











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los datos primarios de biodiversidad (DPB) son observaciones particulares de especies en un lugar y tiempo determinado (Troia y McManamay 2016). En los últimos años, el acceso a este tipo de datos se ha incrementado debido a grandes esfuerzos para liberarlos a través de plataformas como "Global Biodiversity Information Facility" (GBIF), SpeciesLink (http://splink.cria.org.br/), eBird (https://ebird.org/home), VertNet (http://www.vertnet.org/), entre otras. A pesar de los avances recientes en la compilación de los DPB, la heterogeneidad en la calidad de la información es un obstáculo recurrente para inferir patrones de biodiversidad a diferentes escalas (Rondinini et al. 2011, Troia y McManamay 2016). Diferentes autores han reportado sesgos en la distribución de datos espaciales de fauna y flora, especialmente en áreas altamente biodiversas (Amano y Sutherland 2013, Pimm et al. 2014, Amano et al. 2016). Adicionalmente, lugares con grandes vacíos de información a nivel espacial usualmente albergan especies y ecosistemas amenazados (Amano y Sutherland 2013). Estos sesgos dificultan la priorización de proyectos de investigación, el mapeo de riesgos y la toma de decisiones sobre dónde se deben invertir recursos para proteger especies y ecosistemas (Ceballos et al. 2005). En este sentido, existe la necesidad de realizar análisis rigurosos que permitan identificar vacíos de información en los DPB que son utilizados en estudios de ecología y conservación (Troia y McManamay 2016).
Diferentes factores influyen la calidad y cantidad de los DPB. Sesgos relacionados con la obtención de los DPB están asociados con su ubicación geográfica (Meyer et al. 2015) , el tipo de ecosistema (Peterson et al. 2010), el grupo de estudio (Chapman y McCaw 2017) y los criterios de los investigadores, quienes pueden ignorar ciertos registros dependiendo del objetivo de muestreo (Amano et al. 2016) . Adicionalmente, pueden existir sesgos asociados a la disponibilidad de datos científicos a lo largo del tiempo (e.g. Gardner et al. 2014). La identificación de sesgos temporales de los DPB es fundamental, puesto que una buena cobertura temporal es necesaria para monitorear aspectos tales como las variaciones en los patrones de distribución de las especies con respecto a los cambios en las condiciones ambientales (Boakes et al. 2010, Brummitt et al. 2015), así como para proveer una línea base sobre la riqueza de especies en áreas conservadas (Willis et al. 2007). Por lo tanto, los análisis sistemáticos sobre la distribución temporal y espacial de los DPB son importantes para obtener una visión holística del estado de la biodiversidad.
Nuestro conocimiento sobre los patrones de diversidad de los mamíferos colombianos es escaso, a pesar de ser un grupo con una alta diversidad a nivel global (Ceballos et al. 2005, Ramírez-Chaves et al. 2016a), y a que es particularmente vulnerable a presiones antrópicas (Davis et al. 2018). En Colombia, los DPB de mamíferos silvestres han aumentado progresivamente desde el año 2000, cuando se estableció la plataforma de Sistema de Información sobre Biodiversidad de Colombia - SIB Colombia (nodo nacional de GBIF). A través de esta plataforma, se han liberado 8 318 941 registros de mamíferos (https://www.gbif.org/). Sin embargo, no hay estudios sistemáticos sobre cómo los esfuerzos de muestreo han contribuido a disminuir los sesgos espaciales en el conocimiento de los mamíferos del país. Además, hacen falta análisis que combinen información de vacíos de información sobre los patrones de diversidad y distribución de los mamíferos y los diferentes tipos de presiones antrópicas. Esto es particularmente relevante para el país, donde políticas en la última década están encaminadas a incrementar el desarrollo económico en áreas con baja presión antrópica, pero que cuentan con poco conocimiento sobre el estado de su diversidad (Andrade-C 2011).
