











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
Angiosperms are the largest group of land plants worldwide, representing a lineage with high diversity and widely distributed across the globe (Forzza et al. 2010, APG IV 2016). With the diversification of the angiosperms over time, plant cataloging has always been a major challenge for science and the interest in estimating the richness of the world's flora has always been present in the Neotropical region (Antonelli and Sanmartin 2011). Plant cataloging in the Americas has a secular history, but it was only in the last few decades that an overview of all floras has been made possible, mainly by the digitalization and compilation of botanical data (Plant Gateway 2018). The most recent work of Ulloa et al. (2017) presented an integrated assessment of all known native species of vascular plants in the Americas, resulting in 124 993 species, 6227 genera and 355 families, which corresponds to 33 % of the 383 671 species of vascular plants known worldwide.
For Brazil, which corresponds to almost half (48 %) of the total area of the Neotropical region (Schultz 2005), among the first studies on floristic cataloging is that by Martius et al. (1883) with Flora brasiliensis (1840-1906) which includes descriptions of 22 767 species of terrestrial plants. After this publication, botanical knowledge expanded as new species were described, and several names were synonymized through floristic and monographic studies (Forzza et al. 2012). Attempting to catalog plant species in Brazil, the Flora do Brasil 2020 project aimed to elaborate a checklist of the Brazilian flora, reporting 35 550 species of seed plants, 3468 genera, and 266 families (Flora do Brasil 2020 c2019), of species angiosperms recorded for Brazil 18,688 are endemic to the country (BFG 2018, Flora do Brasil 2020 c2019). Despite these data, there are multiple gaps and uncertainties on information regarding species distribution, which remains a central problem in ecological and conservational studies (Meyer et al. 2016). Another problem is the centralization of biodiversity research, making some areas very well-sampled due to being closer to research institutions, while other distant areas are poorly sampled (Magnusson et al. 2016).
The State of Maranhão figures as one of these floristic areas with reduced sampling despite presenting some published floristic checklists. It is currently the 19th State of Brazil (out of 27) in floristic wealth (BFG 2015). The State also appears in the Brazilian scenario as one with the most underestimated for biodiversity (Dias et al. 2009), with only 3080 species of angiosperms, out of which 674 are endemic to Brazil (Flora do Brasil 2020 c2019). The predominant phytogeo-graphic domain in the State of Maranhão is the Cerrado, one of the main tropical ecosystems on the planet, with several endemic species (Myers et al. 2000). Additionally, the Cerrado domain is regarded as a global hotspot for biodiversity conservation (Mittermeier et al. 2004) due to its high level of threat and the loss of already ca. 88 thousand hectares (46 %) of native vegetation cover (Strassburg et al. 2017). The Cerrado from the State of Maranhão is one of the more affected areas by anthropization processes, while only 2122 species of angiosperms (475 of them endemic to Brazil) are currently known in the Cerrado from this State (Flora do Brasil 2020 c2019). Additionally, there are few conservation units in the State of Maranhão and even fewer within the Cerrado domain in this State. The only exception is the Inhamum Municipal Environmental Protection Area (IME-PA), located in the Eastern region of this State, comprising an important fragment of Cerrado vegetation (Barros 2012).
Despite some minor floristic studies carried out in IME-PA in the past, such as those from Neres and Conceição (2010), Conceição et al. (2012), and Silva et al. (2016), no floristic synopsis has ever been carried out in this area. This fact indicates uncertainties regarding the total number of angiosperm species in IMEPA since these previous studies have not fully explored the area. Considering that the IMEPA is a strategic area for the knowledge on Cerrado vegetation from the State of Maranhão, this study presents a catalog of the angiosperms in IMEPA, providing data on the most representative taxa, phytophysiognomies of occurrence, life forms, endemicity to Brazil, new records for the State, and a floristic similarity analysis with other Cerrado areas in the State of Maranhão.
MATERIALS AND METHODS
The State of Maranhão is located in a transitional zone between three macro-regions of Brazil, North, Northeastern, and Midwestern; with an extension of 333 365.6 km2. Its climate and phytogeography cover the Amazon and Cerrado domains, with the additional influence of the Caatinga domain in an ecotonal zone (Galinkin et al. 2004, IMESC c2008, IBGE c2020). The climate varies from humid to semi-humid, with an annual average temperature of 26.2 °C and precipitation values between 800 and 2800 mm. The rainy season occurs between October and May, and the dry season between June and September (MMA 2011).
