











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
La estructura trófica de las comunidades resume en gran medida cómo la energía fluye a través de los diferentes compartimentos de los ecosistemas (Vannote et al., 1980; Cuevas y Medina, 1988). Por lo tanto, el estudio de la estructura trófica de una o más comunidades permite entender de mejor manera cómo se da la transferencia de energía y materia en los ecosistemas y, a su vez, caracterizar relaciones básicas como la depredación o la herbivoría entre las especies pertenecientes a una comunidad (Jepsen y Winemiller, 2002; Andramunio-Acero y Caraballo, 2012). Un ejemplo es en la caracterización de la abundancia y diversidad del grupo de los fragmentadores dentro de la comunidad de los macroinvertebrados acuáticos. Esto debido a que las especies de este grupo cumplen un papel importante en la transformación de la materia orgánica, pues cortan y degradan el material foliar, lo que aumenta las tasas de descomposición. Dicha actividad repercute directamente sobre la biomasa y la producción primaria del sistema (Ramírez y Gutiérrez-Fonseca, 2014; Villada-Bedoya et al., 2017).
A pesar de su importancia, son pocos los trabajos que se han realizado en ecosistemas acuáticos neotropicales, siendo uno de los más relevantes el realizado por Tomanova et al. (2006), quiénes describieron los grupos tróficos de macroinvertebrados acuáticos que coexisten en cuatro ríos tropicales de Bolivia. En Colombia, son escasos los estudios que han abordado la estructura trófica en ecosistemas acuáticos y la mayoría se han realizado en regiones montañosas del país. Chará-Serna et al. (2010) por ejemplo, describieron la estructura trófica de insectos acuáticos en ocho quebradas de la región cafetera y Rivera-Usme et al. (2013) determinaron los grupos tróficos en humedales urbanos Andinos. Por otra parte, Córdoba-Ariza et al. (2020) describieron la estructura trófica en tres ríos de la Guayana colombiana. De igual manera, en un estudio elaborado por Rodríguez-Barrios et al. (2011) se realizó una caracterización de los grupos funcionales alimentarios en el río Gaira en la región Caribe, que luego fue complementada por Tamaris-Turizo et al. (2020). En general, estas contribuciones muestran que existe una amplia variabilidad en la utilización de recursos entre los macroinvertebrados acuáticos. De hecho, es frecuente encontrar una alta plasticidad dentro de un mismo grupo taxonómico (p.e., dentro de la familia Hydropsychidae, Tamaris-Turizo et al., 2020).
La alta variabilidad observada en los grupos tróficos entre los distintos estudios resalta en gran medida la falta de información de la estructura trófica en los ecosistemas acuáticos colombianos. Por ello, se deben aunar esfuerzos para caracterizar la estructura trófica en un mayor número de ecosistemas y, en especial, en aquellos ubicados en regiones poco exploradas como las sabanas y las zonas inundables en la Orinoquía y la Amazonía. En Colombia, una sexta parte de los ecosistemas acuáticos están asociados a las llanuras aluviales. Estas son zonas de sedimentación fluvial en donde la forma y estructura de los cauces se crean por la depositación de sedimentos provenientes de los ríos de zonas altas (González-Trujillo y Alonso-Moreno, 2020). En estas zonas se asientan ecosistemas como los bosques de galería, los cuales juegan un papel preponderante en las dinámicas de los ríos y arroyos, ya que contribuyen con recursos alóctonos que dan forma a la estructura de las redes tróficas y, en especial, a la estructura trófica de las especies que conforman las comunidades acuáticas (Junk et al., 1989). Sin embargo, estos aún son ríos cuya ecología no ha sido totalmente explorada.
Este estudio tuvo como objetivo caracterizar la estructura trófica de las comunidades de macroinvertebrados acuáticos en una llanura inundable de la Orinoquía colombiana, esto con el fin de aportar información que permita expandir el conocimiento de la estructura y funcionamiento de estos ecosistemas y su variabilidad espacio-temporal.