En este trabajo presentamos un análisis sobre los patrones de distribución espacial y temporal de los DPB para los mamíferos de Colombia. Mediante el uso de bases de datos de biodiversidad, identificamos las áreas geopolíticas (i.e. departamentos) y las ecorregiones con los mayores vacíos de información a nivel de clase y de orden en cuanto al número de especies registradas con respecto al número de especies esperadas. Posteriormente identificamos las áreas con los mayores vacíos de información que sufren una mayor presión antrópica, con el fin de establecer prioridades de muestreo. Nuestro trabajo provee la primera síntesis sobre las fortalezas y debilidades de los DPB para mamíferos colombianos. Esperamos que estos datos ayuden a generar bases robustas no solo para priorizar la obtención de información sobre los patrones de la diversidad de especies en diferentes áreas, sino también para enfocar esfuerzos científicos que ayuden a estudiar y proteger localidades y grupos específicos.
MATERIALES Y MÉTODOS
Obtención de registros de mamíferos de Colombia
Los datos de presencia para todas las especies de mamíferos terrestres registradas en Colombia se obtuvieron a partir de registros clasificados como especímenes de colecciones biológicas u Observaciones de máquina del portal "Global Biodiversity Information Facility" en enero 2020 (GBIF c2020). Los datos fueron filtrados usando el paquete rgbif (Chamberlain et al. c2017) por medio de la función occ_search(). Los términos utilizados para descargar los datos pueden ser consultados en el Material suplementario S1. Además, obtuvimos los registros a partir de una consulta directa al SiB Colombia (https://sibcolombia.net/). El SiB Colombia hace cortes trimestrales (datos usados corte 01 oct 2019) y aplica protocolos de validación y limpieza que consisten en identificar datos inconsistentes con respecto al nombre científico y la jerarquía taxonómica, así como la coincidencia entre los topónimos geográficos y las coordenadas (SiB Colombia c2019).
Los nombres científicos fueron estandarizados con base en los compendios taxonómicos más recientes para el país (i.e.Ramírez-Chaves et al. c2019). Para ello, primero filtramos todas las especies registradas en las bases de datos analizadas que no correspondían con los nombres de la lista de Ramírez-Chaves et al. (c2019). Posteriormente identificamos las especies que constituyen sinónimos de acuerdo con bibliografía especializada (Gardner 2008, Patton et al. 2015, Mammal Diversity Database c2020) y descartamos especies no nativas, combinaciones duplicadas de nombres científicos, localidad, coordenadas y fecha, así como especímenes con coordenadas que no se encontraban dentro de los límites continentales del país o que no podían ser asignados a un departamento y ecorregión específica. Los registros con coordenadas incompletas o erróneas fueron eliminados con el paquete scru-br (Chamberlain c2016) disponible en R versión 3.6.2 (R Team Development Core c2019). Este proceso arrojó 14 751 registros para 418 especies, de los cuales 1 545 (10,4 %) corresponden a registros de trampas cámara y 13 206 (89,6 %) a registros de especímenes depositados en colecciones biológicas. El número de registros, sin descartar coordenadas duplicadas por especie, correspondió a 77 603, de los cuales 63 668 (82,0 %) corresponden a especímenes recolectados y 13 935 (18,0 %) a trampas cámara. Toda la rutina empleada para obtener los registros puede ser consultada en el Material Suplementario S1.
Patrones temporales de muestreo y representatividad de especies a nivel nacional
Los patrones temporales de muestreo a nivel de orden fueron identificados siguiendo la metodología propuesta por Meyer et al. (2016). Para ello, calculamos la diferencia media entre el número de años que hay entre la fecha de un registro particular y el registro temporal adyacente más cercano para celdas de 50 * 50 km, en el periodo comprendido entre 1950 y 2019, el cual abarca el 89 % de los registros. Esta métrica tiene valores altamente negativos si la cobertura temporal es baja, ya que indica grandes intervalos de tiempo entre fechas de muestreo adyacentes. Adicionalmente, estimamos el tiempo desde que el último registro fue recolectado para cada especie por ecorregión y por departamento. Estos valores fueron promediados para cada orden. Con el fin de tener un indicador del esfuerzo de muestreo en diferentes años, graficamos la cantidad de registros por orden a lo largo del tiempo. Además, realizamos una curva de acumulación de especies para evidenciar patrones de acumulación de especies a nivel nacional en áreas de 50 * 50 km usando el paquete vegan (Oksanen et al. 2013), también disponible en R versión 3.6.2.
