes (Tukey, p ≤ 0,05).
Contenido de azúcares en néctar. También se presentaron diferencias estadísticamente significativas (p ≤ 0,05) para el contenido de azúcares entre las accesiones (F = 4,44, gl = 4, p = 0,0030). Los valores oscilaron entre 34,63 ± 1,06 °Brix y 28,97 ± 2,05 °Brix (Fig. 5a). En el contenido de azúcares por hora se registraron los valores más altos en las primeras horas de la mañana, mismos que fueron disminuyendo conforme avanzaron las horas del día (Fig. 5b).
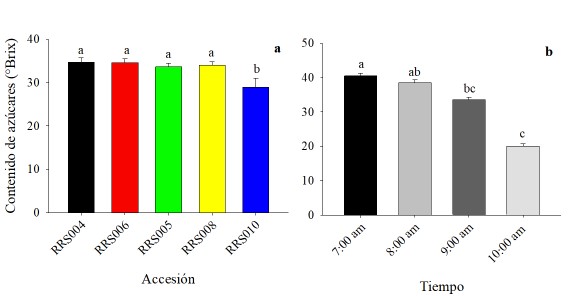
Figura 5 a) Contenido de azúcares en flores estaminadas de accesiones de calabaza (Cucurbita argyrosperma). b) Contenido de azúcares en néctar por hora, de las 5 accesiones de calabaza. Los datos son medias (± error estándar), los valores con literal diferente son estadísticamente diferentes (Tukey, p ≤ 0.05).
Visitantes florales. Se registró un total de 4.465 individuos en todas las accesiones pertenecientes a 7 familias y 12 géneros. El género más abundante fue Frankliniella, por el contrario, dos géneros, Plebeia y Augochlora estuvieron representados por menos de 10 individuos (Tabla 1).
Para la riqueza y abundancia de visitantes florales entre accesiones, se presentaron diferencias estadísticamente significativas (p ≤ 0,05). El número medio de géneros en la accesión RRS010 fue significativamente más alto en comparación con las demás accesiones (Fig. 6a). Y para la abundancia, la accesión RRS008 fue la que presentó el valor más alto, seguido de la RRS006 con un valor por encima de los 40 individuos (Fig. 6b).
Tabla 1 Número total de individuos por género para cada accesión de Cucurbita argyrosperma
| Orden | Familia | Género | RRS004 | RRS006 | RRS005 | RRS008 | RRS010 | Total |
|---|---|---|---|---|---|---|---|---|
| Coleoptera | Chrysomelidae | Acalymma | 6 | 24 | 3 | 7 | 8 | 48 |
| Diabrotica | 8 | 11 | 3 | 8 | 10 | 40 | ||
| Diptera | Drosophilidae | Drosophila | 22 | 15 | 3 | 26 | 4 | 70 |
| Hymenoptera | Halictidae | *Augochlora | - | - | 2 | 1 | 4 | 7 |
| Apidae | *Ceratina | 20 | 8 | 18 | 11 | 12 | 69 | |
| *Partamona | 16 | 6 | 27 | 17 | 43 | 109 | ||
| *Peponapis | 8 | 5 | 6 | 3 | 2 | 24 | ||
| *Plebeia | - | - | 3 | - | 7 | 10 | ||
| Vespidae | Polybia | 72 | 37 | 78 | 11 | 71 | 271 | |
| Formicidae | Neivamyrmex | - | 98 | - | - | - | 98 | |
| Solenopsis | 6 | - | 53 | 190 | 160 | 409 | ||
| Thysanoptera | Thripidae | Frankliniella | 344 | 1084 | 423 | 1220 | 239 | 3310 |
* Visitas legítimas
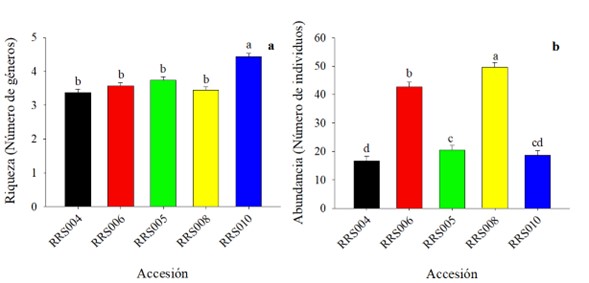
Figura 6 Riqueza y abundancia de insectos visitantes en flores estaminadas de accesiones de calabaza (Cucurbita argyrosperma). Los datos son medias, los valores con literal diferente son estadísticamente diferentes (Bonferoni secuencial).
