










Services on Demand
Journal
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690
Rev Colom Cienc Pecua vol.28 no.2 Medellín Apr./June 2015
https://doi.org/10.17533/udea.rccp.v28n2a04
ORIGINAL ARTICLE
doi: 10.17533/udea.rccp.v28n2a04
Association between BMP4 gene polymorphism and in vitro embryo production traits in Gyr cows¤
Asociación entre polimorfismo en el gen BMP4 y características de producción in vitro de embriones en vacas Gyr
Associação entre o polimorfismo no gene BMP4 e características de produção in vitro de embriões em vacas Gyr
Wilder H Ortiz11*,DVM, MSc, Dr. AnSc(c); Celia R Quirino1, Ag Eng PhD; Amanda Silva1, Biol, MSc; Clara S Oliveira2, DVM, PhD; Raquel V Serapião3, DVM, PhD; Aline Pacheco1, DVM, PhD; Aylton Bartholazzi1, DMV, AnSc.
1Laboratório de reprodução e melhoramento genético animal, Centro de Ciência e Tecnologia, Universidade Estadual do Norte Fluminense, Avenida Alberto Lamego 2000, Parque Califórnia, CEP:28013-602. Campos dos Goytacazes, RJ, Brasil.
2Campo Experimental Santa Mônica, Embrapa Gado de Leite, Valença RJ, Brasil.
3Campo Experimental Santa Mônica, Empresa de Pesquisa Agropecuária do Estado do Rio de Janeiro- Pesagro-Rio, Valença RJ, Brasil.
*Corresponding author: Wilder H Ortiz. Centro de Ciências e Tecnologias, Universidade Estadual do Norte Fluminense, Avenida Alberto Lamego 2000, Parque Califórnia, CEP: 28013-602. Campos dos Goytacazes, RJ, Brasil. Tel. (22)2739-7055. Email: wilortvet@yahoo.es
Received: March 29, 2014; accepted: September 11, 2014
Summary
Background: candidate genes and their polymorphisms have been associated with traits of economic interest in cattle, representing an important strategy for genetic improvement of complex traits. Bone morphogenetic protein 4 (BMP4) is a member of the transforming growth factor beta (TGFβ) superfamily, which is involved with several events of embryonic, fetal, and adult development in vertebrates. Objective: the aim of this study was to evaluate the degree of association between polymorphism in the BMP4 gene (guanine for thymine- G>T, SNP rs109778173) and the performance of Gyr oocyte donors, including the rate of Cumulus-oophorus complex obtained in each session of follicular aspiration (OPU), embryo development, and pregnancy rates. Methods: DNA was extracted from hair follicles from 50 oocyte donors and genotyping was performed by polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP). Data from 212 OPU-IVP sessions was collected, and the following traits were associated with BMP4 polymorphism (SNP rs109778173): number and ratio of viable Cumulus-oophorus complexes; number of cleaved embryos at day 4 of culture; number of transferable embryos at day 7 of culture, and pregnancies on days 30 and 60 after embryo transfer. Results: the studied BMP4 polymorphism was significantly associated (p<0.01) with the number and ratio of viable cumulus-oocyte complexes, and the ratio of pregnancies at 30 days. Conclusion: the GT genotype (SNP rs109778173) was associated with inferior performance regarding OPU-IVP traits. This finding suggests possible genetic effects of BMP4 on performance of Gyr oocyte donors. Studies are needed on the BMP4 gene regarding subsequent changes in embryonic development of said mutation.
Keywords: genetic variability, molecular markers, mutation, OPU-IVP, SNPs.