Cálculo de la representatividad de especies por departamento y ecorregión
Con el fin de identificar las especies de mamíferos esperadas por departamento y ecorregión, los polígonos de departamentos y ecorregión (Olson et al. 2001) fueron interceptados con los polígonos de distribución disponibles en el sitio web de la Unión Internacional para la Conservación de la Naturaleza (IUCN c2019). Los nombres taxonómicos de los polígonos de la IUCN fueron estandarizados siguiendo la lista oficial de mamíferos de Colombia (Ramírez-Chaves et al. c2019) con el mismo protocolo descrito para la limpieza de los registros de bases de datos. En algunas ocasiones, los mapas de la IUCN no consideran la presencia de una especie en áreas con registros confirmados para el país. En estos casos, añadimos el departamento o la ecorregión correspondiente dentro del área de distribución esperada para la especie. En total obtuvimos información de la distribución para 438 especies de mamíferos terrestres, las cuales corresponden al 83 % de las especies terrestres reportadas por Ramírez-
Chaves et al. (c2019). En este proceso, nos aseguramos de que todas las especies con registros en las bases de datos consultadas (GBIF y SIB) contaran con información sobre los departamentos y las ecorregiones esperadas. Debido a que 22 especies con registros confirmados para Colombia no contaban con información disponible en los mapas de la IUCN, los departamentos y las ecorregiones esperadas fueron añadidos basados en una búsqueda de literatura especializada (e.g. Tavares et al. 2014, Patton et al. 2015, Chacón-Pacheco et al. 2018, Burgin et al. 2020). La representatividad del muestreo por ecorregión y departamento fue calculada como el porcentaje del número de especies esperadas que han sido registradas en la base de datos de mamíferos tanto en GBIF como en SIB Colombia. La clasificación de ecorregiones siguió la categorización de ecorregiones terrestres de Olson et al. (2001). Todos los análisis de representatividad fueron realizados a nivel de orden taxonómico.
Identificación de áreas prioritarias de muestreo
Con el fin de identificar áreas para realizar muestreos que maximicen el potencial de registrar nuevas especies, desarrollamos un modelo generalizado de disimilitud (GDM). Este modelo explica la disimilitud composicional entre pares de sitios a partir de diferencias en sus variables ambientales y la distancia geográfica entre los mismos (Ferrier et al. 2007). Dado que la adecuada estimación de la disimilitud composicional depende de la completitud del muestreo, para este análisis consideramos únicamente los datos de presencia de celdas de 5 km muestreadas al menos tres veces, con una completitud de muestreo de 75 % basado en el indicador no paramétrico Chao 2. La disimilitud entre todas las celdas muestreadas se estimó usando el índice de Jaccard. Para calcular la disimilitud ambiental entre sitios muestreados se emplearon datos de World-Clim 2 (Fick y Hijmans 2017) usando aquellas variables con una correlación de Pearson menor a 0,8 en el área de estudio (temperatura media anual, intervalo de temperaturas diurnas, isotermalidad, intervalo anual de temperatura, precipitación anual, precipitación del mes más seco y estacionalidad en la precipitación). El GDM se empleó para identificar los sitios que más complementarían el muestreo existente empleando el método de la p-mediana (Faith y Walker 1996), el cual identifica secuencialmente el sitio que más contribuye a disminuir la distancia promedio entre sitios muestreados y no muestreados. Todos los análisis se realizaron en el paquete de WhereNext de R (Velásquez-Tibatá c2019).
Una vez identificamos las áreas prioritarias de muestreo, usamos la capa de huella humana desarrollada por Correa Ayram et al. (2020) para identificar las áreas con los mayores vacíos de información que sufren una mayor presión antrópica. Luego de obtener la capa de complementariedad, excluimos todas las áreas urbanas usando la capa desarrollada por Liu et al. (2018). Adicionalmente, seleccionamos los pixeles de la capa de complementariedad con valores inferiores a 40 en la capa de huella humana (Correa Ayram et al. 2020), con el fin de identificar áreas con baja presión antrópica prioritarias para el muestreo.
RESULTADOS
Patrones temporales en el esfuerzo de muestreo y representatividad a nivel nacional
El esfuerzo de muestreo por año y por década no ha sido constante y la cantidad de registros se ha incrementado en campañas de muestreo llevadas a cabo en las décadas de los sesenta, ochenta y desde el 2016 (Fig. 1a). Además, la cobertura temporal en celdas de 50 * 50 km tiende a ser baja (Tabla 1). Para la mayoría de los órdenes, las celdas analizadas han sido muestreadas menos de cuatro años en promedio. Solo los órdenes Carnivora, Chiroptera, Didelphimorphia y Rodentia presentan celdas que han sido muestreadas al menos diez años, todas ellas en las regiones Andina y de la Orinoquia (Fig. S1). La diferencia media entre el número de años comprendidos para registros de una misma celda es más alta en Eulipotyphla (27,73 años), mientras que es más baja en Perissodactyla. Este último orden presenta valores bajos de número promedio de años muestreados y número máximo de años muestreado por celda (Tabla 1).