Con respecto a la proporción de individuos por género, Acalymma registró el número más alto de individuos en la accesión RRS006 (Fig. 7a), y Augochlora en RRS010 (Fig. 7b). En el género Frankliniella son las accesiones RRS006 y RRS008 que de igual manera presentaron los valores más altos (Fig. 7f) y en Drosophila la RRS004, RRS006 y RRS008 (Fig. 7e). Por el contrario, en Partamona (Fig. 7h) y Polybia (Fig. 6k) en las accesiones RS006 y RRS008, respectivamente, se encontraron los números más bajos. Por su parte en los géneros Ceratina, Diabrotica, Peponapis y Plebeia no hubo diferencias significativas entre accesiones (Figs. 7c-7d, 7i-7j). Neivamyrmex (Fig. 7g) se observó solamente en la accesión RRS006 y la mayor proporción de Solenopsis en RRS008 y RRS010 (Fig. 7l).

Figura 7 Proporción de individuos por género en flores estaminadas de accesiones de calabaza (Cucurbita argyrosperma). Los valores con literal diferente son estadísticamente diferentes (Bonferroni secuencial).
La riqueza por tipo de visita presentó diferencias significativas en las visitas legítimas (Fig. 8a) y es la accesión RRS004 la que registró los valores más altos, por el contrario, en las ilegítimas no hubo diferencias (Fig. 8c). Así también, con respecto a la abundancia para ambos tipos se encontraron diferencias, en donde la accesión RRS004 obtuvo el valor más alto para las legítimas (Fig. 8b), y RRS006 y RRS008 para las ilegítimas (Fig. 8d).
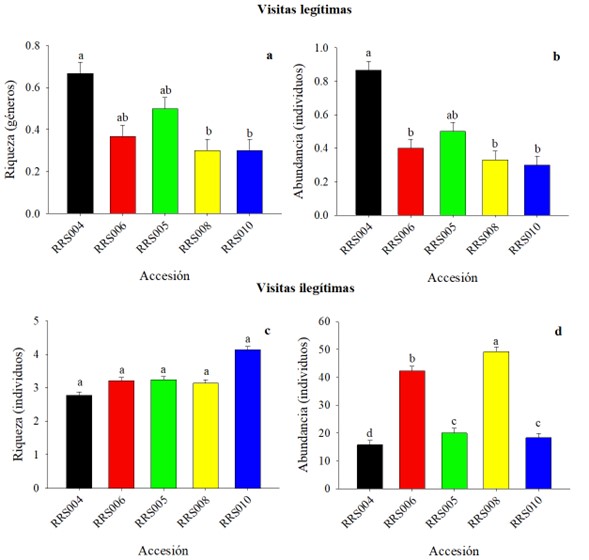
Figura 8 Riqueza y abundancia de insectos visitantes de flores estaminadas en las accesiones de calabaza (Cucurbita argyrosperma) clasificados por tipo de visita. Los datos son medias (± error estándar), los valores con literal diferente son estadísticamente diferentes (Bonferroni secuencial).
Asociación entre rasgos florales y visitantes florales. El análisis de componentes principales se realizó usando siete variables de rasgos florales. Cuatro componentes fueron seleccionados como los factores más significativos con valores propios > 1. Esos componentes explicaron el 74 % de la variación. El primer componente (CP 1) explicó el 22 % de la variación total de los datos originales, el segundo componente (CP 2) explicó el 21 %, y el tercer y cuarto componente (CP 3 y CP 4) explicaron cada uno, el 15 %. Los porcentajes de varianza explicada por los cuatro componentes y su correlación con las variables originales de los rasgos florales son mostrados en la Tabla 2.