Resumen
Antecedentes: genes candidatos y sus polimorfismos han sido asociados a características de interés económico en ganado, representando una excelente estrategia para el mejoramiento genético de características complejas. La proteína morfogénica ósea 4 (BMP4) es un miembro de la superfamilia del factor de crecimiento transformante beta (TGFβ); que controla innumerables eventos del desarrollo embrionario, fetal y de adultos en vertebrados. Objetivo: el objetivo de este estudio fue evaluar el grado de asociación entre un polimorfismo en el gen BMP4 (guanina por timina-G>T-, SNP rs109778173) y el desempeño de donantes de ovocitos de la raza Gyr, incluyendo la tasa de complejos cummulus-oophorus obtenidos en cada sesión de aspiración folicular (OPU), el desarrollo embrionario y la tasa de preñez. Métodos: el ADN fue extraído del folículo piloso de 50 vacas Gyr donantes de ovocitos, el genotipado fue realizado por la técnica de reacción en cadena de polimerasa-polimorfismos de longitud de fragmentos de restricción (PCR-RFLP). Datos de 212 sesiones de OPU-IVP fueron colectados y las características a seguir fueron asociadas con el polimorfismo en el gen BMP4 (SNP rs109778173): número y tasa de complejos de Cumulus-oophorus viables, de embriones clivados al día 4 de cultivo, de embriones transferibles al día 7 de cultivo, y de preñeces a los días 30 y 60 posterior a la transferencia de los embriones. Resultados: el polimorfismo fue significativamente asociado (p<0,01) con número y tasa de complejos Cumulus-oophorus viables y tasa de preñez al día 30. Conclusión: el genotipo GT (SNP rs109778173) fue asociado a resultados inferiores relacionados con características de OPU-IVP. Este resultado sugiere posibles efectos genéticos del gen BMP4 sobre el desempeño de donadoras de oocitos de la raza Gyr, siendo necesarios estudios del gen BMP4 en relación a las alteraciones en el desarrollo embrionario subsecuentes a la mutación citada.
Palabras clave: marcadores genéticos, mutación, OPU-IVP, SNPs, variación genética.
Resumo
Antecedentes: genes candidatos e seus polimorfismos têm sido associados a características de interesse econômico no gado bovino, representando uma importante estratégia para o melhoramento genético de características complexas. A proteína morfogênica óssea 4 (BMP4) é um membro da superfamília do fator de crescimento transformante beta (TGFβ); que controla diversos eventos do desenvolvimento embrionário, tanto fetal quanto adulto em vertebrados. Objetivo: avaliar o grau de associação entre um polimorfismo no gene BMP4 (guanina por timina-G>T-, SNP rs109778173) e o desempenho de doadoras de oócitos da raça Gyr, incluindo a taxa de complexos Cumulus-oophorus obtidos em cada sessão de aspiração folicular (OPU), o desenvolvimento embrionário e a taxa de prenhez dos embriões. Métodos: o DNA foi extraído do folículo piloso de 50 vacas Gyr doadoras de oócitos, e a genotipagem foi realizada pela técnica de reação em cadeia da polimerase-polimorfismo no comprimento de fragmentos de restrição (PCR-RFLP). Dados de 212 sessões de OPU-IVP foram coletados, e as características a seguir foram associadas com o polimorfismo no gene BMP4 (SNP rs109778173): número e taxa de complexos Cumulus-oophorus viáveis dos embriões clivados no dia 4 de cultivo, dos embriões transferíveis no dia 7 de cultivo e de prenhezes no dia 30 e 60 após a transferência dos embriões. Resultados: o polimorfismo estudado foi significativamente associado (p<0,01) com o número e a taxa dos complexos Cumulus-oophorus viáveis e a taxa de prenhezes ao dia 30. Conclusão: o genótipo GT (SNP rs109778173) foi associado a resultados inferiores relacionados a características OPU-IVP. Este resultado sugere possíveis efeitos genéticos do gene BMP4 sobre o desempenho de doadoras de oócitos da raça Gyr, fazendo-se necessários estudos no gene BMP4 em relação às alterações no desenvolvimento embrionário subsequentes ao polimorfismo citado.
Palavras chave: marcador molecular, mutação sinônima, OPU-IVP, SNP, variabilidade genética.
Introduction
Bone morphogenetic protein 4 (BMP4) is a member of the transforming growth factor beta (TGFβ) superfamily, which controls numerous events of embryonic, fetal and even adult development in vertebrates (Chen et al., 2004).
Several BMPs have been suggested as autocrine/ paracrine regulators of bovine ovarian follicular development, demonstrated by the expression of BMP4 and BMP7 in granulosa and theca cells (Glister et al., 2011).
