











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
En Colombia, el Bosque seco Tropical - BsT ha sido expuesto a presiones antrópicas de diferente índole, tales como la ganadería, la expansión agrícola y la minería legal e ilegal, por lo que actualmente se conserva menos del 8% de las coberturas de este ecosistema en el país (Cotler & Ortega-Larrocea, 2006; Ruiz et al., 2013). Por su parte, en el Valle del Cauca el ecosistema de BsT cuenta con aproximadamente el 9.46 % de su cobertura original (Alvarado-Solano & Otero-Ospina, 2015), presentando parches de bosque dispersos, con tamaños reducidos y poca cobertura vegetal. Una de las mayores presiones a la que están expuestos estos bosques son las áreas de cultivo extensivas, las cuales dominan el paisaje vallecaucano (Arcila-Cardona et al., 2012; ). Por estas razones, el BsT se considera como uno de los ecosistemas con mayor grado de amenaza en el planeta, así como uno de los menos estudiados (Pizano & García, 2014). Sin embargo, se ha registrado que dentro de los bosques tropicales se puede encontrar una gran diversidad (; Vargas, 2012), que proporciona beneficios a las comunidades humanas, o lo que se ha establecido como servicios ecosistémicos.
Dentro de la variedad de servicios ecosistémicos asociados a los bosques tropicales se resalta la dispersión de semillas realizada por animales, proceso conocido como zoocoria, el cual es un servicio ecosistémico asociado a las interacciones entre las plantas y animales, donde diferentes grupos de vertebrados, como los mamíferos, intervienen en el movimiento de las semillas permitiendo que las plantas se dispersen y germinen a cierta distancia de los progenitores (Ortiz-Pulido et al., 2000). De esta manera, los bordes de los bosques pueden ampliarse, y así se da el establecimiento de algunos corredores biológicos entre remanentes y un mayor reclutamiento de plántulas al interior del bosque. Dentro de los mamíferos, los murciélagos (Chiroptera) son un grupo clave, ya que es un orden muy diverso, con 217 especies descritas para Colombia (Ramírez-Chaves et al., 2021), contiene especies con hábitos alimenticios contrastantes y pueden ser considerados como indicadores de la perturbación de los ecosistemas (Jones et al., 2005). Particularmente, los murciélagos frugívoros de la familia Phyllostomidae, son considerados importantes para la dispersión de semillas de plantas de los primeros estadíos de sucesión, lo que implica la regeneración natural de los bosques (Muscarella & Fleming, 2007). La dispersión de semillas por parte de este grupo es considerada de alto valor económico para los bosques tropicales, además, es un proceso importante para la estabilidad del BsT, lo que incrementa la importancia de la conservación de los murciélagos en ecosistemas altamente transformados (Aroca et al., 2016).
Adicionalmente, las dinámicas entre animales y plantas son importantes para entender los procesos ecológicos que generan y mantienen la biodiversidad, la estructura y composición, así como, las especies claves para los ecosistemas. El análisis de estas relaciones es posible por medio de interacciones mutualistas, ya que permiten determinar las dinámicas demográficas y la coevolución de las especies en las comunidades (Bascompte et al., 2007). De tal forma, dichas relaciones analizadas a través del tiempo evidencian la acumulación de las interacciones entre los diferentes actores, en distintos niveles y grados de asociación en el tiempo y el espacio (Jordano et al., 2009) y resultan en propiedades distintivas que diferencian las redes mutualistas de otras redes ecológicas (Vázquez et al., 2009). Las denominadas redes de interacción se describen mediante nodos y enlaces, los cuales hacen referencia a los actores relacionados, así como a la fuerza del vínculo entre ellos. La topología de las redes es un reflejo de las dinámicas que ocurren dentro de los ecosistemas, así como de los roles que cumplen las especies en estos ambientes (Bascompte & Jordano, 2007; 2008). Por lo tanto, con el fin de identificar las relaciones entre murciélagos y plantas en el BsT, así como especies clave y las dinámicas que ayudan al mantenimiento del ecosistema, el objetivo del presente estudio fue identificar las interacciones entre murciélagos frugívoros y plantas a través de redes de interacción mutualistas en diez remanentes de BsT en el Valle del Cauca.