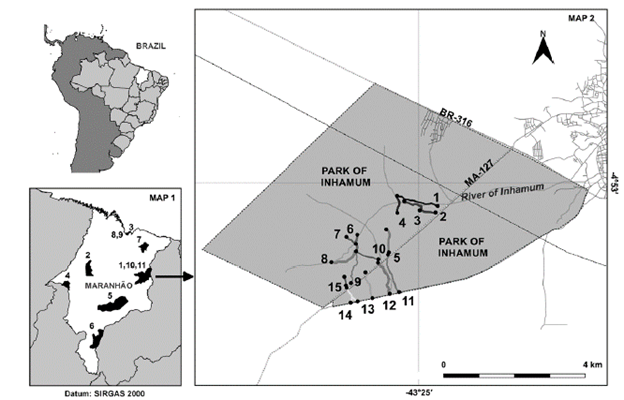
The Inhamum Municipal Environmental Protection Area (IMEPA) is located in the Eastern region of the State of Maranhão (Fig. 1) (04°53' South, 43°24' West), in the municipality of Caxias. The IMEPA is located within the Physiographic Zone of Itapecuru, with approximately 3000 hectares (Caxias 2001). The predominant soils in this State are the red-orange latosol, red-yellow podzolic, sands, and alluvial. Its vegetation is characterized by typical Cerrado phytophysiognomies, including gallery forests (Albuquerque 2012).
(Source: IBGE 2016, SILVA 2019).
Figure 1 Cartographic representation of the Municipal Environmental Protection Area of Inhamum and the State of Maranhão. Map 1. State of Maranhão with the municipalities used in the floristic similarity: 1, 10, 11. Caxias; 2. Buriticupu; 3. Raposa; 4. Imperatriz; 5. Mirador; 6. Tasso Fragoso and Parnaíba; 7. Urbano Santos; 8, 9. São Luis.; Map 2. Delimitation of the Inhamum Municipal Environmental Protection Area with the determination of the collection trails listed from 1 to 15
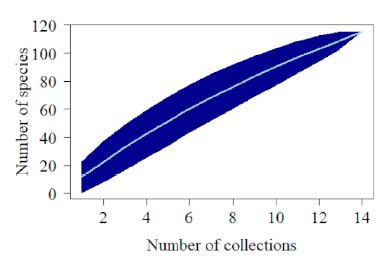
Figure 2 Species accumulation curve (continuous line light blue), 95% confidence intervals (dark blue shading) for the State of Maranhão collected in Inhamum Municipal Environmental Protection Area, Caxias, Maranhão.
Collection expeditions were carried out biweekly from January 2018 to June 2019 in different phytophysiognomies of IMEPA through exploratory walks. Fertile specimens (with flowers and/or fruits) were collected and processed according to usual methods in plant taxonomy (Gadelha-Neto 2013). For sampling and floristic survey of specimens, was carried out "active search" method of Filgueiras et al. (1994) that consisted of walking pre-existing trails on the edges and interiors of physiognomies within IMEPA, making collections of fertile specimens.
The collected specimens were identified at the specific level by comparison with herborized specimens, specialized literature (Souza et al. 2013, Souza and Lorenzi 2014, 2019, Souza et al. 2018), virtual herbaria, taxonomic keys, and by sending samples to specialists in the groups for confirmation and/or identification of specimens, following the classification of APG IV (2016). Additional samples of IMEPA specimens deposited in the HABIT herbarium (Thiers C2019) were also analyzed. Samples already identified in the herbarium were analyzed, and the determinations were verified, and the unidentified materials were posteriorly identified as abovementioned. To estimate the floristic richness, we quantified the number of species, genera, and families, indicating the most representative taxa (Magurran 2004). To ensure the correct spelling of species names, we consulted digital platforms such as Specieslink (CRIA c2020), Tropicos (c2019), and IPNI (c2012). For life forms, categorization of phytophysiognomy of occurrence, and endemicity for Brazil, we followed the Flora do Brasil 2020 (c2019) website. The species geographic distribution was also verified using the Flora do Brasil 2020 (c2019) and Specieslink (CRIA c2020) repositories. For classification of species according to Cerrado phytophysiognomies, we followed IBGE (2012) and Flora do Brasil 2020 website (c2019).