During the follicular development, the BMP4 gene is expressed first in the stromal cells and later in the theca cells. Its receptors are found in granulosa cells as well as in the oocyte itself (Glister et al., 2004; Fatehi et al., 2005). Those receptors participate in primordial to primary follicular transition, stimulate granulosa cells proliferation, pre-antral follicular growth, and follicular survival, and regulate steroideogenesis in granulosa cells (Knight and Glister, 2006).
There is new evidence to support the hypothesis that BMP4 and BMP7 originated from theca and stromal cells to promote the initial transition of primordial follicles to enhance follicle survival (Phil and Claire, 2006). Moreover, in later developmental stages, BMP4 induces embryonic and extra embryonic mesoderm formation (Degrelle et al., 2011) and is also related to vasculogenesis (Astorga and Carlsson, 2007) in the embryo and in the developing placenta as well. As a result, BPMs are of major importance for pregnancy success (La Rosa et al., 2011). These authors also concluded that BMP4 is implicated in oocyte maturation and embryonic development in bovines, affecting the cleavage rates and the pluripotent state of embryonic cells.
Since the introduction of ovum pick-up/in vitro production (OPU-IVP), many efforts to improve embryo production efficiency have been undertaken. These were all focused on non-genetic factors. The genetic component to IVP was approached by Machado et al., (2006), suggesting its influence on the characteristics associated with IVP and, therefore, their heritability.
Genome-wide association studies on economic aspects have shown a clear genetic divergence between Bos taurus and Bos indicus (Fortes et al., 2010; Canavez et al., 2012; Höglund et al., 2012). Previous studies have reported an association between single nucleotide polymorphism (SNP) in BMP4 gene and the blastocyst rates in Holstein cows evaluated in vitro (Li et al., 2012), suggesting a major effect of BMP4 on the performance of oocyte donors. In this study we evaluated the association between G>T, SNP rs109778173 polymorphism in BMP4 gene and in vitro production traits and pregnancy rate of Gyr cows.
Material and methods
Ethical considerations
This study was approved by the Ethics Committee for Animal Experimentation of Universidade Estadual do Norte Fluminense-UENF (Protocol number 243, March 11, 2014).
DNA samples and data collection
DNA was extracted from hair follicles via alkaline method, according to standard procedure (adapted from Coelho, 2001). DNA concentrations were measured using a NanoDropTm 2000 (Thermo Science, New York, NY, USA) spectrophotometer. Data from 212 OPU-IPV sessions of 50 Gyr oocyte donors were collected in 2012 and 2013 (Rio de Janeiro, Brazil). The OPU-IVP procedures were performed by two companies with extensive experience in the field.
The OPU-IVP traits associated with the SNP marker were: number and ratio of viable cumulusoophorus complexes (Nvcoc and Pvcoc) classes I, II, and III that were successfully matured, fertilized, and in vitro cultured. Number and ratio of cleaved embryos at day 4 of culture (Ncleavd4 and Pcleavd4). Number and ratio of transferable embryos at day 7 of culture (Ntembd7 and Ptembd7) as a ratio of Nvcoc, according to the international embryo transfer society (IETS) classification as stage 4 (morula), grade 1 (excellent or good); and stages 5 to 7 (early blastocyst, blastocyst, and expanded blastocyst), grade 1 and 2 (excellent and regular).
Besides the characteristics mentioned above, SNP marker was also associated with number and ratio of pregnancies at days 30 and 60 after transfer (NPrD30, PPrD30 and NPrD60, PPrD60). All characteristics are described considering each OPU-IPV session. Data for all characteristics were recorded and ceded for research purposes.
Single nucleotide polymorphism analysis
Detection of BMP4 polymorphism was carried out according to Li et al. (2012). A segment of the BMP4 gene (exon 2, SNP rs109778173) was amplified by PCR with a specific primer (Table 1). The PCR reaction was performed in a final volume of 20 μl, using 1x PCR buffer [10 mM Tris-HCI (including Mg+2)], 0.5 mM dNTP mix (Promega, Madison,WI, USA), 1U of Taq DNA polymerase (Promega, Madison, WI, USA), 0.5 mM of each primer (Invitrogen, Sao Paulo, SP, Brazil), 50 ng of DNA extracted, and deionized water. A negative control was included for each replicate of the PCR reaction.

