











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
El tomate de mesa (Solanum lycopersicum) es la fruta fresca y procesada más importante en el mundo y es, después de la papa, el vegetal que más se consume (Nangare et al. 2016; Heuvelink, 2018), así como es una fuente importante de antioxidantes, como licopeno, fenólico y vitamina C, en la dieta humana (Toor et al. 2006). La producción de tomate, a nivel mundial, para el 2022, fue de aproximadamente 255 millones de toneladas, en alrededor de 6 millones de hectáreas (FAOSTAT, 2024). En Colombia, el área sembrada para el 2021 fue de 10 mil hectáreas, con una producción anual de 272 mil toneladas y un rendimiento promedio de 26,98 t ha-1 (AGRONET, 2024).
Si bien el tomate es un producto que ayuda a garantizar la seguridad alimentaria, este cultivo se caracteriza por sus altos requerimientos hídricos. Debido a la creciente amenaza de escasez de agua que afecta al mucho en la actualidad (Mekonnen & Hoekstra, 2016) es importante desarrollar metodologías que permitan optimizar el uso del agua, manteniendo los rendimientos y la calidad de la fruta. El agua es el factor mas importante para maximizar la producción y mejorar la calidad frutos en este cultivo; su eficiencia consiste en ofrecer la cantidad adecuada en el momento adecuado requerido por la planta.
Así, los principales factores a controlar en la producción de cultivos son: el aporte hídrico eficiente, el aporte nutricional y la evaluación de variedades mejoradas. En este sentido, la estimación y el ajuste de los coeficientes de cultivo (Kc) eficiente con base al genotipo y al ambiente, permite optimizar la programación de riego y la eficiencia del uso de agua, lo que permite a los cultivos expresar todo su potencial genético de producción (Allen et al. 2006; Jaramillo et al. 2007; Mekonnen & Hoekstra, 2016), evitando la sobreexplotación del recurso hídrico.
Por lo anterior, el objetivo de la presente investigación fue evaluar diferentes coeficientes de cultivo (Kc) en dos fases de cultivo: establecimiento y floración - llenado, en dos híbridos de tomate Pietro F1 y Mateus (7370).
MATERIALES Y MÉTODOS
Este estudio se llevó a cabo en el Centro Internacional de Producción Limpia Lope, perteneciente al Servicio Nacional de Aprendizaje - SENA, del municipio de Pasto, departamento de Nariño, Colombia, situado a N 1°12’13’’ y O 77°15’23’’, con temperatura promedio de 12 °C y evapotranspiración promedio de 2,5 mm día-1.
El diseño experimental empleado fue bloques completos al azar, con arreglo en franjas divididas, donde el primer factor está conformado por tres niveles de láminas de agua, calculados a partir de los coeficientes, donde Kc1<Kc2<Kc3 y el segundo factor estuvo constituido por dos niveles, siendo estos los híbridos de tomate Pietro F1 y Mateus (7370) y tres repeticiones, para un total de 18 unidades experimentales (Lagos & Criollo, 2019).
La medición de variables se realizó durante la fase de establecimiento y de producción; de este modo, las variables de altura de planta y diámetro de tallo de la planta se midieron entre los 0 a 70 despues del transplante DDT y 71 a 149 DDT; por otra parte, el número de flores, frutos y racimos se evaluaron entre los días 71 a 149 DDT. Finalmente, las variables peso unidad y diámetro de fruto, peso de frutos por planta y rendimiento se evaluaron durante los días 104 a 153 DDT (Tabla 1).
Los métodos de toma de registros se hicieron con base a las metodologías expuestas por Ramírez-Vargas & Nienhuis (2012), para desarrollo fenológico del tomate.
Con los datos obtenidos, se realizó un análisis de varianza, para determinar diferencias estadísticas con un grado de confiabilidad del 95 %, con la prueba de comparación de medias LSD Fisher (p<0,05). Los análisis se realizaron en el software Infostat v.2019 y las gráficas de interacción de factores, cuando hubo presencia de esta, se realizaron en R v. 2021.09.
RESULTADOS Y DISCUSIÓN
Para la fase de establecimiento de 0 a 70 DDT, las variables de altura de planta y diámetro de tallo no presentaron diferencias estadísticas, indicando que durante esta fase no hay una respuesta diferencial de los híbridos a los Kc evaluados (0,1; 0,2 y 0,3), resultando ser iguales estadísticamente; asimismo, los resultados indican que durante esta fase, la eficiencia hídrica de los dos híbridos es igual. Resultados similares encontraron Barraza et al. (2004), quienes al evaluar altura de planta entre los 15 y 60 DDT, no encontraron diferencias estadísticas y atribuyen este hecho a que en esta fase aún no existe competencias entre plantas, afirmando que los genotipos vegetales responden morfológica y fisiológicamente frente a la densidad de población.
Para la fase de producción evaluada entre los 71 a 153 DDT, la única variable que presentó diferencias estadísticas para los híbridos fue el número de flores, siendo el híbrido Pietro superior, con una media de 8,72 flores planta1, a diferencia de Mateus, que presentó una media de 8,16 flores planta.1. Esto se debe, posiblemente, a una respuesta diferencial de los híbridos a la fertilización, como lo expresa Cadahía (2005), quien afirma que la fertirrigación influye drásticamente sobre la producción si se tiene en cuenta que algunos híbridos son nutricionalmente más eficientes que otros. Por otra parte, el hecho de encontrar estas diferencias estadisticas se deba, posiblemente, a una mejor adaptación del híbrido Pietro a las condiciones edafoclimaticas del invernadero donde se realizó el estudio, ya que, como lo afirman Higashide & Heuvelink (2009), condiciones específicas, como altas temperaturas, humedad y radiación solar son determinantes para que los diferentes materiales expresen sus capacidades; asimismo, estas condiciones específicas podrían estar relacionadas con la reducción de abortos florales, ya que, como lo afirma Guzmán (1991), este fenómeno se causa por un mejor acondicionamiento de las plantas a unas determinadas condiciones ambientales, entonces, se podría inferir que el híbrido Pietro, se adapta mejor a condiciones de estrés por sequía, teniendo en cuenta que esta es una de las causas para producir aborto floral (Zegbe et al. 2006).