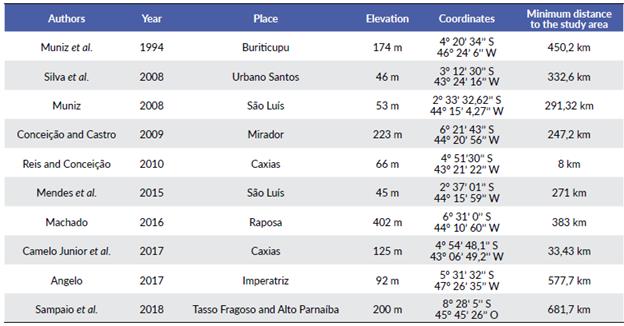
An analysis of floristic similarity was carried out using checklists from ten other floristic studies with a similar methodology in the State of Maranhão (Table 1). A record matrix of presence and absence for 627 species was compiled from the abovementioned studies. The similarity index was calculated by a Cluster Analysis obtained with a similarity index of Jaccard (Krebs 1999). Grouping was verified by the method of paired groups, with the elaboration of a dendrogram using the R 3.2.0 software (R Development Core Team c2009). An accumulation curve was also drawn up from the analyzed data to verify whether the sample size can directly influence species richness. Thus, the ideal was to mitigate wealth for the same number of individuals, using the rarefaction method (Magurran 2011). These analyses were performed using the vegan package implemented in the R 3.2.0 software (R Development Core Team c2009).
RESULTS AND DISCUSSION
A total of 446 angiosperms specimens, comprising 228 species, 148 genera, and 50 families were recorded in the study area (Fig 1). This species number is quite representative when considering the diversity of the Cerrado flora of Northeastern Brazil. The Neotropics comprise a series of phytogeographic and phytophysiognomy factors, including transitional zones, as those from the State of Maranhão, which are greatly influenced by the Amazon, Cerrado and Caatinga domains (MMA c2007, MMA c2016). When relating the collected data with the territorial area IMEPA in km2, the area shows fifteen collections per km2, demonstrating that it has a representative collecting effort. According to sufficiency indexes proposed in the literature, such as Shepherd (2003), an adequate sample value for tropical areas is three collections per km2. As for the number of species recorded, there is a considerable increase in the floristic knowledge at IMEPA, considering that the most representative study done so far recorded only 116 species of angiosperms (Silva et al. 2016). This study added a total of 112 species to the IMEPA angiosperms checklist. The IMEPA angiosperm flora corresponds to about 8 % of the species from Cerrado in the State of Maranhão (Flora do Brasil 2020 c2019).
The species accumulation curve using the rarefaction method (Fig. 2), demonstrated that the species richness for the sample area is much greater than the numbers shown in the study. Inferring that the sample size directly influenced the species richness, mainly due to the accumulation curve not being stabilized, indicating that a greater sample sufficiency must be performed.
It is unanimous according to Schilling and Batista (2008) and Schilling et al. (2012) that the use of the species accumulation curve to determine the sampling sufficiency is problematic. The concept assumes that the plant community is a spatially discrete entity with fixed and defined species composition. In tropical forests, on the other hand, the definition of community boundaries becomes particularly difficult due to the high richness of species, so the curve does not stabilize even with large sample sizes. Although the rarefaction curve demonstrates the advantage of establishing a confidence interval with statistically significant differences between the curve and the sample area studied (Meyer et al. 2016).
The most representative family in this study was Leguminosae with 54 species (Fig. 3), as expected, given the high number of species of this family in Brazil (BFG 2015). As for the genera, Cyperus (Cyperaceae) was the richest at IMEPA with seven species. Cyperus comprises about 550 species of cosmopolitan distribution and high representativeness in the tropics (Goetghebeur 1998). In Brazil, a total of 125 species of Cyperus are recorded with a wide distribution in the country, colonizing different phytophysiognomies, but mainly wetlands and drylands, such as savannas, pastures and rocky outcrops (Goetghebeur 1998, Souza et al. 2019).
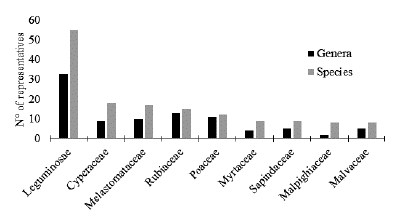
Figure 3 Distribution of the representativeness of genera and species by family of Angiosperms in the Inhamum municipal Environmental Protection Area, Caxias, Maranhão.