The cycling conditions consisted of an initial state of 95 °C for 1 min, followed by 40 amplification cycles with denaturation at 95 °C for 30 s, annealing of primers at 55 °C for 1 min, and extension at 72 °C for 1 min. After the last cycle, reactions were led to a final step of 7 min at 72 °C for the final extension of the tapes. PCR was conducted using a thermocycler (Applied Veriti® 96-Well, Applied Biosystems, Foster City, CA, USA). The product was separated by polyacrylamide gel (8%) stained with silver nitrate to confirm the amplification (Sigma-Aldrich, Sao Paulo, SP, Brazil).
Polymerase chain reaction-restriction-fragment length polymorphism (PCR-RFLP) was used for genotyping the amplified samples. The restriction enzyme HinfI used (Life technologies, Sao Paulo, SP, Brazil) recognizes and cleaves the 5' G ̌ ANTC 3' sequence (Table 2). This enzyme recognizes only one cleavage site in the amplified fragment, with a reaction product of two fragments sized 80 bp and 110 bp. The expected result was cleavage of the samples that did not display the G>T mutation. The cleavage reaction was carried out in a final volume of 20 μl, including HinfI specific buffer, 5 IU restriction enzyme, 4 μl PCR product, and deionized water.

To confirm the digestion reaction, the product was separated by polyacrylamide gel (8%) and stained with silver nitrate (Sigma-Aldrich). After this visualization, the samples were submitted to capillary electrophoresis using the equipment for allelic discrimination by size and polymorphism identification (Fragment Analyzer™, Advanced Analytical, Ames, IA, USA).
Statistical analysis
Genotypic and allele frequency, Hardy-Weinberg equilibrium probability (P-HW), gene homozygosity and heterozygosity (Ho and He), effective allele number (Ne) and polymorphism information content (PIC) were statistically analyzed according to the approaches by Nei and Roychoudhury (1974) and Nei and Li (1979). These tests were conducted using PowerMarker v.3.25 software.
The association between SNP marker and OPUIVP traits and pregnancy rates was determined by analysis of variance of repeated data using the following model:
Yij = μ + Ei + εij
Where:
Yij is the observation for the OPU-IVP traits.
μ is the mean for each trait.
Ej is the genotype effect.
εij is the random error.
This model was analyzed using the PROC GLIMMIX procedure of SAS 9.2 program (SAS Institute Inc., Cary, NC, 1999).
Results
The protein coding domain of BMP4 gene is located into exon 2. We amplified a fragment of DNA sequence in exon 2, where the SNP mutation number rs109778173 was identified in Holstein cows (Li et al., 2012). We found the mutation G>T in Gyr cows using PCR-RFLP and capillary electrophoresis methods. Three genotypes were identified (GG, TT and GT) with genotypic frequencies of 0.64, 0.32, and 0.04, respectively. Allelic frequency and Hardy- Weinberg equilibrium are described in Table 3.

The results displayed in Table 4 show heterozygosity, polymorphism information content, and effective number of alleles with medium values. This data points out that polymorphism levels and genetic variation were medium, according to the classification of PIC (low polymorphism if PIC value <0.25, medium polymorphism if 0.25<PIC value <0.5, and high polymorphism if PIC value >0.5). The Chi-squared test showed that genotypic distributions in the population studied agreed with Hardy-Weinberg equilibrium (p>0.05).
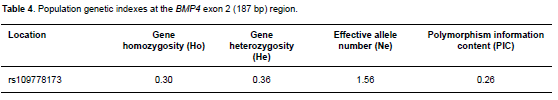
Medium values of SNP polymorphism for the characteristics above mentioned indicate its potential use as a molecular marker (Fu et al., 2013).
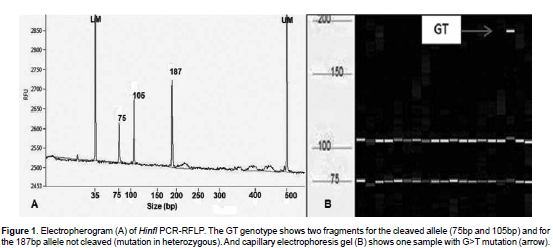
Electropherogram and capillary electrophoresis gel for G>T mutation in BMP4 gene are shown in Figure 1. Alleles of 75 bp and 105 bp were obtained after cleavage with HinfI restriction enzyme. The mutant allele was not cleaved because the recognizing site was absent, and the allele size was 187 bp, leading us to conclude that the mutation was present in heterozygous.