Figura 1 Patrones de representatividad y acumulación de registros de mamíferos terrestres continentales en Colombia a. Distribución temporal de registros por metodología utilizada durante el periodo 1950-2019 b. curva de acumulación de especies por sitio (celdas de 50 x 50 km).
Tabla 1 Número de registros por metodología de muestreo y cobertura temporal de muestreo para los órdenes de mamíferos terrestres continentales de Colombia. Registros únicos se refiere a registros no repetidos de especie, metodología, año y localidad; NPA = Número promedio de años muestreados para todas las celdas con presencia esperada por orden; NMA = Número máximo de años muestreados por celda; DMC = Diferencia media entre años consecutivos de muestreo para todas las celdas con presencia esperada por orden

Los órdenes con la mayor cantidad de registros comprenden aquellos con los valores más altos de riqueza de especies pequeñas. Chiroptera cuenta con la mayor cantidad de registros con combinación única de especie, localidad y fecha (n = 9 547), seguido por Rodentia con 2 554 (Tabla 1). En contraste, los órdenes con los menores registros comprenden especies de tamaño grande tales como Artiodactyla (n = 252) y Perissodactyla (n = 56), u órdenes que abarcan especies que habitan mayormente zonas andinas y altoandinas como Eulipotyphla (n = 55) y Paucituberculata (n = 43) (Tabla 1). La curva de acumulación de especies para toda la clase a nivel nacional no alcanzó una asíntota (Fig. 1b), lo que indica que el muestreo de la diversidad de mamíferos de Colombia es aún incompleto.
Distribución espacial de registros y representatividad por departamento y ecorregión
La representatividad de los registros de mamíferos terrestres de Colombia a nivel de departamento y ecorregión tiende a ser baja y altamente variable. Para departamentos, la representatividad promedio fue de 48 % (Tabla 2), mientras que para ecorregiones el promedio disminuyó a 42 % (Tabla 3). Existen claros sesgos espaciales en los patrones de recolección de información sobre la distribución de las especies de mamíferos terrestres en Colombia (Fig. 2). La mayoría de los datos provienen de la región Andina y Caribe. Los departamentos de Santander, Caldas, Tolima, Cundinamarca (Región Andina) y Meta (Orinoquia) mostraron los mayores valores de representatividad (> 60 %) mientras que los departamentos con los menores valores (< 30 %) corresponden a Guainía, Vaupés, Guaviare (Amazonia) y La Guajira (Caribe), seguidos por Atlántico, Cesar, Arauca, Amazonas y Nariño (30-35 %) (Tabla 1). No todas las regiones donde se concentran los asentamientos humanos en Colombia presentan altos valores de representatividad. Por ejemplo, varios departamentos como Boyacá, Cesar y Quindío tienen valores de representatividad menores al 40 %. Otros ejemplos incluyen los bosques húmedos al occidente de Ecuador en el departamento de Nariño, así como los bosques húmedos del Catatumbo en Norte de Santander, los cuales no alcanzan el 30 % de representatividad.

Figura 2 Patrones de representatividad espacial y temporal de registros de mamíferos terrestres continentales en Colombia. a. Distribución de registros de especímenes recolectados y trampas cámara (observaciones de máquina). b. Número medio de años con registros para todos los órdenes. c. Porcentaje de representatividad, basado en el número de especies registradas vs esperadas por departamento y d por ecorregión.
Tabla 2 Porcentaje de representatividad por departamento para las especies continentales de mamíferos terrestres presentes en Colombia. Los departamentos se encuentran organizados del menor al mayor porcentaje de representatividad.