MATERIALES Y MÉTODOS
Área de estudio
Las llanuras aluviales de la Orinoquía son zonas cálidas y bastante húmedas, poseen lluvias entre 2500 y 3300 mm, una humedad relativa aproximada al 60 % y temperaturas que oscilan entre 26 y 28 ° C en las épocas de sequía. Estas llanuras se caracterizan por inundaciones constantes o temporales a lo largo del año, lo cual hace que existan interacciones altamente dinámicas entre los componentes terrestre y acuático, contribuyendo así a la conectividad entre diferentes unidades ecológicas (Briceño, 2014). Este estudio se llevó a cabo en la Hacienda Matepantano, ubicada a 15 km del casco urbano del municipio de Yopal-Casanare (5°19'13.4" N - 72°17'47.4" W). La región presenta un régimen monomodal de lluvias con un primer período de sequía entre diciembre y marzo y otro de lluvias intensas entre abril y noviembre. Las campañas de muestreo se realizaron en los arroyos Las Flores (5°19'34" N - 72°17'51), El Tiestal (5°19'13" N - 72°17'47) y El Güio (5°18'56" N - 72°18'09) en los meses de julio - aguas altas (2016), octubre - aguas en descenso (2016) y enero - aguas bajas (2017).
Fase de campo
Se realizó la medición de las diferentes variables fisicoquímicas por triplicado en los arroyos: la temperatura (°C) y el oxígeno disuelto (mg l-1 O2) se midieron con un oxímetro (Hach HQ30); también se evaluaron la conductividad (µS cm-1) y el pH con una sonda multiparamétrica (Eijkelkamp 18.52.01) y la velocidad de la corriente de agua con un medidor global de flujo digital y de descarga (Q). La determinación del amonio (NH4 +, mg/L) y del fosfato (PO4 -) se hizo por el método de colorimetría (MColortest - Compact Laboratory for Water Testing Freshwater Test Kit). Para el muestreo de macroinvertebrados se delimitaron tramos de 50 metros en dos puntos distintos de los arroyos (aguas arriba y aguas abajo). En cada tramo se recolectaron cinco muestras tomadas con una red Surber (900 cm2 con un ojo de malla de 250 |m) sobre sustratos rocosos (Rocas-Cantos rodados-Guijarros y gravas). Adicionalmente, se tomó una muestra integral en los sustratos rocosos del tramo, para incluir la mayor cantidad de organismos presentes en el cuerpo de agua. Las muestras se congelaron previo a la fijación con alcohol, con el objetivo de que los organismos preservaran sus contenidos estomacales. Las muestras se preservaron en etanol al 70 %.
Fase de laboratorio
El material obtenido se identificó hasta el nivel taxonómico más bajo posible (género principalmente) mediante las claves taxonómicas de Merrit et al. (2008), Domínguez y Fernández (2009) y Trivinho-Strixino (2011). Para el orden Ephemeroptera y la familia Chironomidae, se realizó un aclarado previo con una solución caliente de hidróxido de potasio (KOH, 10 %); posteriormente la muestra se deshidrató en alcohol al 70 % y 96 % y el material se montó en Euparal® de acuerdo con lo establecido por Prat et al. (2014). Los especímenes se depositaron en el museo de Historia Natural de la Universidad de La Salle (MLS).
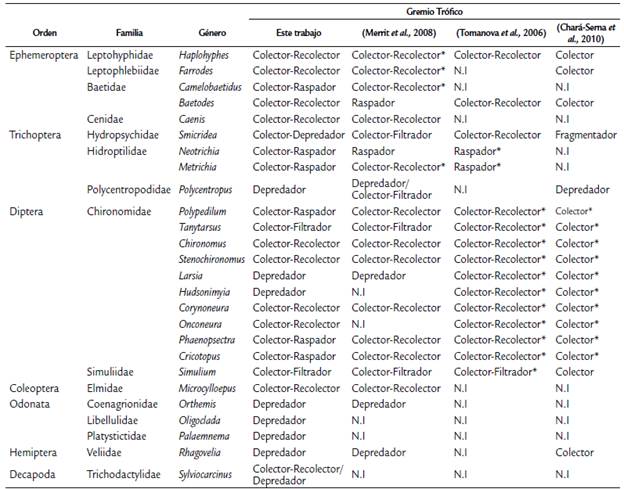
