










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín de Investigaciones Marinas y Costeras - INVEMAR
Print version ISSN 0122-9761
Bol. Invest. Mar. Cost. vol.23 no.1 Santa Marta Jan./Dec. 1994
FLORECIMIENTO DE MICROALGAS RELACIONADO CON MORTANDAD MASIVA DE PECES EN EL COMPLEJO LAGUNAR CIÉNAGA GRANDE DE SANTA MARTA, CARIBE COLOMBIANO
José Ernesto Mancera P. y Luís Alfonso Vidal V.
INVEMAR, Apartado 1016, Santa Marta, Colombia.
RESUMEN
Entre los meses de julio y agosto de 1994 un fenómeno discontinuo de muerte masiva de peces se registró en aguas del complejo lagunar de la Ciénaga Grande de Santa Marta, Colombia. El primer evento se presentó del 15 al 19 de julio, el segundo alrededor del día 20 de agosto y el tercero entre el 26 y 31 del mismo mes. La presencia en altas concentraciones de la cianofita filamentosa Cf. Anabaenopsis sp., registrada en la literatura como altamente tóxica, podría estar relacionada con las causas del primer evento. Un incremento en las concentraciones de fósforo, y un posterior florecimiento de nano y picoplancton, especialmente cianofíceas y algas flageladas, explicaría en parte las condiciones hipóxicas y anóxicas observadas en el tercer evento, las cuales aparentemente pudieron causar la muerte masiva de peces. Sin embargo, no se descartan otras causas como la liberación de gases tóxicos desde el sedimento y la anoxia debida a la actividad bacteriana. Si bien la biomasa de peces muertos no pudo ser cuantificada, estimativos obtenidos a partir de la información suministrada por pescadores, arrojan para el caso de los chivos (Ariidae) valores que superan las 20 toneladas. Al parecer se trata de uno de los fenómenos de este tipo más grandes de los últimos años.
ABSTRACT
A discontinuous phenomenon of massive fish mortality occurred in the estuarine lagoonal complex of the Ciénaga Grande de Santa Marta, Colombia, in July and August 1994. The first event occurred from the 15th to the 19th of July, the second took place one month later around the 20th of August, and the last one was evident from the 26th to the 31st of the same month. High concentrations of Cf. Anabaenopsis sp., a filamentous cyanobacteria, reported in the literature as highly toxic, may have been related to the causes of the first event. An increase in phosphorus concentrations resulting in massive blooms of cyanobacteria and flagellate algae of nano and picoplankton could explain the hypoxic and anoxic conditions associated to the third event of massive fish mortality. However, other possible causes, such as toxic gas liberation from the sediment and anoxia from bacterial activity cannot be roled out. The dead fish biomass was not quantified, but information obtained from fishermen suggests for sea catfish (Ariidae) values well over 20 tons. The massive death phenomenon here reported seems to be the strongest in the last years.
INTRODUCCIÓN
En la costa norte colombiana, departamento del Magdalena, se encuentra un complejo ecosistema conformado por numerosos canales, pantanos de manglar y lagunas, dentro de las cuales la de mayor área es la Ciénaga Grande de Santa Marta (CGSM) (figura 1), considerada como la más grande de Colombia (450 Km2) y una de las más importantes de la cuenca caribeña (Botero y Botero, 1989). En la actualidad el complejo lagunar afronta una grave problemática ambiental, que se ve reflejada en el estado de deterioro de sus recursos naturales y en el empobrecimiento de sus habitantes (Mancera y Botero, 1993). Como una de las actividades contempladas en el proceso de recuperación del ecosistema, el proyecto Colombo-Alemán "Rehabilitación de la Ciénaga Grande de Santa Marta", PRO-CIENAGA, pretende reestablecer las condiciones hidráulicas, reabriendo canales que transportan agua dulce desde el Río Magdalena hasta la CGSM.

Fenómenos de mortandad masiva de organismos han sido registrados anteriormente en la CGSM y son conocidos por los habitantes de la región con el nombre de "tabaquillo" Mercado (1971) describe el "afloramiento" (sic) de la timonera, Anabaena circinalis, y la relaciona con la muerte por intoxicación de peces, perros, cerdos y gallinas. Años después Bula (1985) identificó la cianofícea Nostoc commune (= Anabaena flos-aquae) como responsable de otro fenómeno de mortandad masiva de organismos. Hasta el momento la única causa señalada como responsable del fenómeno fue la ingestión de algas tóxicas, sin que hubieran sido consideradas y documentadas otras posibles causas.