La variación en los patrones de representatividad por ecorregión es mayor. Aunque más del 50 % presenta una representatividad menor al 50 % (por ejemplo, los Páramos de la Sierra Nevada de Santa Marta y las Planicies sedimentarias en el sur de la Amazonia), otras alcanzan niveles superiores al 85 % (Tabla 3). Las áreas con los mayores valores de representatividad tienden a presentarse en las tierras bajas de la región Andina y al Occidente de la región Caribe (Tabla 3, Fig. 2). Este es el caso de los Bosques del valle del Magdalena con valores superiores al 70 %, seguidos por Bosques Húmedos del Valle del Cauca (67,9 %), Arbustos Xerófilos de la Región Guajira-Barranquilla (66,5 %) y Bosques montanos de la cordillera Oriental (62,3 %). Una excepción a este patrón es la región de los Llanos de la Orinoquia, donde áreas de sabanas inundables y altillanuras cerca del piedemonte llanero tienen una muy alta representatividad. En contraste, bosques de tierra firme en las regiones de la Guyana y la Amazonía se encuentran pobremente representadas. Aunque algunos departamentos como el Meta muestran una alta representatividad para toda la clase, varias ecorregiones dentro de estos se encuentran pobremente estudiadas. Este es el caso de los Bosques Húmedos Transicionales entre la Orinoquía y la Amazonía en los límites con los departamentos de Guaviare, Caquetá y Vichada.
Tabla 3 Porcentaje de representatividad por ecorregión para las especies continentales de mamíferos terrestres presentes en Colombia. Las ecorregiones se encuentran organizadas del menor al mayor porcentaje de representatividad.

Los patrones espaciales de representatividad para toda la clase son similares a nivel de orden, con los mayores valores en departamentos y ecorregiones de la región Andina (Fig. 3, Material suplementario Figs. S2-S6). A nivel de ecorregión, la mayoría de los órdenes presentan una representatividad menor al 50 %, con los valores promedio más bajos para Carnivora (27,9 %). Adicionalmente, encontramos que una alta representatividad a nivel de ecorregión no se relaciona necesariamente con un adecuado muestreo por departamento. Este es el caso de Paucituberculata, un orden con una alta representatividad promedio por ecorregión (75 %), pero con una baja representatividad departamental (37 %; Fig. 3).
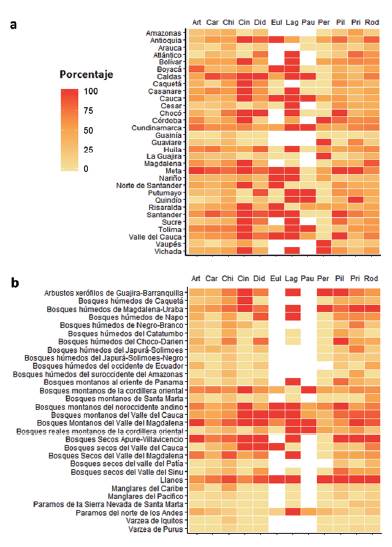
Figura 3 Representatividad del muestreo por a departamento y b ecorregión para los órdenes de mamíferos terrestres presentes en Colombia. La representatividad aumenta con la oscuridad del tono. Art= Artio-dactyla, Car= Carnivora, Chi= Chiroptera, Cin= Cingulata, Did= Didelphi-morphia, Eul= Eulipotyphla, Lag= Lagomorpha, Pau= Paucituberculata, Per= Perissodactyla, Pil= Pilosa, Rod= Rodentia.
La representatividad no estuvo relacionada con el número de especies esperadas por orden. Por ejemplo, mientras algunos órdenes con pocas especies esperadas como Lagomorpha y Paucituberculata tienen altos valores de representatividad a nivel de ecorregión (68,9 y 75 % respectivamente), otros como Artiodactyla (37,3 %) y Perissodactyla (30,6 %), que cuentan con cerca de quince especies esperadas para el país, están pobremente representados a nivel de ecorregión y departamento (Fig. 3).
Identificación de áreas prioritarias de muestreo
El análisis de complementariedad muestra que las zonas donde se maximizará el registro de especies se encuentran en los bosques tropicales de la Amazonia en los límites entre los departamentos del Caquetá y Amazonas, y el escudo Guayanés en la región de la Orinoquia (Fig. 4a). Así mismo, otras zonas prioritarias se encuentran distribuidas en una gran proporción de los bosques del departamento del Vaupés, las sabanas hiperestacionales y de altillanura plana en el departamento del Vichada, y las sabanas inundables de los departamentos de Casanare y Arauca. Aunque las áreas prioritarias con una alta presión antrópica tienden a presentar valores más bajos de complementariedad (Fig. 4b), nuestro análisis identifica áreas que incluyen bosques secos adyacentes a la Sierra Nevada de Santa Marta, así como los Bosques húmedos tropicales de la Amazonia en la región del Guaviare y la región de los Llanos en los límites entre los departamentos de Casanare, Vichada y Meta.