Para la determinación de la dieta de los diferentes grupos tróficos, se siguió el método planteado por Tomanova et al. (2006) y Muñoz et al. (2009). De esta manera, se seleccionaron cinco individuos por taxón, luego se diseccionaron bajo un estereoscopio (Zeiss Stemi DV4) y se extrajo su tracto digestivo; los organismos se abrieron longitudinalmente o solamente estirando la cabeza. Posteriormente, se realizaron los montajes con alcohol polivinílico en portaobjetos; el material tuvo que homogenizarse para garantizar la dispersión del contenido estomacal sobre la placa. Las muestras se observaron bajo un microscopio (Zeiss Primo Star) en 40X. En cada muestra se seleccionaron 20 campos al azar y se tomaron fotografías del contenido del tracto digestivo representativo e identificable. Se identificaron los siguientes ítems alimenticios: materia orgánica fina, materia orgánica gruesa, restos de animales terrestres, restos de animales acuáticos y algas. Después de tomar cada foto, y con ayuda del programa ImageJ, se calculó el área ocupada por el material; los resultados se transfirieron al software de Microsoft Office Excel 2007, en el cual se calculó la proporción de cada tipo de alimento con respecto al total. De acuerdo a los porcentajes de cada ítem alimenticio y a la morfología de las partes bucales, los diferentes géneros se categorizaron en los grupos tróficos establecidos por Tomanova et al. (2006), Merrit et al. (2008) y Chará-Serna et al. (2010). Esto son: Colectores-Recolectores (C-R), Depredadores (D), Colectores-Filtradores (C-F) y Colectores-Raspadores (C-Ra). No fue posible observar el contenido estomacal en seis géneros, por lo que sus grupos tróficos se asignaron de acuerdo con las referencias bibliográficas de Tomanova et al. (2006) y Merrit et al. (2008). Estos géneros fueron: Microcylloepus, Orthemis, Oligoclada, Palaemnema, Rhagovelia y Polycentropus.
Tabla 2 Caracterización de los grupos tróficos de Macroinvertebrados en tres arroyos de la llanura inundable colombiana. La asignación de cada género se compara con lo registrado por Merrit et al. (2008), Tomanova et al. (2006) y Chará-Serna et al. (2010). *Clasificación establecida a nivel de familia. N.I.: No hay información.
Análisis estadísticos
Mediante el programa PAST Versión 3 (Hammer et al., 2001) se realizó una prueba ANOVA para establecer posibles diferencias temporales entre los arroyos en términos de sus características físicas y químicas, así como para conocer las diferencias entre los índices de diversidad entre los arroyos y entre las temporadas de muestreo. Para probar los supuestos de normalidad y homogeneidad de los datos, se realizó la prueba de Shapiro-Wilk; en caso de no cumplirse estos supuestos, se utilizó la prueba no paramétrica de Kruskal-Wallis (K-W) para determinar si la abundancia relativa de los organismos y de los grupos tróficos difería entre los arroyos y entre los meses. Ambas pruebas se realizaron en el programa PAST Versión 3 y el programa R 3.6.1 (Hammer et al., 2001; R Core Team, 2019).
Finalmente, para explorar la influencia de las variables físicas y químicas sobre la estructura y composición de los grupos tróficos, se realizó un análisis de redundancia (RDA). El poder explicativo de cada variable se evaluó mediante una prueba de Montecarlo (999 permutaciones, p-valor <0,001). El RDA se realizó con el programa CANOCO 4.5 (Ter Braak y Smilauer, 2002). Los datos se transformaron con el logaritmo (Log 10 +1) para estabilizar la varianza de los datos (Anderson y Whitcomb, 2001).
RESULTADOS
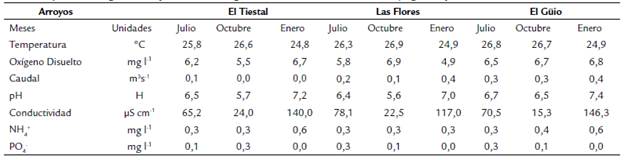
Los arroyos registraron una temperatura promedio de 26 °C y un pH de 6.5, ligeramente ácido; las variables que presentaron cambios significativos en los diferentes meses de muestreo fueron la conductividad, que aumentó durante la época de aguas bajas (ANOVA, F=10,38, p-valor <0,01) y la concentración de fósforo reactivo soluble (ANOVA, F=4,22, p-valor <0,01), que disminuyó en el mes de enero (Tabla 1).