Tabla 2 Varianza explicada por cuatro componentes principales derivados de siete variables de rasgos florales de Cucurbita argyrosperma.
| Ejes | CP1 | CP2 | CP3 | CP4 |
|---|---|---|---|---|
| Eigenvalores | 1,51 | 1,44 | 1,07 | 1,04 |
| Variación (%) | 0,22 | 0,21 | 0,15 | 0,15 |
| Var. Acumulada (%) | 0,22 | 0,42 | 0,57 | 0,72 |
| Correlación con las variables originales | ||||
| Ancho corola | 0,43 | 0,47 | 0,36 | -0,21 |
| Altura flor | 0,03 | 0,45 | 0,70 | -0,21 |
| Área floral | 0,73 | 0,16 | -0,28 | -0,21 |
| Producción polen | 0,67 | -0,46 | 0,05 | 0,14 |
| Viabilidad polen | -0,42 | 0,62 | -0,37 | -0,03 |
| Producción néctar | 0,40 | 0,60 | -0,37 | 0,30 |
| Contenido azúcar | -0,50 | 0,13 | 0,31 | 0,91 |
El CP 1 tuvo una contribución mayoritaria por el área floral y producción de polen, mientras que el CP 2 fue formado principalmente por la viabilidad del polen y la producción de néctar. En este mismo sentido, el CP 3 tuvo una casi exclusiva contribución por la altura de la flor y el CP 4, fue formado fuertemente por el contenido de azúcar (Tabla 2). Por otro lado, es interesante mencionar que la accesión RRS006 fue la única que tuvo una clara proximidad a las variables de área floral y producción de polen, el resto de las accesiones no mostró un patrón tan definido (Fig. 9).
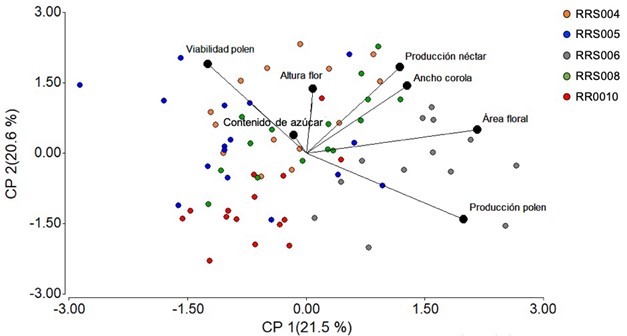
Figura 9 Análisis de Componentes Principales mostrando la proximidad de las accesiones de Cucurbita argyrosperma con rasgos florales.
Cuando se analizó una posible asociación de los visitantes florales hacia los rasgos florales, se encontró una reducida separación de los visitantes florales sobre los ejes, aunque el eje 1 y los cuatro ejes mostraron una ordenación de los visitantes florales de parcialmente significativo a significativo (Tabla 3).
Tabla 3 Valores propios y resultados de los análisis de Monte Carlo para un Análisis de Correspondencia Canónica (CCA) de los visitantes florales totales (VFT) y Análisis de Redundancia (RDA) de los visitantes florales legítimos (VFL), asociados a los rasgos florales de cinco accesiones de Cucurbita argyrosperma.