Materiales y métodos
Fase de campo
El muestreo se llevó a cabo entre diciembre del 2019 y septiembre del 2021 en diez remanentes de BsT ubicados en la región centro-norte del departamento del Valle del Cauca, Colombia (Anexo 1). La fase de campo contó con un total de ocho noches efectivas de muestreo. Los remanentes se visitaron en temporadas climáticas diferentes cuando fue posible y en cada visita se contó con cuatro noches de muestreo efectivo. Los remanentes de BsT muestreados representan fragmentos de bosque ubicados en áreas protegidas, entre matrices de cultivos, o en predios privados (Figura 1). A todos los remanentes de BsT se les asignó una letra con el fin de codificar los individuos capturados en cada sitio, siendo (A) Parque Natural Regional Mateguadua, (B) Reserva Forestal Protectora Nacional Quebrada La Valenzuela, (C) Hacienda El Volcán, (D) Hacienda La Venta, (E) Predio La Gitana, (F) Reserva Natural Chimbilaco, (G) Parque Natural Regional El Vínculo, (H) Bosque comunitario de Higueroncito, (I) Hacienda El Medio y (J) Hacienda Verdún. Los remanentes abarcan los municipios de Andalucía, Bugalagrande, Guadalajara de Buga, Riofrío, Roldanillo, San Pedro, Tuluá, Yotoco y Zarzal (Figura 1). La altitud de los sitios visitados varió en un rango desde los 940 hasta los 1160 metros sobre el nivel del mar (msnm). El BsT del Valle del Cauca se caracteriza por presentar un régimen climático bimodal, con épocas lluviosas entre abril - mayo y octubre - noviembre, que representa el 70 % de la precipitación anual, y dos temporadas secas entre enero - febrero y julio - agosto (Armbrecht & Ulloa-Chacón, 1999).

Figura 1 Mapa de los remanentes de bosque seco tropical visitados durante el estudio en el centro y norte del departamento del Valle del Cauca. Colombia.
Los muestreos se llevaron a cabo utilizando diez redes de niebla (111 metros lineales), las cuales estuvieron activas desde las 18:00 hasta las 24:00 horas, con revisiones cada 30 minutos. El esfuerzo de muestreo se determinó multiplicando el número de redes utilizadas en cada remanente por el número de horas netas que las redes estuvieron abiertas por cada noche efectiva (Fenton et al., 1992); para el presente estudio el esfuerzo total fue de 4800 horas.red-1 (8 noches x 10 redes x 6 horas x 10 remanentes). Todas las redes se ubicaron de manera ad libitum a nivel de sotobosque, considerando que los murciélagos frugívoros tienden a forrajear a esta altura (Bernard, 2001; Kalko et al., 2008), además, se tuvo en cuenta la presencia de plantas que se encontraban en fructificación, claros y bordes de bosque y refugios potenciales. Cada individuo capturado fue depositado en bolsas de tela a la espera de la excreción de las heces por un periodo máximo de 4 horas y finalmente liberado la misma noche de captura. Posteriormente, las heces fueron recolectadas en bolsas de papel y rotuladas con el código del individuo para análisis posteriores. Cuando fue posible, los individuos capturados se identificaron en campo utilizando la Clave de identificación de los murciélagos de Sudamérica (Díaz et al., 2016) considerando medidas morfométricas y caracteres morfológicos externos; posteriormente, los individuos fueron marcados y liberados. Cuando fue necesaria la revisión de caracteres craneales se efectuó la colecta del individuo; para ello, los individuos fueron inyectados con una solución de lidocaína al 2% directamente en el corazón utilizando una jeringa de insulina de 1 mL, este procedimiento se realizó rápidamente para evitar mayor estrés o dolor a los individuos (Sikes & Gannon, 2011). Los especímenes colectados fueron depositados en el Museo de Ciencias Naturales Federico Carlos Lehmann Valencia (IMCN:MAM110-IMCN:MAM200) y en la Colección de Mamíferos de la Universidad del Valle (UV:ColeccMamifUV:15960-15987). Todas las muestras y colectas realizadas se efectuaron bajo el permiso de colecta otorgado por parte de la Corporación Autónoma Regional del Valle del Cauca (CVC) en la Resolución No. 1122 de 2018.