Diversidad de macroinvertebrados
En total se contaron e identificaron 2396 individuos, representados en 42 géneros, correspondientes a 23 familias y ocho órdenes (Fig. 1). La familia Chironomidae fue la más abundante, constituida principalmente por los géneros Corynoneura, Hudsonimyia, Tanytarsus, Larsia, Onconeura, Polypedilum y Phaenopsectra. Se observaron diferencias significativas en la abundancia relativa de los grupos taxonómicos en los tres arroyos (K-W=7,2816, fd=2, p <0,01) y durante los diferentes periodos climáticos (K-W= 13,876, fd= 8, p <0,01). El arroyo El Tiestal presentó los valores más altos (2,89 bits) con relación al índice de Shannon (H'), mientras que las diversidades más bajas se registraron en el arroyo Las Flores para el mes de octubre (1,92 bits); sin embargo, no se encontraron diferencias significativas entre los meses ni entre los arroyos muestreados (p=0,14; F=4,24).

Estructura trófica
En total se identificaron diez géneros de Colectores-Recolectores, seis de Colectores-Raspadores, ocho de Depredadores, dos de Colectores-Filtradores y uno del grupo Colector-Depredador. Dentro de los grupos tróficos no se encontraron organismos que pudieran clasificarse como Fragmentadores (Tabla 2). La familia Chironomidae fue la que tuvo la mayor diversidad de grupos tróficos.
Cambios espaciales y temporales
En la Figura 2 se evidencia que durante el mes de enero, correspondiente al periodo de aguas bajas, hubo una dominancia de organismos Colectores-Recolectores en los arroyos El Tiestal y El Güio. Julio, mes de aguas altas, presentó un alto porcentaje de Colectores-Recolectores en Las Flores (46 %), en El Tiestal del (58,7 %) y en El Güio el (47 %), mientras que los Raspadores-Colectores tuvieron baja representatividad en los arroyos El Tiestal y Las Flores en ese mes. En octubre (aguas en descenso) se observó un aumento de organismos Depredadores (57 %) en el arroyo Las Flores, además de una baja representatividad de Colectores-Raspadores en Las Flores (0,9 %) y en El Tiestal (1,8 %). En enero hubo un aumento general de los grupos tróficos, que correspondió, como se mencionó, a los organismos Colectores-Recolectores. En general, estos fueron el grupo más representativos en todos los arroyos (Fig 2).
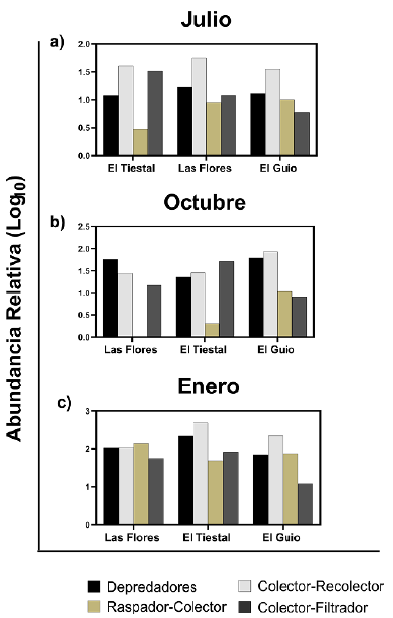
Fig. 2 Variación espacial y temporal de los grupos tróficos de macroinvertebrados. a) Julio (aguas altas), b) Octubre (aguas en descenso), c) Enero (aguas bajas).
El análisis de redundancia (Fig 3) mostró que los dos primeros ejes explicaron conjuntamente el 91 % del total de la varianza de los datos, siendo el caudal (p <00,1; F=3,91) y la temperatura (p <0,001; F=4,86) las variables más significativas en el modelo. El primer eje se asoció negativamente con el caudal y el segundo se relacionó negativamente con la temperatura. Los Colectores-Recolectores, Depredadores y Colectores-Raspadores se relacionaron con caudales altos, mientras que los Colectores-Filtradores lo hicieron con las altas temperaturas del agua, principalmente en la época seca (enero).