DISCUSIÓN
Este estudio presenta el primer análisis sistemático sobre los patrones de distribución de registros para los mamíferos silvestres de Colombia. A pesar de que existen esfuerzos importantes para incrementar el conocimiento sobre la diversidad de este grupo de fauna en el país (González-Maya et al. 2016, Ramírez-Chaves et al. 2016a,b, Morales-Martínez et al. 2018), los análisis demuestran que persisten vacíos importantes de conocimiento para la mayoría de órdenes y para regiones que son consideradas altos focos de biodiversidad. Este es el caso de los Páramos de la Sierra Nevada de Santa Marta, las Planicies sedimentarias del sur de la Amazonia y los Manglares del Pacífico. Aunque estudios previos han encontrado una fuerte correlación entre el número de especies registradas y las zonas de mayor influencia antrópica (Reddy y Dávalos 2003), la distribución espacial de los vacíos de información varia para diferentes órdenes y no se limita a regiones aisladas. En este sentido, aumentar la cobertura y representatividad de los DPB requiere análisis detallados de riesgo de transformación de paisaje donde existe una alta probabilidad de que ocurran procesos de extinción local.
Nuestros análisis muestran que no existe un incremento claro en el esfuerzo de muestreo a lo largo del tiempo. Los mayores esfuerzos recolección de mamíferos (Fig. 1a) corresponden a picos producto de estudios de enfermedades zoonóticas en mamíferos silvestres durante las décadas de los 60 y 70, especialmente por parte de C. J. Marinkelle (González-Astudillo et al. 2016). El pico más reciente en los DPB de mamíferos corresponde al auge de las trampas cámara como método de muestreo. Además, la curva de acumulación de especies no alcanzó una asíntota (Fig. 1b), lo que demuestra que el muestreo de la diversidad de especies de mamíferos en Colombia es aún incompleto y que cabe la posibilidad de encontrar nuevas especies en áreas pobremente muestreadas. Estos resultados concuerdan con estudios previos que muestran que la tasa de incremento de registros en el GBIF para varios grupos de vertebrados es baja e incluso ha disminuido en regiones como el Neotrópico (Collen et al. 2008, Gardner et al. 2014, Amano et al. 2016). Resolver estos sesgos de información es prioritario para proveer evidencia robusta sobre variaciones de los patrones temporales y espaciales de la biodiversidad en Colombia.
Existe una alta heterogeneidad en los patrones de representatividad de los diferentes órdenes de mamíferos analizados. Aunque los órdenes que incluyen especies de pequeños mamíferos son los que presentan mayor representatividad espacial y temporal, nuestro análisis encontró vacíos para diferentes ecorregiones del país. Por ejemplo, las especies de Eulipotyphla que habitan las mayores elevaciones de los Andes, aún están escasamente representadas en colecciones (ver Gardner 2008, Noguera-Urbano et al. 2019). Una situación similar ocurre con las especies endémicas de pequeños mamíferos como roedores, marsupiales y murciélagos, que en muchos casos abarcan especies conocidas por menos de cinco localidades (e.g.Suárez-Castro et al. 2017). Un ejemplo son las ocho especies endémicas de murciélagos registradas en Colombia, las cuales son conocidas en menos de 25 localidades (Ramírez-Chaves et al. 2020). Aunque algunos órdenes como Paucituberculata cuentan con especímenes recolectados para más del 75 % de las especies en todas las ecorregiones esperadas, el número de especímenes y la cobertura temporal es baja. Por lo tanto, los resultados muestran que una amplia cobertura espacial no se relaciona necesariamente con una representatividad alta en número de registros.