Polymorphism variance analysis for OPU-IVP traits and pregnancy rates at rs109778173 -BMP4 gene exon 2 (187 bp) locus showed significant effect of genotype GT and characteristics of number and ratio of viable Cumulus-oophorus complexes (Nvcoc and Pvcoc) and ratio of pregnancies at 30 days (PPrD30). Means of GT genotype showed lower performance for the characteristics listed above at probability level of 1% (Table 5). The GT genotype might be an unfavorable genotype.

The mean values for the other characteristics evaluated showed no significant effect depending on genotype, although all of them were numerically lower in mutant genotype when compared with other genotypes (GT vs. GG and TT). Genotypes GG and TT showed no significant effect on OPU-IVP traits and pregnancy rates.
Discussion
The BMP4 gene, located at chromosome 10 can be found in humans, chimpanzees, rhesus monkeys, dogs, mice, rats, and zebra fish. B. taurus is one of 65 orthologous species with the human BMP4 gene (Waterhouse et al., 2011).
Involvement of bone morphogenetic proteins in development processes of embryonic tissues has been documented starting from the gastrulation process. Gastrulation is a highly dynamic process. When gastrulation evolves properly, it allows two major events of early development of vertebrates: I) the establishment of morphologically visible anteroposterior (head or tail) and dorsal-ventral axis of the cord (or mouth-spinal), and II) the specification and standardization of early germ layers, especially the mesoderm (Downs and Davies, 1993).
Mutants obtained during the gastrulation process recently revealed that the axis of the embryo proper specification consists of a series of reciprocal interactions between the embryonic and extraembryonic tissues, so genes involved in these interactions can then contribute to the morphogenic development of the embryo (Pfister et al., 2007; Tam and Loebel, 2007).
Animals displaying G>T mutation in BMP4 gene in this study presented 4.29 less viable oocytes per OPU session compared to the GG genotype, which represents a 16.23% loss to the TT genotype. Also, pregnancy rate at 30 days from transferred embryos obtained from the GT cows was 26.5% lower compared to the TT genotype.
Li et al. (2012), using an in vitro model in Holstein cows, showed similar results for the G>T mutation in BMP4 gene. They concluded that embryos produced from genotype TT cows showed 10.5 and 16.1% higher blastocyst rates than GG and GT cows, respectively. The same authors showed that blastocyst rate was significantly associated with SNP rs109778173 of BMP4 (p = 0.006), whereas the association with fertilization rate was not statistically significant (p = 0.095).
La Rosa et al. (2011) demonstrated that BMP4 slows down the development of only in vitro fertilized embryos, and concluded that it influences bovine in vitro oocyte maturation and affects cleavage rates and the pluripotent state of embryonic cells, suggesting that a balance of BMP signaling is needed for a proper development during pre-implantation of bovine embryos.
The genotypic frequency in the Gyr cows population studied was 0.64, 0.32 and 0.04 for the GG, TT and GT genotypes, respectively, concluding that the GT genotype is less present, in contrast with the genotypic frequencies observed by Li et al. (2012) (approximately 0.56, 0.05, and 0.37, respectively). Therefore, the GT genotype has a higher ratio for the BMP4 gene (SNP rs109778173) in Holstein cows, compared with the same genotype in the Gyr population studied in this work.
Probability values for the Hardy-Weinberg equilibrium (p>0.05) in both populations show disequilibrium with the evaluated SNP, and therefore, with their ability to segregate (Ardlie et al., 2002).
We hypothesized that the difference between genotypic frequencies for GT in two populations could be associated with selection processes aimed at increasing milk production. However, studies in both populations (Holstein and Gyr) involving SNPs related to milk production and fertility must be conducted to test the hypothesis of joint segregation.
The widespread use of artificial insemination in Nordic Red cattle has resulted in increased selection intensity, leading to increased productivity. However, cow fertility has concurrently severely declined. Kadri et al. (2014) identified a 660-kb deletion encompassing four genes as the causative variant. They showed that the deletion is a recessive embryonically lethal mutation. This study demonstrates that embryonic lethal mutations account for a non-negligible fraction of the decline in fertility of domestic cattle, and that associated positive effects on milk yield may account for part of the negative genetic correlation.