Adicionalmente, en cada uno de los remanentes se tomaron muestras de frutos provenientes de plantas que podrían ser fuente de alimento para los murciélagos frugívoros, con el fin de establecer una colección de referencia de semillas útil para la identificación de las muestras de las heces. Para esto, se realizaron recorridos diurnos en los bordes e interior de bosques buscando plantas en estado de fructificación. Las plantas y los frutos colectadas fueron depositadas en el Herbario TULV del INCIVA y en el Herbario de la Universidad del Valle (CUVC) (Carvajal-Nieto et al., 2021). Posteriormente, los frutos fueron disectados para extraer las semillas y estas fueron depositadas en tubos Eppendorf con alcohol al 70%.
Fase de laboratorio
Se realizó la limpieza de las muestras de heces con agua y alcohol para separar las semillas del resto de materia orgánica, utilizando pinzas y punzones. Posteriormente las semillas se revisaron a través de un estereoscopio Leica EZ4 para realizar su identificación taxonómica. Para la identificación se empleó la guía de semillas de Lobova et al. (2009), Ríos et al. (2004) y las semillas de referencia obtenidas durante la fase de campo (Carvajal-Nieto et al., 2021).
Análisis de datos
Con el fin de determinar cómo es la estructura de la red de interacción resultante entre murciélagos frugívoros con las plantas que consumen, se evaluaron propiedades topológicas de la red, que permiten entender cómo se dan las interacciones planta-animal: 1) para cuantificar el anidamiento de las redes de cada sitio se usó el índice WNODF (Almeida-Neto & Ulrich, 2011), el cual varía entre 0, cuando no existe anidamiento, y 100, cuando el anidamiento es perfecto y 2) para cuantificar la modularidad se utilizó el algoritmo DIRT-LPA-wb+ (Beckett, 2016), que maximiza la modularidad en redes bipartitas y que es una modificación del algoritmo LPAwb+ (). DIRTLPAwb+ calcula varias veces LPAwb+ (sensible a la inicialización de módulos) con diferentes inicializaciones aleatorias de módulos del nodo, elegidas entre μ posibles etiquetas únicas; y devuelve la solución que encuentra el mayor puntaje de modularidad. La significancia de los índices WNODF y DIRT-LPA-wb+ se estimó con un modelo nulo en el cual se generan 1000 matrices aleatorias a partir de la matriz original con el algoritmo de Patfield (1981), implementado en la función r2dtable del paquete stats del lenguaje de programación R (R Core Team, 2022). Posteriormente, se comparó el valor observado con el intervalo de confianza del 95% para los valores de WNODF y DIRT-LPA-wb+ del modelo nulo, de tal forma que si el valor observado es mayor a los valores del intervalo de confianza se considera una red anidada o modular, si el valor observado es menor a los valores del intervalo de confianza se considera una red menos anidada o modular de lo esperado al azar, y sí el valor observado está dentro del intervalo de confianza se considera que la red no es anidada o modular.
Adicionalmente, para evaluar la robustez, que es un procedimiento basado en la extracción al azar de especies (aleatorio, especialistas y generalistas), para la cual el área debajo de cada curva se calculó como una medida de la robustez de toda la red (R); donde R = 1 representa un sistema en el que la mayoría de las plantas o animales permanecen después de las extracciones, mostrando una mayor robustez y R = 0 representa un sistema que se derrumba después de haber eliminado las primeras especies, mostrando una menor robustez (Burgos et al., 2007). Para esto, los datos de las semillas encontradas en las heces se organizaron en matrices por sitio y a nivel general, donde las especies de murciélagos estuvieron en las filas y las especies de plantas en las columnas. De esta forma, se observa la variación de la estructura de la red en el tiempo y se analiza el sistema teniendo en cuenta la fuerza de interacción. Todos los análisis se hicieron a través del paquete Bipartite (Dormann et al., 2019) del software R (R Core Team, 2022).