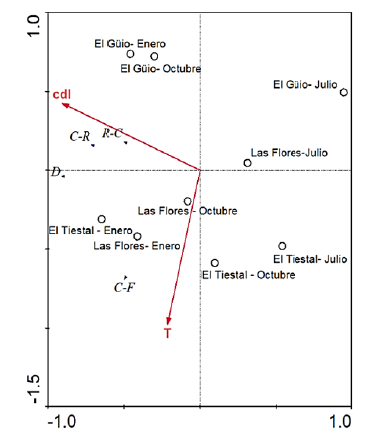
Fig. 3 Representación gráfica de la tripleta de variables (grupos tróficos, arroyos y características ambientales) modeladas por el análisis de redundancia (RDA). El ángulo entre vectores representa el grado de correlación entre variables ambientales y su correlación con los ejes del RDA. C-R (Colectores-Recolectores), R-C (Colectores-Raspadores), D (Depredadores) y C-F (Colector-Filtrador). T: temperatura, cdl: caudal.
DISCUSIÓN
De acuerdo con lo hallado en la bibliografía, este trabajo presenta una de las primeras caracterizaciones de la variabilidad espacial y temporal en la estructura trófica de las comunidades de macroinvertebrados acuáticos de la llanura aluvial de la Orinoquía colombiana. Se encontró una alta representatividad de organismos Colectores - Recolectores en todos los meses y arroyos estudiados. Así mismo, en cuanto a las variaciones físicas e hidrológicas, se vio que tuvieron un notable impacto en la abundancia de todos los grupos tróficos, en especial en la épocas de aguas bajas. Se resalta el papel que juegan los Depredadores, pues tuvieron una alta abundancia, probablemente por el aumento general de los otros grupos. De acuerdo con los resultados, la estructura de las comunidades de macroinvertebrados de estos ríos de la llanura aluvial parece responder a la variabilidad hidrológica de la región, siendo especialmente cambiantes la abundancia y riqueza de algunos grupos tróficos. El grupo de los Colectores-Recolectores, por ejemplo, fue muy abundante y diverso durante los meses de julio (aguas altas) y enero (aguas bajas), mientras que el grupo de los Colectores-Raspadores lo fue en enero, mes que corresponde a la época de menor intensidad de lluvias. Sin embargo, cabe resaltar que el análisis de los contenidos estomacales sugiere que los taxones de macroinvertebrados acuáticos de la zona de estudio, ubicada en la llanura aluvial, tienen una alta flexibilidad alimenticia, lo que les permite adaptarse al tipo de alimento disponible en el medio. Dicha flexibilidad es más evidente cuando se analizan los contenidos estomacales al nivel taxonómico de familia. Un claro ejemplo se da dentro de la familia Chironomidae, en la cual se encontraron géneros de las categorías Colectores-Recolectores, Depredadores y Colectores-Raspadores.
En este estudio se evidenció que existe una relación inversa entre el periodo hidrológico y el caudal, dado que los caudales fueron bajos incluso en el periodo de aguas altas. Esto podría deberse a que estos arroyos se encuentran próximos a zonas de conducción, bombeo y extracción de agua para consumo doméstico (Briceño, 2014). Sin embargo, el caudal influyó sobre la abundancia y diversidad de los grupos tróficos. Intentando entender esta relación, algunos autores sugieren que la variabilidad hidrológica es una covariable esencial para modelar la estructura trófica de las comunidades acuáticas, ya que incide sobre procesos como la acumulación y descomposición de la hojarasca o la heterogeneidad de los sustratos, lo cual influye en la presencia y abundancia de determinados grupos tróficos (González-Trujillo, 2016; Ríos-Pulgarín et al., 2016; Walteros y Castaño, 2020).
La abundancia y diversidad de los Colectores-Recolectores y de los Fragmentadores, por ejemplo, se ven afectadas por la disponibilidad de material vegetal, el cual tiende a ser más abundante en épocas de bajo caudal (Mozzaquattro et al., 2020). Además, en estas épocas la velocidad de la corriente es menor y se presenta un aumento en la temperatura del agua, una mayor concentración de oxígeno disuelto y el posible incremento en el dióxido de carbono proveniente de la descomposición del material vegetal, factores que filtran la presencia de las especies de acuerdo con sus adaptaciones. Por este motivo, los estudios futuros podrían ahondar en otros aspectos auto-ecológicos, diferentes a los grupos tróficos, para entender cómo la variabilidad hidrológica incide sobre las comunidades de las llanuras aluviales del Orinoco y la Amazonía.