The G>T mutation at SNP rs109778173 is classified as silent or synonymous, given that the replacement of guanine by thymine nucleotide in codon (CTG by CTT) does not alter the amino acid sequence in the protein (in the case of the protein coding of BMP4 gene it is leucine). This phenomenon is often referred to as redundancy of the genetic code.
Despite this redundancy, synonymous codons are not used with equal frequency due to a phenomenon known as codon bias (Ikemura, 1981). If codon bias is the result of natural selection, a change from a preferred to an undesirable codon should lead to reduced protein expression, caused by a decrease in the efficiency or fidelity of translation or a combination of both (Bulmer, 1991). Adoligbe et al. (2012) showed a significant effect for synonymous mutation in GDF10 gene with body traits in indigenous Chinese cattle.
When a synonymous or silent mutation occurs, the change is often assumed to be neutral, meaning that it does not affect the fitness of the individual carrying the new gene to survive and reproduce. Synonymous changes may not be neutral because certain codons are translated more efficiently (faster and/or more accurately) than others (Carlini and Stephan, 2003).
The association between BMP4 gene polymorphism and OPU-IVP traits in Gyr cattle confirmed the influence of this gene in embryonic development and fertilization rates after embryo transfer. The GT genotype negatively affects characteristics such as number and ratio of viable Cumulus-oophorus complexes and ratio of pregnancies at day 30 after transfer. This finding could be an indication of a genetic effect of BMP4 on these characteristics.
Acknowledgements
The authors wish to thank Embrapa-Gado de Leite- CESM and PECGEN-embriões for providing the data used in this study, as well as Universidade Estadual do Norte Fluminense-UENF for a scholarship awarded to the first author, and to CNPq for financial support.
Conflicts of interest
The authors declare they have no conflicts of interest with regard to the work presented in this report.
Notes
¤To cite this article: Ortiz WH, Quirino CR, Silva A, Oliveira CS, Serapião RV, Pacheco A, Bartholazzi A. Association between BMP4 gene polymorphism and in vitro embryo production traits in Gyr cows. Rev Colomb Cienc Pecu 2015; 28:156-164.
References
Adoligbe C, Zan L, Farougou S, Wang H, Ujjan JA. Bovine GDF10 gene polymorphism analysis and its association with body measurement traits in Chinese indigenous cattle. Mol Biol Rep 2012; 39:4067-4075. [ Links ]
Ardlie KG, Kruglyak L, Seielstad M. Patterns of linkage disequilibrium in the human genome. Nat Rev Genet 2002; 3:299-309. [ Links ]
Astorga J, Carlsson P. Hedgehog induction of murine vasculogenesis is mediated by Foxf1 and BMP4. Development 2007; 134(20):3753-3761. [ Links ]
Bulmer M. The selection-mutation-drift theory of synonymous codon usage. Genetics 1991; 129:897-907. [ Links ]
Canavez F, Luche D, Stothard P, Leite K, Sousa-Canavez J, Plastow G. Genome sequence and assembly of Bos indicus. J Hered 2012; 103:342-348. [ Links ]
Carlini DB, Stephan W. in vivo introduction of unpreferred synonymous codons into the Drosophila ADH gene results in reduced levels of ADH protein. Genetics 2003; 163:239-243. [ Links ]
Chen D, Zhao M, Mundy GR. Bone morphogenetic proteins. Growth Factors 2004; 22(4):233-241. [ Links ]
Coelho GA. Analise quali quantitativa de tecnicas para extração de DNA de sangue sêmen e pêlos. Belo Horizonte, Minas Gerais; 2001. [ Links ]
Degrelle SA, Cao KA L, Heyman Y, Everts RE, Campion E, Richard C. A small set of extra-embryonic genes defines a new landmark for bovine embryo staging. Reproduction 2011; 141(1):79-89. [ Links ]
Downs K, Davies T. Staging of gastrulating mouse embryos by morphological landmarks in the dissecting microscope. Development 1993; 118:1255-1266. [ Links ]