| Visitantes florales totales (VFT) | Ejes | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| Valores propios: | 0,124 | 0,071 | 0,038 | 0,018 |
| Correlaciones de los rasgos florales con los VFT: | 0,506 | 0,486 | 0,453 | 0,413 |
| Varianza acumulada de los VFT (%): | 6,5 | 10,2 | 12,1 | 13,1 |
| Varianza acumulada de los rasgos florales -VFT: | 47,2 | 74,0 | 88,5 | 95,3 |
| Significancia del primer eje canónico: | F = 4,63 | p = 0,09 | ||
| Significancia de todos los ejes canónicos: | F = 1,52 | p = 0,04 | ||
| Visitantes florales legítimos (VFL) | ||||
| Valores propios: | 0,174 | 0,025 | 0,012 | 0,002 |
| Correlaciones de los rasgos florales con los VFL: | 0,580 | 0,296 | 0,345 | 0,189 |
| Varianza acumulada de los VFL (%): | 17,4 | 19,9 | 21,1 | 21,3 |
| Varianza acumulada de los rasgos florales -VFL: | 81,6 | 93,2 | 98,8 | 99,8 |
| Significancia del primer eje canónico: | F = 14,107 | p = 0,002 | ||
| Significancia de todos los ejes canónicos: | F = 2,592 | p = 0,002 | ||
Además, se encontró que los rasgos florales tales como la producción de polen (F= 2,88, p= 0,008) y néctar (F = 2,37, p = 0,03), y el área floral (F = 2,27, p = 0,05) tuvieron efectos importantes en la asociación con los visitantes florales. En este sentido, el género Neivamyrmex estuvo asociado a flores con mayor área floral y producción de polen, en tanto que el género Solenopsis estuvo asociado a flores con menor producción de néctar (Fig. 10a).
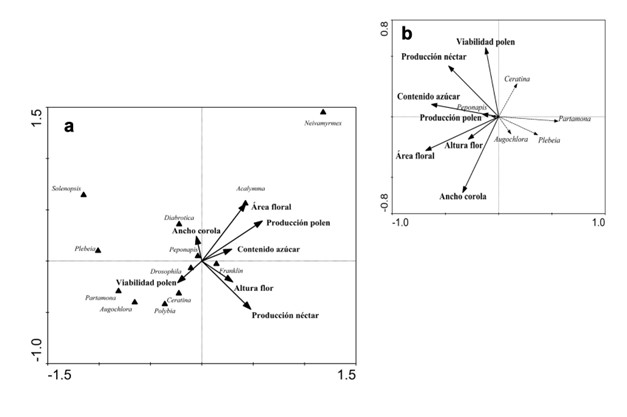
Figura 10 Asociación de los visitantes florales totales (a) y legítimos (b) hacia los rasgos morfológicos florales en cinco accesiones criollas de Cucurbita argyrosperma.
Por otra parte, cuando se analizó la asociación de los rasgos florales con los visitantes florales legítimos, se encontró que la ordenación de los géneros a pesar de ser reducida, si fue significativa para todos los ejes (Tabla 3).
Los rasgos florales como área floral (F = 6,72, p = 0,004), contenido de azúcar (F = 5,85, p = 0,004), producción de néctar (F = 3,35, p = 0,03) y ancho de corola (F = 2,26, p = 0,07), tuvieron efectos mayormente significativos en la asociación con los géneros de visitantes florales. Por ejemplo, se registró que el género Partamona estuvo asociado negativamente con el área floral, contenido de azúcar y producción de néctar; mientras que el género Ceratina tuvo también una asociación negativa, pero con el ancho de la corola (Fig. 10b).
Discusión
En este estudio se ha demostrado que existe una variación considerable en los rasgos florales de las accesiones de calabaza y que influyen en la atracción de diferentes visitantes florales. Los resultados obtenidos proporcionan evidencia de esta relación en la cual un mayor número de visitantes puede implicar la predominancia de visitas legítimas o bien de ilegítimas, lo cual dependerá de la afinidad de los visitantes hacia la accesión evaluada.
Se encontró que el tamaño de la corola tuvo asociación con los visitantes florales tanto positiva como negativa. En el caso de la asociación positiva, los resultados sugieren que esta característica hace más fácil la ubicación de las flores desde lejos y se traduzca como un indicador de la gran disponibilidad de recursos (Brunet et al., 2015). Bauer et al. (2017) encontraron que mayores tamaños de flores atrajeron más abejas, en nuestro estudio encontramos esta asociación positiva con el género Neyvamirmex.