Resultados
En total se capturaron 1142 individuos de murciélagos, agrupados en 36 especies, de las cuales 28 pertenecieron a la familia Phyllostomidae. En el presente estudio se tuvieron en cuenta únicamente las especies de filostómidos con registros de semillas en sus heces, teniendo así 991 individuos agrupados en 18 especies (Anexo 2). Se registraron 85 interacciones entre 18 especies de murciélagos y 19 especies de plantas, contabilizando al menos 19669 semillas, donde la mayoría pertenecieron a la especie de planta Piper aduncum (11636 semillas). Por parte de los murciélagos, la especie que aportó mayor número de semillas fue Carollia perspicillata (n=10228). En la Figura 2 se muestran algunas de las especies de murciélagos y plantas que presentaron interacciones.

Figura 2 Ejemplos de especies de murciélagos y plantas con interacciones. A. Carollia perspicillata, B. Sturnira ludovici, C. Artibeus lituratus, D. Dermanura phaeotis, E. Piper aduncum, F. Piper umbellatum, G. Ficus insipida, H. Psidium guajava. Escala 1mm en todos los casos.

Figura 3 Red de interacción resultante entre murciélagos frugívoros y plantas en el BsT del Valle del Cauca.
Las especies de murciélagos que mayor número de interacciones presentaron con especies de plantas fueron C. perspicillata con 14, A. lituratus con 13, D. phaeotis con 11, C. brevicauda con 10 y C. castanea con nueve, las demás especies de murciélagos presentaron seis interacciones o menos. En cuanto a las plantas, las que más interacciones presentaron fueron P. aduncum, Muntingia calabura, P. umbellatum, P. arboreum y Solanum rugosum. Es importante mencionar que algunas especies de murciélagos que no son frugívoros estrictos se encontraron dentro de las especies de las cuales se logró colectar muestras de semillas, tales como Glossophaga soricina, G. commissarisi y Phyllostomus hastatus, dónde G. soricina presentó interacción con seis especies de plantas aportando un total de 513 semillas, seguida por G. commissarisi que interactuó con M. calabura (140 semillas) y P. hastatus presentó relación con F. maxima (1 semilla) (Figura 3).
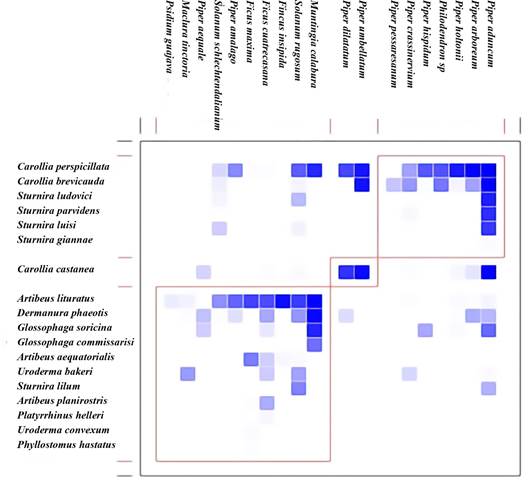
Figura 4 Grado de modularidad entre las especies de murciélagos y plantas de acuerdo con la densidad de las interacciones entre especies.