La presencia continuada de investigadores en el área de la CGSM permitió detectar a tiempo el fenómeno de mortandad masiva de peces ocurrido entre el 15 de julio y 31 de agosto/94, lográndose observaciones detalladas que condujeron a una explicación más amplia de las causas del fenómeno.
MATERIALES Y MÉTODOS
El fenómeno de mortandad masiva de peces se presentó en el costado oriental de la Ciénaga de Pajaral, Caño Grande y costado occidental de la CGSM, durante tres pulsos o eventos. El primero ocurrió entre el 15 y 19 de julio/94, el segundo alrededor del 20 de agosto y el tercero del 26 al 31 del mismo mes. Para establecer las causas del fenómeno, se realizaron salidas de campo los días 18 y 22 de julio y 30 y 31 de agosto. Para el segundo evento no se realizó visita al área del problema, sin embargo, versiones de pescadores permitieron acopiar información. Los días 18 y 22 de julio se tomaron muestras puntuales de agua superficial para el análisis de fitoplancton. El 22 de julio, utilizando sondas WTW previamente calibradas, se realizaron medidas in situ de temperatura, pH y salinidad en cuatro estaciones (figura 1); esta última variable se midió y expresó empleando la escala de salinidad práctica, la cual no tiene ni dimensiones ni unidades (UNESCO, 1985); además, fueron tomadas muestras de agua para determinar la concentración de clorofila a en laboratorio, siguiendo el método de Lorenzen (1967).
Los resultados obtenidos y las hipótesis planteadas durante el primer evento, permitieron preparar un diseño de muestreo más completo para el tercero, determinándose además de las variables anteriores, la profundidad y transparencia de la columna de agua (disco de Secchi), las concentraciones de las clorofilas b y c (APHA, 1975), el índice de pigmentos rojos/verdes, los niveles de fósforo total, ortofosfatos y oxígeno disuelto. Todas estas variables se midieron durante un ciclo diario (agosto 30-31/94), en dos estaciones de muestreo ubicadas en Caño Grande (parte central) y El Morro (figura 1). Los valores de oxígeno disuelto fueron determinados in situ, mediante una sonda WTW OXI-92, a dos niveles de la columna de agua (superficie y fondo localizado a 0.6 m) en el Morro, y a cuatro profundidades (0, 2, 4 y 6 metros) en Caño Grande. Los ortofosfatos y el fósforo total, se determinaron por duplicado en espectrofotómetro, siguiendo los métodos descritos en Grasshoff et al (1983). Relacionando la profundidad y la transparencia del agua, se calculó el porcentaje de penetración de luz.
Los muestreos de fitoplancton, para el tercer evento, se realizaron empleando una botella tipo Ruttner de 11 de capacidad. La identificación de fitoplancton para el primero y tercer evento se llevó a cabo utilizando un microscopio óptico, utilizando las descripciones de Vidal (1993); los conteos se realizaron sobre un portaobjeto con cubreobjeto mediante barridos en zig-zag, previa calibración del volumen de la alícuota de agua.
La biomasa de peces muertos en cada uno de los tres eventos no pudo ser cuantificada, por lo que se recurrió a una estimación cualitativa de los peces que se encontraron flotando y de los capturados por los pescadores.
RESULTADOS
El recuento de las características más importantes de cada uno de los tres eventos del fenómeno, muestra que el número de especies ícticas afectadas aumentó del primer evento al tercero, y que tanto boconas (Engraulididae) como chivos (Ariidae) fueron las especies más afectadas en los dos casos (tabla 1). Estimativos obtenidos a partir de las capturas de los pescadores durante el tercer evento arrojan, para el caso de los chivos, valores que superan las 20 toneladas.

La mayoría de variables medidas en el primer evento mostraron valores que se encuentran dentro de los comúnmente observados para estas aguas (tabla 2). Se destacan las concentraciones de clorofila a, por ser más elevadas que las normalmente observadas, indicando esto una alta concentración de fitoplancton. Estos resultados concuerdan con los obtenidos en el análisis de la composición del fitoplancton de dos muestras de agua tomadas en Caño Grande (tabla 3). El conteo de algas durante el primer evento fue semi-cuantitativo, pero permitió determinar que Cf Anabaenopsis sp. (figura 2), una cianofita filamentosa, fue la forma predominante.