En cuanto a la asociación negativa en Ceratina y Partamona, Mallinger y Prasifka (2017) encontraron resultados similares debido a que registraron que las tasas de visitas disminuían con la longitud de la corola. Estas variaciones pueden deberse a la especificidad de las especies involucradas ya que familias de abejas silvestres y grupos de abejas funcionalmente distintos tienen respuestas únicas a la expresión de rasgos florales (Rowe et al., 2020).
Consideramos que este trabajo aporta información complementaria ya que incluye visitantes florales legítimos como ilegítimos y que, en ambos grupos de insectos, los mismos rasgos atraen a los dos tipos de visitantes con variaciones específicas a nivel grupo o género.
Por otro lado, llama la atención que cuando evaluamos la altura de los tubos de las corolas, estas no difirieron en profundidad. No obstante, en otros estudios como el de Ricou et al. (2014) se han encontrado relaciones entre la morfología de las corolas y la de los insectos visitantes específicos para optimizar su forrajeo. Newman y Jhonson (2021) también encontraron que las longitudes de los tubos florales correspondieron a la longitud de la probóscide de sus visitantes (en este caso moscas).
La producción y viabilidad de polen tuvo diferencias entre accesiones y si presentó asociación con los visitantes florales. Tal como informaron Ricou et al. (2014) el polen es un recurso importante, que influye en las decisiones de forrajeo de abejorros, pero también en visitantes ilegítimos como los crisomélidos (Eben, 2012).
Ahora bien, para complementar esta información se sugiere realizar estudios de composición. Vaudo et al. (2020) analizaron proporciones de proteína-lípidos en el polen que dan forma a las preferencias de alimentación. Los resultados de Ghosh et al. (2020) de igual manera indicaron que el primer criterio para la preferencia de alimentación de las abejas sería el contenido nutricional de proteínas, pero además la disponibilidad de recursos de las fuentes florales.
En la producción de néctar y contenido de azúcares no se encontraron diferencias entre accesiones, sin embargo también este rasgo floral tuvo asociación con los visitantes florales, esto puede deberse a que el néctar proporciona la principal fuente de energía, y su calidad tiene un papel crítico en las especies que forrajean sobre las flores, tomando en este sentido gran importancia el contenido balanceado de los azúcares basados en sacarosa, glucosa y fructosa. Por ejemplo, la disimilaridad de las preferencias florales de las abejas melíferas, atraídas por un contenido balanceado de los tipos de azúcares; y los abejorros, atraídos por la mezcla (no necesariamente balanceada) de los azúcares (Ricou et al., 2014). Además, habría que considerar la influencia de los compuestos volátiles (Gaffney et al., 2020; Rering et al., 2018) y el contenido de aminoácidos del néctar (Barberis et al., 2021) ya que podrían estar interviniendo en estas respuestas.
Para los géneros registrados en este estudio se encontraron similitudes con otras investigaciones en Cucurbita (aunque en diferentes abundancias). Bazo et al. (2018) registraron en C. maxima familias como Drosophilidae, Chrysomelidae, Apidae y Vespidae. En Yucatán, Meléndez-Ramírez et al. (2002) reportaron de 8 a 12 géneros en cultivos de Cucurbita incluyendo solo polinizadores, pero sólo cuatro géneros como más abundantes, Augochlora, Ceratina, Partamona, y Peponapis, registrados también para nuestro estudio.
En cuanto al género Peponapis es considerado especializado al género Cucurbita con el que comparte una historia evolutiva y el cual ha resultado en una alta eficiencia de polinización (Delgado-Carrillo et al., 2017).
Delgado-Carrillo et al. (2018) mencionaron otro grupo de polinizadores considerados potenciales para Cucurbita, las abejas halíctidas nativas, abejas Ceratina y abejas sin aguijón, a las cuales pertenecen las reportadas para este estudio. El mismo autor compara la eficiencia en polinización con respecto a Peponapis con la observación de ser menos abundantes, pero que implica una complementariedad en los procesos de polinización que es reflejada en los rendimientos.