Por otro lado, al evaluar las propiedades estructurales de la red de interacción se encontró que la red general presentó una estructura menos anidada de lo esperado al azar, con un valor del índice WNODF de 30.29, el cual fue menor al intervalo de confianza, IC: 73.20 - 82-16 (Tabla 1). Asimismo, se determinó que la red general es modular ya que el índice DIRT_LPAwb+ presentó un valor de 0.31 el cual es mayor al intervalo de confianza, IC: 0.01 - 0.02 (Tabla 1). En la Figura 4 se evidencia la conformación de tres módulos principales de especies tanto de plantas como de murciélagos, en el primer módulo se observa la afinidad entre las especies del género Piper con especies del género Carollia y Sturnira; en el segundo módulo se agrupan las especies P. umbellatum y P. dilatatum con C. castanea, mientras que en el tercer módulo se agrupan las especies que presentan una mayor diversidad de interacciones.
Al evaluar la robustez a través del área bajo la curva (R) para la red de interacción general, se encontró que tanto las especies de plantas como de murciélagos presentaron valores similares para los tres métodos de extracción de especies utilizados. Teniendo en cuenta el modelo aleatorio los valores de R fueron 0.79 para plantas (Rp) y 0.74 murciélagos (Rm). Al extraer primero las especies menos abundantes a las más abundantes (especialistas) se encontró que el valor del índice fue más alto para plantas Rp = 0.95 en comparación con el obtenido para murciélagos Rm = 0.86. Por último, el valor del índice a través de la extracción de las especies más abundantes a las menos abundantes (generalistas) fue de Rp = 0.44 y Rm = 0.48, respectivamente (Tabla 1). Lo anterior indica que tanto las especies de plantas como de murciélagos son más susceptibles a la extinción al perder las especies generalistas, mientras que bajo los otros modelos de extracción de especies ambos grupos presentan valores de R cercanos a 1; por ende, son más robustos a la pérdida de especies bajo un modelo aleatorio y de eliminación de las especies especialistas.
En la Tabla 1 se observan los resultados obtenidos de las propiedades estructurales de las redes de interacción para cada remanente evaluado. Se evidenció que para todos los sitios las redes fueron menos anidadas de lo esperado al azar y a pesar de que todos los valores de modularidad fueron menores a 0.55, representan una estructura modular. Por otro lado, para la robustez se encontró que, teniendo en cuenta el método de extracción aleatorio y de especialistas tanto plantas como murciélagos presentaron valores similares; siendo el Bosque comunitario de Higueroncito (H) el sitio que presentó el valor más bajo para murciélagos bajo el método de extracción de las especies especialistas. Por el contrario, al evaluar el método de extracción de las especies generalistas se encontró que, para la mayoría de los sitios, la red es más susceptible a la pérdida de especies de plantas, a excepción de la red para A (PNR Mateguadua) e I (Hacienda El Medio); resaltando que bajo este método se presentan los valores más bajos de robustez tanto para plantas como para murciélagos.
Tabla 1 Valor de los índices de anidamiento (WNODF), modularidad (DIRT_LPAwb+) y robustez (R) para la red de interacción de murciélagos y plantas en diez remanentes de bosque seco tropical del Valle del Cauca. Representados con el código de letra asignado y para el conjunto de datos de la matriz completa. IC (Intervalo de confianza), P (Plantas) y M (Murciélagos).

Remanente: (A) Parque Natural Regional Mateguadua, (B) Reserva Forestal Protectora Nacional Quebrada La Valenzuela, (C) Hacienda El Volcán, (D) Hacienda La Venta, (E) Predio La Gitana, (F) Reserva Natural Chimbilaco, (G) Parque Natural Regional El Vínculo, (H) Bosque comunitario de Higueroncito, (I) Hacienda El Medio y (J) Hacienda Verdún.