En la tabla 4 se presentan las concentraciones de fitopigmentos (clorofilas a, b y c), índice de pigmentos, fósforo total, ortofosfatos, transparencia, salinidad y temperatura durante un ciclo diario (agosto 30-31/94) para las dos estaciones de muestreo. Se destacan los valores de fósforo tanto inorgánico como total, por ser muy altos, superando tres veces los valores normalmente obtenidos en períodos diferentes para los mismos sitios (Base de datos INVEMAR, sin publicar).


Estos valores altos de fósforo parecen coincidir con la gran abundancia fitoplanctónica, que por su tamaño corresponde a las fracciones del pico y nanoplancton principalmente (tablas 5 y 6). Las observaciones al microscopio y los análisis de los fitopigmentos clorofilas b y c e índice de pigmentos muestran que las formas predominantes de microalgas correspondieron a cianofíceas (tablas 4 y 6). Así mismo, los valores obtenidos para clorofila a, resultaron ser muy elevados (95 a 137 μg/l), comparados con los normalmente observados páralos mismos sitios (11 a 71 μg/l, Base de datos de INVEMAR, sin publicar). La gran concentración de plancton fue la responsable de los bajos valores de transparencia, que para Caño Grande estuvieron entre 0.25 y 0.30 m, y para el Morro entre 0.12 y 0.15 m.

Si bien las concentraciones de clorofila y fósforo no mostraron variaciones importantes a lo largo del día en ninguna de las estaciones de muestreo, la concentración de oxígeno disuelto para las estaciones de Caño Grande y El Morro fue muy variable, descendiendo a concentraciones de hipoxia (menores a 2 mg/l) y anoxia (cercanas a cero) (figura 3) durante las horas de la noche y la madrugada.
En la estación del Morro, los valores de oxígeno disminuyeron drásticamente a valores inferiores a 2 mg/l, en ambos niveles de profundidad, entre las 18:00 y 21:00 horas, permaneciendo la concentración de oxígeno por debajo de 1 mg/l entre las 24:00 y las 10:00 del día siguiente, e incrementándose luego a valores superiores a 3 mg/l hacia el mediodía. Estos resultados muestran que los valores de hipoxia-anoxia se prolongaron durante 13 horas.
En la estación de Caño Grande los valores de oxígeno se incrementaron para todas las profundidades entre las 13:00 y las 17:00 horas, disminuyendo luego fuertemente, principalmente a 4 y 6 m de profundidad, entre las 17:00 y 23:00 horas a concentraciones por debajo de 1 mg/l; más tarde, a las 02:00 horas toda la columna de agua presentó dicha situación, la cual se prolongó por lo menos hasta las 11:00 horas del día siguiente en las profundidades de 4 y 6 m, siendo ligeramente superior a la profundidad de 2 m y de 2.6 mg/l en superficie.

DISCUSIÓN
Aunque la magnitud del fenómeno no pudo ser cuantificada, la presencia confirmada (por varias fuentes de pescadores y personal del INVEMAR) de peces muertos durante tres eventos diferentes en un período de 45 días, permite presumir que se trata de uno de los fenómenos de éste tipo más grande de los últimos años.
Al parecer el fenómeno es multicausal. El primer evento estuvo directamente asociado con el florecimiento de la microalga Cf. Anabaenopsis sp., una cianofita ó alga azul-verdosa, cuyos filamentos miden 3.5 μm de diámetro, y la cual ha sido registrada, con el nombre de Nostoc commune, como altamente tóxica (Bula, 1985). Según algunos autores el incremento relativo de un determinado elemento puede determinar un florecimiento microalgal (Ryther y Dunstan, 1971; Cloern, 1982; Margalef, 1983).