Fatehi A, Van den Hurk R, Colenbrander B, Daemen AJ, van Tol HTA, Monteiro RM. Expression of bone morphogenetic protein2 (BMP2), BMP4, and BMP receptors in the bovine ovary but absence of effects of BMP2 and BMP4 during IVM on bovine oocyte nuclear maturation and subsequent embryo development. Theriogenology 2005; 63(3):872-879. [ Links ]
Fortes MR, Reverter A, Zhang Y, Collis E, Nagaraj SH, Jonsson NN. Association weight matrix for the genetic dissection of puberty in beef cattle. PNAS 2010; 107(31):13642-13647. [ Links ]
Fu C, Wang H, Mei C, Wang J, Jiang B, Ma X. SNPs at 3ꞌ-UTR of the bovine CDIPT gene associated with Qinchuan cattle meat quality traits. Genet Mol Res 2013; 12(1):775-782. [ Links ]
Glister C, Kemp C, Knight P. Bone morphogenetic protein (BMP) ligands and receptors in bovine ovarian follicle cells: actions of BMP-4, -6 and -7 on granulosa cells and differential modulation of Smad-1 phosphorylation by follistatin. Reproduction 2004; 127(2):239-254. [ Links ]
Glister C, Satchell L, Knight PG. Granulosal and thecal expression of bone morphogenetic protein- and activin-binding protein mRNA transcripts during bovine follicle development and factors modulating their expression in vitro. Reproduction 2011; 142(4):581-591. [ Links ]
Höglund JK, Guldbrandtsen B, Lund MS, Sahana G. Analyzes of genome-wide association follow-up study for calving traits in dairy cattle. Bmc Genet 2012; 13:1471-2156. [ Links ]
Ikemura T. Correlation between the abundance of Escherichia coli transfer RNAs and the occurrence of the respective codons in its protein genes: a proposal for a synonymous codon choice that is optimal for the E. coli translational system. J Mol Biol 1981; 151:389-409. [ Links ]
Kadri NK, Sahana G, Charlier C, Iso-Touru T, Guldbrandtsen B, Karim L. A 660-Kb Deletion with antagonistic effects on fertility and milk production segregates at high frequency in Nordic Red Cattle: additional evidence for the common occurrence of balancing selection in livestock. PLOS Genet 2014; 10(1):e1004049. [ Links ]
Knight P, Glister C. TGF-beta superfamily members and ovarian follicle development. Reproduction 2006; 132(2):191-206. [ Links ]
La Rosa I, Camargo LS, Pereira MM, Fernandez-Martin R, Paz DA, Salamone DF. Effects of bone morphogenic protein 4 (BMP4) and its inhibitor, Noggin, on in vitro maturation and culture of bovine preimplantation embryos. Reprod Biol Endocrin 2011; 1:9-18. [ Links ]
Li G, Khateeb K, Schaeffer E, Zhang B, Khatib H. Genes of the transforming growth factor-beta signaling pathway are associated with pre-implantation embryonic development in cattle. J Dairy Res 2012; 79:310-317. [ Links ]
Machado S, Reichenbach H, Weppert M, Wolf E, Gonçalves PB. The variability of ovum pick-up response and in vitro embryo production from monozygotic twin cows. Theriogenology 2006; 65:573-583. [ Links ]
Nei M, Li W. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc Natl Acad Sci 1979; 76:5269-5273. [ Links ]
Nei M, Roychoudhury A. Sampling variances of heterozygosity and genetic distance. Genetics 1974; 76:379-390. [ Links ]
Pfister S, Steiner K, Tam P. Gene expression pattern and progression of embryogenesis in the immediate post-implantation period of mouse development. Gene Expr Patterns 2007; 7:558-573. [ Links ]
Phil GK, Claire G. TGF-b superfamily members and ovarian follicle development. Reproduction 2006; 132:191-206. [ Links ]
SAS Institute Inc. SAS/STAT 9.2 User's Guide. Cary, NC: SAS Institute Inc; 1999. [ Links ]
Tam PP, Loebel DA. Gene function in mouse embryogenesis: get set for gastrulation. Nat Rev Genet 2007; 8(5):368-381. [ Links ]
Waterhouse RM, Zdobnov EM, Tegenfeldt F, Li J, Kriventseva EV. OrthoDB: the hierarchical catalog of eukaryotic orthologs in 2011. Nucleic Acids Res 2011, 39:283-288. [ Links ]