Discusión
De acuerdo con la red de interacción resultante (Figura 3) se pueden observar las relaciones ampliamente descritas en literatura sobre la relación entre los murciélagos del género Carollia y Sturnira con frutos del género Piper y Solanum, respectivamente. Esto deja en evidencia un comportamiento diferente por parte de las especies dentro de Carollia y Sturnira, respecto a las demás especies de murciélagos. Asimismo, se destaca la interacción entre los murciélagos del género Artibeus y frutos del género Ficus (Giannini & Kalko, 2004; Sánchez et al., 2012). La red de interacción presentó un patrón menos anidado de lo esperado al azar, modular y robusto, lo que concuerda con lo encontrado en otros remanentes de BsT de Colombia (Velásquez-Roa & Murillo-García 2019). Contrario a lo esperado para redes de interacción murciélago-planta (Mello et al., 2011) no encontramos un patrón anidado, en el cual las especies más generalistas tienden a interactuar entre ellas formando un núcleo en la matriz de interacción, mientras que las especies más especialistas tienden a interactuar con las especies del núcleo (Bascompte & Jordano 2014, ). Una estructura menos anidada de lo esperado al azar se produce por la ausencia de interacción entre especies especialistas y el núcleo de especies generalistas (Dormann et al., 2017). Este patrón puede resultar si las especies más generalistas de murciélagos (A. lituratus y C. perspicillata) pueden ser dominantes y monopolizar especies de plantas (F. cuatrecasana y P. aduncum, respectivamente) forzando a las especies especialistas a alimentarse de ciertas especies de plantas (). Por el contrario, de acuerdo con lo esperado en las redes mutualistas se encontró una estructura modular (Olesen et al., 2007) donde hay grupos de especies que están más conectados entre ellos que con especies de otros grupos, lo cual se evidencia en nuestros resultados (Figura 4). Los módulos resultantes dejan en evidencia que el primer módulo es conformado por especies del género Piper y especies del género Carollia y Sturnira; en el segundo módulo se aprecia la separación de las especies P. umbellatum y P. dilatatum y C. castanea, mientras que el tercer módulo agrupa especies con diversas interacciones. Lo anterior indica que la modularidad observada no es máxima y, por ende, no se observa una compartimentalización marcada de cada módulo, lo cual es concordante con Castaño et al. (2018). De acuerdo con esto sugerimos que la estructura modular de la red no es el resultado de interacciones aleatorias entre las especies de plantas y murciélagos en los diferentes remanentes, ya que al observar la identidad de las especies que conforman cada módulo en cada red por separado se evidencia que hay grupos de especies de murciélagos y plantas que interactúan fuertemente en diferentes remanentes, es decir que comparten los recursos.
Por otra parte, los resultados obtenidos en cuanto a la robustez evidencian que la red de interacción para el BsT es más susceptible a la pérdida de especies generalistas, pues los valores de R fueron cercanos a cero (Rp = 0.44 y Rm = 0.48). Nuestros resultados concuerdan con lo encontrado por Velásquez-Roa & Murillo (2019), para un BsT del Valle del Cauca, donde se encontró que la red podría colapsar rápidamente debido a la pérdida de murciélagos con muchas interacciones (generalistas). El hecho de que las especies generalistas sean más importantes para la estabilidad de la red de interacción resultante está en consenso con lo mencionado por Robinson & Strauss (2020), quienes encontraron que en hábitats con menos recursos las especies generalistas de una red de interacción promueven la estabilidad de esta. Carollia perspicillata fue la especie de murciélago con mayor número de interacciones y a su vez la más abundante, esta especie es considerada como un indicador potencial de hábitats deteriorados (Schulze et al., 2000), además de presentar una amplia variedad de frutas en su dieta, presentando tendencias generalistas. No obstante, esta especie presenta una alta preferencia por las infrutescencias de plantas del género Piper, representando hasta el 65 % de su dieta (Thies & Kalko, 2004; Lopez & Vaughan, 2007). Esto se pudo evidenciar en la alta cantidad de interacciones que C. perspicillata registró con múltiples especies del género Piper en el estudio. Además, tuvo una alta preferencia por Piper aduncum, incluso en presencia de especies similares. Este mayor consumo de P. aduncum por parte de C. perspicillata puede estar relacionado con el contenido nutricional de las infrutescencias, ya que se ha comprobado que esta especie presenta una mayor cantidad de lípidos y un mayor valor calórico (Lima & Reis, 2004), por lo cual representa una importante fuente de energía para los murciélagos. El alto consumo de Piper, que son plantas pioneras con características principalmente arbustivas (Figueiredo & Sazima, 2000) por parte de especies de murciélagos del género Carollia reafirma la importancia de estas interacciones para la regeneración de bosques, incluso en áreas perturbadas. Una de las ventajas del consumo de plantas pioneras por parte de murciélagos, es que, debido a sus hábitos alimenticios, la alta riqueza de especies y su movilidad, los quirópteros pueden jugar un rol determinante en la promoción de procesos como la dispersión de semillas y el flujo genético dentro y entre poblaciones de plantas (Dumont, 2003).