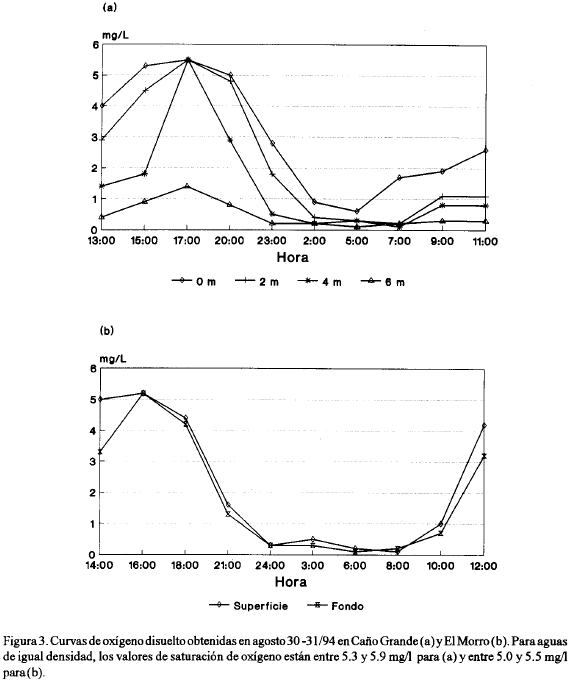
Para el caso de las cianofitas, incrementos de fósforo, al parecer, favorecen su crecimiento (Howarth et al, 1988; Cuker et al, 1990; Konoen, 1992 en Nehring, 1993). De otra parte, en sistemas lagunares costeros, el fósforo en formas solubles, es principalmente liberado de los sedimentos cuando estos están en condiciones anaeróbicas (Day et al, 1989; Nixon, 1982). En presencia de oxígeno, el fósforo permanece de manera insoluble en los sedimentos, asociado a un complejo de fósforo y hierro (P-Fe (III)), pudiendo ser reducido luego, en ausencia de oxígeno, a la forma ferrosa la cual es soluble (Reid y Wood, 1976). En períodos de calma debido a la ausencia prolongada de vientos y ante condiciones de estratificación salina de la columna de agua, como la observada en el Complejo Pajarales (tabla 4), es de esperar consumo de oxígeno hasta condiciones de hipoxia o anoxia. Si la calma se presenta durante la noche, la anoxia será mayor, como consecuencia de la carencia de producción de oxígeno al verse interrumpido el proceso fotosintético. Con el crecimiento de la biomasa fitoplanctónica, es de esperar aumento de oxígeno disuelto en el agua durante el día, pero elevado consumo del mismo durante la noche, lo cual se pudo comprobar mediante las curvas diarias de oxígeno disuelto (figura 3). Aunque las condiciones de vientos se reestablezcan, el proceso respiratorio de una gran cantidad de organismos, puede durante la noche, conducir a condiciones de hipoxia y anoxia. En condiciones normales, la concentración de oxígeno disuelto puede bajar durante la noche; sin embargo, los valores permanecen por arriba de 3 mg/l, como lo muestran las curvas de oxígeno (figura 4), durante ciclos de 48 horas en el centro de la CGSM (Mancera y Botero, 1990, datos no publicados). En este caso no se presentan niveles de hipoxia debido a la acción de los vientos, los cuales mantienen oxigenada y mezclada la columna de agua, aún cuando la cantidad de organismos sea relativamente grande (no tanto como en una discoloración) y por lo tanto el consumo de oxígeno durante la noche también lo sea.

Aumentos en las concentraciones de fósforo también se pueden producir con el aporte de nutrientes por escorrentía (Nichols et al, 1986; Valiela et al., 1992); en este caso, los nutrientes pueden ingresar al Complejo Pajarales por caños, y como producto del lavado de pantanos de manglar, con poca vegetación o sin ella. La entrada de nutrientes favorece el florecimiento de cianofitas, que a la larga son las posibles responsables del fenómeno de mortandad de peces. Fenómenos de este tipo se han presentado antes en el sistema, generalmente durante las dos época lluviosas que se presentan cada año en la región (mayo - junio y septiembre - noviembre).
La fertilización de las aguas, bien por liberación de nutrientes del sedimento, bien por aporte de otros sistemas, permite el florecimiento de especies fitoplanctónicas, principalmente de tamaño reducido (menor de 10 m), las cuales presentan altas tasas de renovación (Margalef, 1986). Estas algas pueden ser tóxicas como en el caso de Cf. Anabaenopsis sp.; o pueden no serlo. En el primer caso, organismos como peces pueden morir al ingerirlos. En ambos casos la probabilidad de generar hipoxias es muy grande, especialmente durante la noche, cuando el proceso fotosintético cesa y solamente hay consumo de oxígeno por respiración. Esta hipótesis ha sido comprobada por los niveles de oxígeno encontrados, y por la observación de peces moribundos en horas de la madrugada. El efecto de falta de oxígeno puede ser aumentado si hay obstrucción branquial por parte de la biomasa fitoplanctónica, situación que pudo suceder, pues en la autopsia practicada a varios individuos de C. spixi, se observó gran cantidad de cianoflceas adheridas a las branquias. En la medida en que se agotan los nutrientes disponibles, la población fitoplanctónica del florecimiento empieza a morir, constituyéndose la biomasa muerta en objeto de descomposición, proceso que demanda consumo de oxígeno, explicando los valores bajos de oxígeno, aún durante el día (figura 3).