En la proporción de visitantes a nivel género se encontró alta variación en la predominancia entre las accesiones. Es posible que este resultado sea debido a que cada género presenta particularidades en sus elecciones en sinergia con los rasgos florales. Sin embargo, en los géneros en los cuales no se hallaron diferencias entre accesiones existe la probabilidad de que el motivo sea que los visitantes se enfrentan a las señales en conjunto de toda la flor y esto puede repercutir para que en sus elecciones no discriminen la calidad de la recompensa (Carr et al., 2015).
También, podrían estar involucradas las interacciones de los rasgos florales con otros factores como las feromonas para la atracción de pareja y esto podría explicar las grandes agregaciones de individuos en Frankliniella (Akinyemi, 2021). De igual manera la variación en la presencia de los géneros de hormigas en las accesiones puede estar relacionado con la composición del néctar (tipo de azúcares y aminoácidos) que además interactúa con los requerimientos fisiológicos óptimos de la especie y de sus relaciones competitivas dentro de la comunidad (Blüthgen & Fiedler, 2004).
Por otra parte, al igual que en el trabajo de Gélvez et al. (2018), la mayoría de las visitas en las flores fueron predominantemente ilegítimas. A pesar de esto, es posible que las actividades de los visitantes ilegítimos no tengan efectos directos en la producción de semillas (Irwin et al., 2015), o bien puedan afectar o no a los visitantes legítimos. Mazzei et al. (2020) encontraron que la presencia de drosophílidos resultaba neutral para los visitantes legítimos, contrario a la presencia de hormigas y coleópteros en el mismo estudio. Por tal motivo, se sugieren realizar otros estudios que involucren el rendimiento y producción de semillas, así como identificar el recurso utilizado en las visitas ilegítimas.
A pesar de que la calabaza es un cultivo de gran importancia, son pocos los estudios que complementan la información de rasgos y visitantes. De manera general sucede con una gran parte de las especies de plantas, algunos trabajos se enfocan solo en características florales (Bertazzini & Forlani, 2016) o solo en diversidad de abejas (Meléndez-Ramírez et al., 2002). Por tal motivo la riqueza y abundancia de visitantes florales en el género Cucurbita como tal no se puede comparar fácilmente con otros estudios debido al enfoque y los objetivos de los mismos. A pesar de esto, en este estudio se demostró que los rasgos florales si influyen en la atracción de visitantes florales, y que esto puede repercutir en los procesos de polinización sumamente importantes para la agricultura.
Conclusiones
Encontramos variación de los rasgos florales entre las accesiones de calabaza, y estos, sí tuvieron asociación con la presencia de insectos visitantes. Las características del tamaño floral, así como el polen presentaron un papel determinante en la atracción de visitantes legítimos e ilegítimos y estos variaron entre las accesiones. Con respecto al néctar, este de igual manera tuvo asociación con los visitantes. Además, en todas las accesiones evaluadas se presentó una mayor cantidad de visitas ilegítimas que visitas legítimas y entre los visitantes ilegítimos se identificaron géneros de crisomélidos, drosofílidos, trips, hormigas y avispas; para los visitantes legítimos se encontró la presencia de abejas de los géneros Augochlora, Ceratina, Peponapis, Partamona y Plebeia.
En cuanto a la relación de los rasgos con los visitantes a nivel género se encontró variabilidad en los resultados, que va a depender del género en estudio y de sus necesidades específicas.
También es importante destacar que los resultados obtenidos en este trabajo enriquecen los recursos de información existentes y se sugiere realizar estudios que incluyan más variables tales como aroma y color floral, para usar esta información en la selección de accesiones con atributos de interés. Dichos estudios se necesitan a nivel local ya que cada región contiene un ambiente particular con necesidades diferentes.