Adicionalmente, según diversos autores (Aizen et al., 2012; Chacoff et al., 2018; Resasco et al., 2021) las especies generalistas son la base de una red de interacción estable, pues perduran en el tiempo, lo cual podría indicar una mayor tolerancia a cambios y perturbaciones en el hábitat en el que se encuentren. Sin embargo, la relevancia de estas especies en los ecosistemas es comúnmente subestimada debido a sus altas abundancias, a pesar de ser el eje principal en las complejas redes de interacción que mantienen el equilibrio del ecosistema. En el contexto de los remanentes de Bosque seco Tropical evaluados, las especies como C. perspicillata y P. aduncum, recobran mayor importancia ya que estos remanentes se encuentran aislados entre sí, varían en tamaño y en estado de conservación y por ende existe variación en la oferta de recursos que pueden llegar a reconfigurar las interacciones de las especies, evidenciando interacciones con plantas que generalmente crecen fuera de los bosques como Psidium guajava (Castaño et al., 2020); por lo que las especies generalistas resultan más especializadas en hábitats con pocos recursos, incrementando así la estabilidad de las redes de interacción (Robinson & Strauss, 2020), sin embargo, no dejan de ser susceptibles a grandes reducciones en el tamaño de sus poblaciones a causa de fuertes cambios ambientales (Wagner, 2020). Por esta razón, es importante incluir a las especies generalistas en los esfuerzos de conservación y planes de restauración de áreas naturales, los cuales usualmente se enfocan sólo en especies raras (Lindenmayer et al., 2011). Además, la presencia de los generalistas es considerada particularmente importante para la persistencia de las especies especialistas, contribuyendo a que la red sea resiliente ante perturbaciones (Bascompte et al., 2003). De acuerdo con los resultados, especies vegetales pioneras como aquellas del género Piper y especies abundantes de dispersores como C. perspicillata y A. lituratus representan la base para soportar la red de interacción dentro del BsT. Lo anterior toma mayor relevancia al tratarse de especies que se encuentran en el BsT, pues al aportar a la estabilidad de interacciones planta-animal, promueven la restauración de este ecosistema tan altamente amenazado, ya que dada la configuración espacial de los remanentes de bosque se hace necesaria la intervención por parte de especies con roles ecológicos claves como los murciélagos. Además, los murciélagos debido a sus hábitos pueden ser un grupo focal en la implementación de planes de restauración para el BsT.
Conclusiones
Con los datos obtenidos se concluyó que la red de interacción resultante para diez remanentes de BsT del Valle del Cauca presenta una estructura menos anidada que lo esperado al azar, modular y robusta a la pérdida de especies especialistas, mientras que resalta el rol fundamental que están llevando a cabo las especies generalistas al mostrar susceptibilidad a la pérdida de estas. No obstante, se debe considerar la complejidad de las relaciones existentes entre las comunidades de especies presentes en los hábitats, incluyendo las especies especialistas, lo cual implica preservar la mayor cantidad de diversidad posible que permita conservar funcionalmente dichas especies en los ecosistemas. Adicionalmente, los resultados hacen un llamado de atención sobre la inclusión de especies con hábitos generalistas en estrategias de conservación y en procesos de implementación de planes de restauración del BsT, ya que con la información que aportan promueven el mantenimiento del ecosistema en el que se encuentran y ayudan a la regeneración natural de los bosques.