CONCLUSIONES
La fertilización adicional en un sistema eutroficado como la CGSM (Hernández y Gocke, 1989), condiciona el florecimiento microalgal, el cual puede ser tóxico y/o generar, por diferentes vías, problemas de hipoxia-anoxia en la columna de agua y mortandad de organismos aerobios obligatorios. Aunque las observaciones y mediciones realizadas apoyan en gran medida las hipótesis planteadas, no se descartan otras causas de mortalidad como la liberación de gases tóxicos (H2S y CH4) desde los sedimentos anóxicos, y anoxia debida a actividad bacteriana.
El problema de mortandad masiva de peces en CGSM es un fenómeno recurrente, al parecer incrementado por la situación de deterioro del sistema; por lo tanto, no existe solución a corto plazo. La situación tiende a agravarse si se considera que el fenómeno actual afectó gran cantidad de peces claves en la trama trófica (engraúlidos), los cuales transfieren energía desde los productores primarios a los carnívoros (Cervigón, 1980; Santos-Martínez, 1989; Duque, 1993; Santos-Martínez y Arboleda, 1993), y que en el futuro cercano pueden ocurrir más eventos.
Esta situación demanda afanosamente el proceso de recuperación, reabriendo los flujos de agua Río Magdalena - CGSM - Mar Caribe. Aunque en principio, podría esperarse un impacto similar de mortandad de peces, es posible que a largo plazo el sistema se recupere, disminuyendo sus niveles de eutroficación y mejorando la calidad de los productores primarios (plancton, manglar). Es necesario entonces, desarrollar un monitoreo permanente del ecosistema, con especial énfasis en los sitios aledaños a las obras de reapertura de canales de agua dulce provenientes del Río Magdalena, observando curvas diarias de oxígeno, liberación de gases tóxicos, carga de nutrientes, desarrollo y composición del plancton, toxicidad de microalgas en peces, así como también, dinámica de vientos y lluvias.
AGRADECIMIENTOS
Los autores agradecen a COLCIENCIAS, al Instituto de Investigaciones Marinas y Costeras "José Benito Vives D'Andreis" INVEMAR y al proyecto de Cooperación Técnica Colombo-Alemán "Rehabilitación de la Ciénaga Grande de Santa Marta", PRO-CIENAGA, por el apoyo financiero. Al Ing. Efraín Viloria, al Lic. Walberto Troncoso y la M.Sc. Adriana Santos-Martínez integrantes del programa de investigación en Lagunas Costeras del INVEMAR, por su trabajo de campo y laboratorio. A la Dra. Leonor Botero por la revisión del abstract. Así mismo a los habitantes de los pueblos de Buenavista y Nueva Venecia por su valiosa información.
BIBLIOGRAFÍA
1 APHA. 1975. Standard methods for examination of water and waste water, 14th ed. APHA, AWWA, WPCF, Washington, D.C., USA, 1193 p. [ Links ]
2 Botero, J. E. y L. Botero. 1989. Problemática del sistema Ciénaga Grande de Santa Marta, Delta Exterior del Río Magdalena. En: Colombia y el agua: tres aspectos. Bogotá. Fescol 5:11-28. [ Links ]
3 Bula, G. 1985. Florecimientos nocivos de algas verde-azules en dos lagunas del departamento del Magdalena. Ingen. Pesq., 5 (1,2):89-99. [ Links ]
4 Cervigón, F. 1980. Ictiología Marina. Vol. I. Ed. Arte, Caracas, 358 p. [ Links ]
5 Cloern, J.E. 1982. Does the benthos control phytoplankton biomass in south San Francisco Bay? Mar. Ecol. Prog. Ser. 9:191-202. [ Links ]
6 Cuker, B.E.; P.T. Gama y J.M. Burkholder. 1990. Type of suspended clay influences lake productivity and phytoplankton community response to phosphorus loading. Limnol.Oceanogr, 35 (4):830-839. [ Links ]
7 Day, J.; C. Hall; M. Kemp y A. Yáñez-Arancibia. 1989. Estuarine Ecology. John Wiley and Sons, Inc. New York, USA, 558 p. [ Links ]