En la fase de producción evaluada entre los 71 a 153 DDT, el análisis de varianza mostró diferencias estadísticas para el efecto causado por las láminas de riego, para la variable peso por unidad de fruto e, igualmente, se encontró el efecto de la interacción de los factores láminas por híbridos en las variables número de frutos, peso de frutos por planta y rendimiento.
La prueba de comparación de medias LSD Fisher mostró que la lámina con una media más alta para la variable de peso de fruto fue la correspondiente al Kc de 1,1, con una media de 125,03g (Tabla 1); esto se debe, a que un riego deficitario reduce el tamaño de la fruta, el rendimiento comercial durante la floración y el desarrollo de la fruta y las etapas de maduración de esta (Pulupol et al. 1996). Resultados similares encontraron Cui et al. (2020), quienes reportan una disminución en el peso y el volumen de los frutos, cuando el suministro hídrico del cultivo fue limitado encontrando pesos de 108,17 g, cuando el suministro de agua fue de 394 mm y una media en peso superior de fruto de 120,03 g, cuando el suministro de agua fue de 490 mm.
Tabla 1 Comparación de medias de las variables número de frutos por planta, peso de fruto, producción por planta, diámetro ecuatorial del fruto y rendimiento.
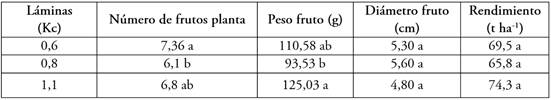
Medias seguidas de letras diferentes en la columna, presentan diferencias estadísticas entre sí, según la prueba de LSD Fisher (P<0,05). Medias con igual índice en las letras son semejantes estadísticamente, según la prueba de LSD Fisher (P<0,05).
Por otra parte, la gráfica de interacción mostró para la variable número de frutos por planta que el híbrido Pietro, con una lámina Kc de 1,1, presentó una media superior, con 9,1 frutos planta-1, seguida por el híbrido Mateus con la lámina Kc de 0,6, con una media de 8,1 frutos planta-1 (Figura 1a), indicando que los híbridos presentan un comportamiento diferencial a las láminas de riego evaluadas. Las diferencias en el número de frutos causadas por los efectos de la interacción lámina e híbrido encontradas, se debe, posiblemente, a que las capacidades productivas de los híbridos pueden aumentar o disminuir, dependiendo del genotipo (Florido & Bao, 2014; Shao et al. 2015).
En cuanto al rendimiento, la gráfica de interacción para la variable rendimiento (Figura 1b) muestra que la interacción con un rendimiento más alto corresponde al híbrido Pietro con una lámina Kc de 1.1, presentando una media de 101 t ha-1, seguida del híbrido Mateus con una lámina Kc de 0.6, con un rendimiento promedio de 79,5 t ha-1, mostrando una relación directa e inversamente proporcional entre el rendimiento y la cantidad de agua suministrada para los híbridos Pietro y Mateus, respectivamente. El hecho de encontrar estos resultados con el híbrido Mateus, se debe, posiblemente, a que son cualidades propias del híbrido que se expresaron genotípicamente, al estar en condiciones específicas (Florido y Bao, 2014), en este caso, la lámina de riego. Lo anterior indica que, posiblemente, el híbrido Mateus, tenga un potencial para soportar condiciones de estrés hídrico o riego deficitario, resultando ser más eficiente al momento de utilizar el recurso hídrico, sin perder su potencial productivo.
Por otra parte, el comportamiento mostrado por el híbrido Pietro resultan ser más comunes, teniendo en cuenta que un aporte hídrico eficiente se ve reflejado en mejores rendimientos, como en el estudio realizado por Biswas et al. (2015), donde se reporta que los mejores rendimientos se obtienen con una lámina basada 100 % en el requerimiento hídrico del cultivo con 63,84 t ha-1 y disminuye cuando aplica el 75 y 50 % del requerimiento, presentando medias de producción de 54,38 y 50,60 t ha-1, respectivamente. Así, estos estudios permiten soportar los datos obtenidos en esta investigación, respecto al híbrido Pietro.
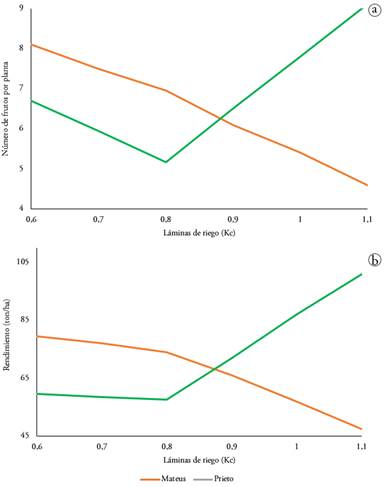
Figura 1 Gráfica de interacción para las variables. a) número de frutos por planta y b) rendimiento.
Finalmente, se concluye que los híbridos evaluados, Pietro y Mateus, presentan un comportamiento diferencial a las distintas láminas de riego evaluadas, siendo el híbrido Pietro un material más exigente en del recurso hídrico, para alcanzar su potencial genético de producción; por el contrario, el híbrido Mateus, se presenta como un material más eficiente para el uso de este recurso, lo que hace que pueda ser catalogado como un material ideal para zonas donde las condiciones ambientales sean adversas y la disponibilidad del recurso sea limitada.