Se han observado cambios en la estructura trófica de los macroinvertebrados a una escala temporal en algunos estudios de ecosistemas colombianos, como el de Rodríguez-Barrios et al. (2011), quienes hallaron que en temporadas de baja precipitación aumenta la abundancia general de todos los organismos, con la predominancia de los Colectores-Recolectores y los Depredadores. En su estudio, Rivera-Usme et al. (2013) establecen que la variación de la estructura trófica puede verse influenciada por cambios hidrológicos, pues en periodos de baja precipitación y de altas temperaturas hay una alta proliferación de algas, que, a su vez, generan un aumento en la abundancia de organismos Colectores-Raspadores. Al comparar los resultados de ambos estudios, se observa que existe una asociación entre la variación ambiental en el tiempo con la diversidad (H') y con la abundancia de los distintos grupos tróficos, pues en meses de bajas precipitaciones aumenta la cantidad de Colectores-Raspadores y de Colectores-Recolectores debido al aumento de la oferta de productores primarios y de materia orgánica en época de baja precipitación (Castro-Rebolledo et al. 2014).
La variación espacial de la estructura trófica no fue tan notable como la variación temporal, entre los tres arroyos muestreados. De acuerdo con lo que han planteado otros autores, esto se ajusta a la idea de que la temporalidad en las condiciones ambientales promueve la capacidad de dispersión de las especies y una recolonización de éstas (Jocque et al., 2010; Tonkin et al., 2018). Además, en algunos estudios se destaca que la variabilidad espacial puede explicarse por medio de la heterogeneidad de los mesohábitats, pues éstos presentan ciertas condiciones que permiten que las especies accedan a una serie de recursos particulares y encuentren refugio ante los cambios hidrológicos (Ríos-Pulgarín et al., 2016; González-Trujillo y Alonso-Moreno, 2020).
Lo anterior ha permitido asociar la diversidad de taxones con los tipos de mesohábitat y con su distribución en el lecho del río (Córdoba-Ariza et al., 2020). Sin embargo, en este estudio no se tuvieron en cuenta diferentes tipos de sustratos, lo cual explicaría por qué no se hallaron diferencias notables a escala espacial. Cabe resaltar que para estas mesoescalas espaciales, algunos factores como la cobertura vegetal pueden ser determinantes para la comunidad, pues pueden influir en la dieta de los grupos tróficos, tales como los fragmentadores de hojas y los recolectores de materia orgánica particulada (LeRoy y Marks, 2006; Walteros y Castaño, 2020).
La brecha entre la falta de información sobre la estructura trófica en ríos de las zonas tropicales es cada vez menor. No obstante, aún existen trabajos que utilizan literatura secundaria de zonas templadas para la clasificación de los grupos tróficos de macroinvertebrados. Lo anterior resulta problemático, pues en distintos estudios se ha podido comprobar que la dieta de las especies puede variar dependiendo de las condiciones de su hábitat y de los recursos disponibles (Díaz-Villanueva y Albariño, 1999; Tomanova et al., 2006). Este estudio no es la excepción, ya que cuando se comparan los grupos tróficos descritos en otros estudios con los de este trabajo, se observa que existen diferencias en su categorización. Baetodes, por ejemplo, es clasificado como Raspador por Merrit et al. (2008), mientras que en los trabajos de Tomanova et al. (2006), Chará-Serna et al. (2010) y en este estudio, se estableció que su comportamiento trófico se asemeja más al de un Colector-Recolector. Además de la alta plasticidad trófica de las especies de macroinvertebrados tropicales, de deben tener en cuenta las limitaciones relacionadas con el nivel taxonómico al cual se hacen algunas clasificaciones.
CONCLUSIONES
Es importante seguir realizando estudios que contemplen la determinación de los grupos tróficos de macroinvertebrados neotropicales, ya que el basarse en la bibliografía para la clasificación de estos organismos puede generar clasificaciones insuficientes o incorrectas. Además, los análisis directos de los hábitos tróficos arrojan información importante, pues los organismos son en su mayoría generalistas y su dieta puede variar de acuerdo a los recursos disponibles en el ecosistema. Por otra parte, es importante que esta clasificación de los grupos tróficos se haga al menos a nivel de género, ya que dentro de una familia determinada, pueden presentarse muchos tipos de dietas.