8 Duqe, G. 1993. Ecología trófica y aspectos reproductivos de las especies del género Oligoplites (Pisces: Carangidae) de la Ciénaga Grande de Santa Marta, Caribe colombiano. Tesis Biol. Mar., Univ. Valle, Cali, 100 p. [ Links ]
9 Grasshoff, K.; M. Ehrhardt y K. Kremling (Eds). 1983. Methods of seawater analysis.Verlag Chemie, Weinheim, Alemania, 142 p. [ Links ]
10 Hernández, CA. 1988. Características físicas, químicas y biológicas de la Ciénaga Grande de Santa Marta, Colombia. Inf. Proy., II parte, INVEMAR, Santa Marta, 73 p. [ Links ]
11 Hernández, CA. y K. Gocke. 1988. Productividad primaria en la Ciénaga Grande de Santa Marta,Colombia. An. Inst. Invest. Mar. Punta Betín. 19-20:101-119. [ Links ]
12 Howarth, R.W.; R. Marino y J.J. Cole. 1988. Nitrogen fixation in fresh water, estuarine and marine ecosystems. 2. Biogeochemical controls. Limnol. Oceanogr., 33:688-701. [ Links ]
13 Lorenzen. C.J. 1967. Determination of chlorophyll and pheopigments: spectophotometric equations. Limnol. Oceanogr., 12:343-346. [ Links ]
14 Mancera, J.E. y L. Botero. 1993. Estudio Ecológico de la Ciénaga Grande de Santa Marta, Delta Exterior del Río Magdalena. III etapa. Inf. Proy., INVEMAR, Santa Marta, 128 p. [ Links ]
15 Margalef, R. 1986. Ecología. Ed. Omega S.A., Barcelona, 951 p. [ Links ]
16 Margalef, R. 1983. Limnología. Ed. Omega S.A., Barcelona, 1010 p. [ Links ]
17 Mercado, J.E. 1971. Inventario preliminar de la fauna y flora de la Ciénaga Grande de Santa Marta. Proy. Desar. Pesca Marit. Colombia (INDERENA - PNUD - FAO), CIP, Cartagena, 8 p. [ Links ]
18 Nehring, S. 1993. Mortality of dogs associated with a mass development of Nodularia spumigena (Cyanophyceae) in a brackish lake at the German North Sea coast. J. Plank. Res., 15(7):867-872. [ Links ]
19 Nichols, F.H.; J.E. Cloern; S.N. Luoma y D.H. Peterson. 1986. The modification of an estuary. Science, 231:567-573. [ Links ]
20 Nixon, S.W. 1982. Nutrient dynamics, primary production and fisheries yields of lagoons. Oceanol. Acta, 152:357-371. [ Links ]
21 Reid, G. K. y R. D. Wood. 1976. Ecology of inland waters and estuaries. D. Van Nostrand. New York, USA, 485 p. [ Links ]
22 Ryther, J.H. y W.M. Dunstan. 1971. Nitrogen, phosphorus, and eutrophication in the coastal marine environment. Science, 171:1008-1013. [ Links ]
23 Santos-Martínez, A. 1989. Estudio biológico y ecológico de la ictiofauna de la Ciénaga Grande de Santa Marta, Caribe colombiano. Tesis. M.Sc. Biol. Mar., Univ. Nacional de Colombia, Bogotá, 178 p. [ Links ]
24 Santos-Martínez, A. y S. Arboleda. 1993. Aspectos biológicos y ecológicos del Macabí Elops saurus Linnaeus (Pisces: Elopidae) en la Ciénaga Grande de Santa Marta y costa adyacente, Caribe colombiano. An. Inst. Invest. Mar. Punta Betín, 22:77-96. [ Links ]
25 UNESCO. 1985. The international system of units (SI) in oceanography, UNESCO Technical Papers No. 45, IAPSO Pub. Sci. No. 32, Paris, France, 124 p. [ Links ]
26 Valiela, I.; K. Foreman; M. LaMontagne; D. Hersen; J. Costa; P. Peckol; B. DeMeo-Anderson D'Avanzo; M. Babione; C. Sham; J. Brawley y K. Lajtha. 1992. Couplings of watersheds and coastal waters: Sources and consequences of nutrient enrichment in Waquoit Bay, Massachusetts. Estuaries, 15(4):443-457. [ Links ]
27 Vidal, L.A. 1993. Estudio del fitoplancton en el sistema lagunar estuarino tropical Ciénaga Grande de Santa Marta, Colombia, durante el año 1987. Tesis M.Sc. Biol. Mar., Univ. Nacional de Colombia, Bogotá, 208 p + 62 láminas. [ Links ]