Analyzing all species life forms, we verified that 55 % (125 species) falls within a single life form, and 45 % (104 species) were collected with more than one life form. Tree was the most recorded life form for all species, corresponding to 38 % (88 species) of the taxa (Fig. 4). Throughout their diversification history, many families of angiosperms such as Leguminosae, Malvaceae, and Myrtaceae developed varied life forms, adapting to different conditions and environments where they live (Judd et al. 1999), these were quite common in the survey.
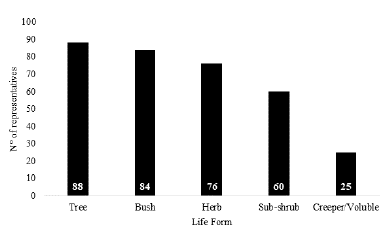
Figure 4 Distribution of life forms by species of Angiosperms in the Inhamum Municipal Environmental Protection Area, Caxias, Maranhão.
Combretum mellifluum Eichler (twelve specimens) and Desmoscelis villosa (Aubl.) Naudin (ten specimens) were the most collected species in IMEPA. Combretum mellifluum was recorded in small populations but it is widely distributed, occurring in several phytophysiognomies ("cerradao", gallery forest, and "campo sujo"). Desmoscelis villosa was found in a single population with several individuals associated to gallery forests, recorded in practically all watercourses in the area. These characteristics explain the high number of collections recorded when compared to other species in the survey. Desmoscelis villosa is widely distributed in South America, being registered in the Northeast of Brazil with several populations for the states of Bahia, Ceará, Piauí and Maranhão (Martins 2009).
When analyzing the most representative phytophysiognomy in the number of species within IMEPA, we found that campo sujo comprised 105 species, followed by gallery forests with 96 species, and cerradão with 43 species, emphasizing that some species were recorded in more than one phytophysiognomy (thirteen species). However, most were restricted to a single phytophysiognomy. The campo sujo phytophysiognomy was characterized by a dense herbaceous cover, mainly grasses on which subshrubs and shrubs are quite common, and few small trees. The great variation of life forms of this phytophysiognomy is one of the main reasons why campo sujo is greatly diversified (Coutinho 2002, Tannus and Assis 2004).
IMEPA's angiosperm flora includes 39 endemic species to Brazil (Flora do Brasil 2020 c2019), corresponding to approximately l0 % of the endemic species in the Cerrado domain from the State of Maranhão. According to Giulietti et al. (2005), the identification of endemism or threatened species means a great step towards conserving biodiversity services, mainly by identifying priority areas for conservation.
The following ten species are new records of angiosperms for the State of Maranhão: Eleocharis maculosa (Vahl) Roem. & Schult, Ancistrotropis peduncularis (Kunth) A. Delgado, Alophia drummondii (Graham) R.Foster, Eugenia subterminalis DC., Palicourea grandiflora (Kunth) Standl, Sacciolepis angustissima (Hochst. ex Steud.) Kuhlm., Borreria multiflora (DC.) Bacigalupo & E.L.Cabral, Matayba inelegans Spruce ex Radlk, Serjania erecta Radlk., and Piriqueta racemosa (Jacq.) Sweet (Fig. 5). For these species, only A. peduncularis is widely distributed in Brazil, with few States without records of it, demonstrating that its occurrence in the State of Maranhão was already expected. The species E. maculosa, E. subtermi-nalis, S. erecta, P. racemosa, and S. angustissima do not occur close to the State of Maranhão but show records in the Cerrado and Amazon domains. The species A. drum-mondii, B. multiflora, M. inelegans, and P. grandiflora are restricted in four to eight Brazilian States and distant from our study site, demonstrating that distribution expansion for these species was quite significant.