El tipo de metodología utilizada para registrar las especies, así como los patrones de riqueza, también ha influenciado los patrones de registro de especies. El mayor número de registros proviene de especies de murciélagos (Chiroptera) y roedores (Rodentia), los cuales abarcan la mayor riqueza en el país, con más de 130 cada uno (Ramírez-Chaves et al. c2019). Estos órdenes son generalmente registrados por medio de redes y trampas de captura viva, metodologías que son relativamente económicas y ampliamente utilizadas en inventarios rápidos. Además, el desarrollo de muestreos exhaustivos para la evaluación de patógenos de pequeños mamíferos, para los cuales se han analizado más de 60 000 muestras de Colombia (González-Astudillo et al. 2016), ha contribuido fuertemente a un mayor número de localidades muestreadas, especialmente de la región Andina y el Caribe. A pesar de estos esfuerzos, la representatividad de muestreo de órdenes de mamíferos que incluyen especies de tamaño pequeño es menor al 50 % en varias de las ecorregiones analizadas. Por otro lado, los órdenes de especies de tamaño grande como Artiodactyla y Perissodactyla, presentan los mayores vacíos de información, lo que se debe, en gran medida, a que el uso de trampas cámara para registrar estas especies es relativamente reciente.
El menor esfuerzo de muestreo ocurre generalmente en la zona cis-andina de Colombia, especialmente en el extremo oriental de la Orinoquía y la transición Orinoquía-Amazonía (departamentos Guainía y Vichada), así como en la región Amazónica (departamentos de Vaupés y Amazonas), en la frontera con Venezuela, Brasil y Perú. Por ello, varias especies, especialmente de pequeños mamíferos como Didelphimorphia, Chiroptera y Rodentia, cuya presencia ha sido detectada en dichos países (Tabla 4), carecen de registros verificados en Colombia (Gardner 2008, Patton et al. 2015, Burgin et al. 2020). Dada la cercanía de los registros a la frontera con Colombia, y a que estas especies se encuentran en ecorregiones existentes en este último, es posible que más de 20 especies de pequeños mamíferos registradas en Brasil, Ecuador, Panamá y Perú se encuentren en Colombia en las áreas pobremente muestreadas (Gardner 2008, Patton et al. 2015, Tabla 4).
Tabla 4 Especies de mamíferos terrestres con presencia probable en Colombia basados en registros confirmados en países limítrofes

Es clara la necesidad de realizar esfuerzos sistemáticos para llenar los principales vacíos de conocimiento y disponibilidad de los DPB en Colombia. Nuestros resultados muestran que dichos esquemas deberán considerar tanto focos de biodiversidad en zonas aisladas, como también ecorregiones cercanas a asentamientos humanos con mayor presión antrópica que pueden sufrir extinciones locales a corto plazo. Debido a que los recursos para realizar inventarios son limitados, el uso de herramientas de ciencia ciudadana (Chandler et al. 2017), así como el análisis riguroso de datos producto de estudios de impacto ambiental, puede constituir una fuente importante de información en áreas bajo una fuerte presión antrópica. En este sentido, se necesita continuar con el fortalecimiento institucional para reportar y curar la información proveniente de múltiples fuentes.
La apropiación y liberación de los DPB en Colombia ha avanzado notablemente, y este hecho es visible en las cifras reportadas por GBIF, ya que, entre las diez publicaciones de registros de mamíferos de Colombia más notables, la mayoría provienen de instituciones del país (Instituto Humboldt, Universidad Nacional de Colombia, Universidad del Valle, Fundación Panthera Colombia, Universidad de Caldas y Corantioquia; GBIF c2020). Además, algunas áreas en el país ya cuentan con diagnósticos de conocimiento de mamíferos (IAvH c2020), así que el siguiente paso consiste en comparar los datos de estos reportes con los patrones reportados en este estudio para enfocar esfuerzos en nuevas áreas de muestreo.
Nuestro trabajo constituye el primer análisis sistemático de la distribución de los esfuerzos de muestreo y generación de DPB para los mamíferos en Colombia. La variada representatividad de especies y registros en las ecorregiones y unidades político-administrativas de Colombia, resalta la necesidad de priorizar evaluaciones ecológicas rápidas, el fortalecimiento de las colecciones biológicas y la liberación de información mediante bases de datos en línea. En la medida que mayor número de registros y de mayor calidad estén disponibles, se mejorarán considerablemente los esfuerzos de planificación, regulación y diseño de estrategias de conservación para la gestión adecuada de la biodiversidad en Colombia.
Todos los autores estuvieron involucrados en las discusiones metodológicas y aportaron en la escritura del documento final.