Figure 5 New records for the State of Maranhão collected in Inhamum Municipal Environmental Protection Area, Caxias, Maranhão. a. Ancistrotropis peduncularis; b. Borreria multiflora; c. Inflorescence of B. multiflora; d. Eleocharis maculosa; e. Inflorescence of E. maculosa; f. Piriqueta racemosa; g. Sacciolepis angustissima; h. Inflorescence of S. angustissima; i. Serjania erecta L.; j. Fruits of S. erecta; k. Matayba elegans; I. Fruits of M. elegans; m. Palicourea grandiflora; n. Eugenia subterminalis; o. Fruits of E. subterminalis; p. Alophia drummondii; q. Flower of A. drummondii. Examined material: a. SOUSA, D.H.S. 321 (HABIT); b-c. SOUSA D.H.S. 306 (HABIT); d-e. SOUSA, D.H.S. 196 (HABIT); f. SOUSA, D.H.S. 117 (HABIT); g-h. PINHO, A.N. 29 (HABIT); i-j. RODRIGUES, M, & CONCEIÇÃO, G.M. 27 (HABIT); k-I. SOUSA, D.H.S. 04; 179 (HABIT); m. MATOS & CONCEIÇÃO, G.M 53 (HABIT); n-o. SOUSA, D.H.S. 14; RODRIGUES, M. & CONCEIÇÃO, G.M 20 (HABIT); p-q. SOUSA, D.H.S. 93 (HABIT).
The State of Maranhão has several gaps in species distribution and records, mainly due to the low investment in extensive field studies. Despite the recent years, this perspective is changing with several new occurrences being published as those mentioned in the studies of Diniz et al. (2017), Santos-Silva et al. (2018), Nascimento et al. (2019), among others. With the recognition of these new records for the State of Maranhão, the geographic distribution of these species is expanded. According to Santos-Silva et al. (2018), initiatives like these contribute directly to the growing knowledge of the Brazilian flora and elaborate conservation strategies for phytophysiognomies and species cataloged in this study.
The floristic study carried out at IMEPA demonstrated a similar pattern to other surveys carried out in Cerrado fragments. Leguminosae, Cyperaceae, and Melastoma-taceae are generally mentioned as the richest families in species number (Neres and Conceição 2010, Conceição et al. 2012, Silva et al. 2016). The similarity analysis showed that the surveyed area is more related to the Flora of Buriti do Meio Environmental Protection Area in the municipality of Caxias, State of Maranhão, both in the eastern region of the state and close distances (33 km) (Junior et al. 2017) (Fig. 6). The two areas are located in the same Mesoregion of State of Maranhão, which explains the similar floristic aspects and the strong anthropic activities recorded in both of them. These areas are the direct result of the extensive fragmentation of the Cerrado domain, impacting most of the water resources in this domain and resulting in similar anthropogenic physiognomies (Françoso et al. 2015, Strassburg et al. 2017, Françoso et al. 2019). The clusters in the analysis also established very intrinsic geographic relationships with clusters in the east (6/7/10) and two clusters in the north center of the state (3/2/9) and (4/8).
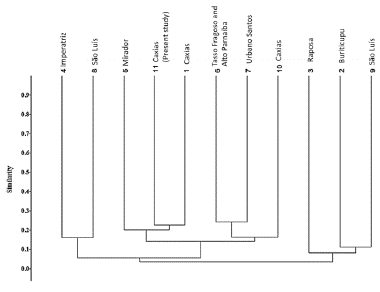
Another important point to be analyzed is that despite the formation of groups relating the studied floras, the values of similarity were low, varying from 0.1 to 0.3, with few shared species and many exclusive species, demonstrating that the floristic pattern is quite locally specific. Several studies show that geographic distance influences plants' distribution in tropical regions (Scudeller et al. 2001, Condit et al. 2002, Carneiro and Valeriano 2003), and attributes the variation in species richness and composition between places basically to the limitation of dispersion. This fact corroborates that the species similarity of a plant community decreases with the geographical distance in tropical regions, forming floristic compositions exclusive to certain places (Matos et al. 2013).
The characterization of the floristic composition of an area is important due to the recognition of new floristic records, categorization of endemic species, and elimination of distribution gaps, demonstrating that the surveys generate important information and provide consistent bases for the creation and improvement of conservation units (Gomes et al. 2004). Besides, there is no other way to get to know the regional flora than through the catalog and record of the representatives that compose it (Hopkins 2007).
This study synthesized the data on the angiosperm flora of IMEPA, recording the largest number of species up to date for the study site. It also presented important data for future conservation studies since the area is greatly affected by several anthropogenic processes. Based on the new records and endemic species in its composition, preservation efforts must be stimulated by reducing the biodiversity loss, filling the Cerrado knowledge gaps in the flora from the State of Maranhão.

