











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Los ecosistemas naturales en Colombia han sufrido grandes transformaciones antrópicas que han modificado la biodiversidad (Etter et al. 2006), afectando en consecuencia las interacciones entre poblaciones animales y vegetales (Márquez 2001). Uno de los ecosistemas más amenazados actualmente es el Bosque Seco Tropical (BST), debido a factores como la explotación de madera y la expansión de áreas agrícolas y ganaderas (Ulloa-Delgado 2016, Casas-Pinilla et al. 2017). Además, históricamente este ecosistema no ha recibido el cuidado suficiente para su conservación, por lo que actualmente en Colombia solo queda el 5 % (Baptiste et al. 2013). Entre las principales afectaciones al BST están las alteraciones en abundancia, riqueza e interacciones que sufren las poblaciones, ocasionadas por la ganadería intensiva (Ospina-López et al. 2015). Por ejemplo, se ha demostrado que, en ecosistemas afectados por la ganadería, el crecimiento y la diversidad vegetal se ven disminuidos, alterando a su vez parámetros microclimáticos como la radiación solar, la temperatura y humedad, y provocando la disminución en la abundancia y riqueza de las mariposas y afectando su distribución y composición (Collinge et al. 2003, Vélez Lemos et al. 2015).
El Departamento del Tolima, cuenta con 23 562 km2 en donde el 49 % de esta área es considerada ecosistema de BST (Gobernación del Tolima 2014), el cual está sujeto a diferentes niveles y fuentes de perturbación por lo que solo el 1,87 % está en buen estado, aunque sujeto a fuertes presiones por efecto de la ganadería extensiva y la agricultura (García et al. 2014).
Hasta hace relativamente poco, la mejor forma de describir la diversidad de una zona consistía en la caracterización de la diversidad alfa y beta empleando índices tradicionales. Estos índices además de arrojar un valor que indican si la diversidad de especies es alta o baja, poco permiten describir y analizar la diversidad en términos de la estructura, composición de especies y sobre todo en términos de sus interacciones, que son parte fundamental de una comunidad en la medida en que permiten entender los mecanismos que generan la coexistencia de las especies (Medel et al. 2009), además del mantenimiento de la diversidad biológica y su evolución (Thompson 2002).
Recientemente, el uso de análisis ecológico de redes ha permitido la descripción de la estructura de las comunidades de organismos en función de las especies que las componen y de sus interacciones a partir del desarrollo de parámetros que brindan información sobre estos elementos (Chagnon et al. 2012) tales como la diversidad y abundancia de interacciones en la red, los nodos generalistas y especialistas, la abundancia de nodos y la susceptibilidad de la red a las perturbaciones, entre otros (Blüthgen et al. 2008). En una gráfica de red ecológica, que representa por ejemplo la interacción mariposa-planta, los nodos son las especies de plantas y mariposas y los enlaces son líneas que representan su interacción y abundancia (Bascompte 2010). Dado que las comunidades son entidades dinámicas, las redes varían temporal y espacialmente, por ejemplo, con el tipo de hábitat (Hagen et al. 2012) y en función de las perturbaciones antrópicas, tales como la expansión de áreas agrícolas y ganaderas (Bascompte y Jordano 2007, Tylianakis y Morris 2017).
El orden Lepidoptera ha generado una estrecha relación evolutiva con las angiospermas, puesto que las mariposas se benefician de los recursos alimenticios que las flores les proveen en forma de polen y néctar y las plantas obtienen un beneficio reproductivo gracias a la polinización que las mariposas realizan (Waser y Ollerton 2006, Jordano et al. 2009, Boom-Urrueta et al. 2013). En razón a esto, esta interacción es definida como mutualista (Bascompte et al. 2006), sin embargo, no se puede desconocer que este grupo también presenta carácter herbívoro ya que en su etapa larval se alimentan de plantas estableciendo de esta manera una relación antagónica.
Los Lepidoptera han sido ampliamente reconocidos como un grupo de organismos muy sensible a las perturbaciones ambientales (González-Zuarth et al. 2014), por lo que es de esperarse que los cambios causados por acción humana afecten además de la diversidad de especies, sus interacciones.
Dada la necesidad de comprender la dinámica de los cambios en composición y diversidad de especies y de sus interacciones en respuesta a perturbaciones antropogénicas en un ecosistema altamente vulnerable, como es el Bosque Seco Tropical Colombiano, este estudio analizó y comparó la diversidad de especies, estructura y composición de las redes de interacción entre mariposas adultas (Lepidopte-ra: Papilionoidea y Hesperoidea) y angiospermas en floración entre zonas conservada e intervenida de Bosque Seco Tropical en diferentes épocas climáticas del año, en un sector del valle medio del Magdalena, Colombia.
MATERIALES Y MÉTODOS
Área de estudio
Este estudio se realizó en el municipio de Carmen de Apicalá, situado al oriente del departamento del Tolima, Colombia, en el piso térmico cálido con altitud de 328 m (Gobernación del Tolima 2014). Específicamente, el muestreo se adelantó en la finca La Sultana, localizada en la vereda La Antigua con un área total de 280 hectáreas, 4°10' Norte 74°46' Oeste. La finca está inmersa en ecosistema de Bosque Seco Tropical, en el que predominan las familias de árboles Fabaceae, Sa-pindaceae, Capparidaceae, Euphorbiaceae y Rubiaceae, encontrándose especies muy abundantes como Zanthoxylum monophyllum (Lam.) P.Wilson, Attalea butyracea (Mutis ex L.f.) Wess.Boer y Buddleja americana L., con plantas con troncos lisos con espinas, hojas compuestas y foliolos pequeños. La altura del dosel oscila entre los 15 y 25 metros (IAvH 1998) con hasta cuatro estratos vegetativos: herbáceo, arbustivo, palmeto y arbóreo; este último es el que predomina (Sanmartín-Sierra et al. 2016). Además de los fragmentos de bosque conservado, se encuentran pasturas para la ganadería, rastrojos y bosque secundario resultante de procesos de abandono de los potreros.
La precipitación está entre los 250 y 2000 mm anuales con dos períodos marcados de sequía (Ariza et al. 2014), su temperatura es de alrededor de 25 °C y sus climas son árido, cálido semiárido y cálido seco, caracterizados porque durante la mayor parte del año la evapotranspiración supera en gran medida a la precipitación. Esto genera déficit de agua, lleva a que la vegetación pierda parcialmente su follaje en dos períodos del año (IAvH 1998).
Muestreo y recopilación de datos
El muestreo se realizó durante diez días en el mes de abril en época de lluvia y diez días en agosto en época seca en 2019. En cada época se realizó el muestreo en dos zonas de estudio: zona de fragmentos de bosque conservado y zona con grandes áreas de expansión agrícola y ganadera. En cada una de estas zonas se definieron cuatro franjas de 50 x 4 metros, al ser áreas que en diversos estudios se registran como adecuadas para el muestreo de diversidad de especies, siendo además un factor que permite estandarizar el área de muestreo (Caranqui y Ortiz 2019, Jaramillo-Cruz 2021).
En primer lugar, durante los primeros cuatro días de cada salida se realizó un muestreo en dichas franjas con el fin de identificar las mariposas adultas y plantas de cada zona identificando las angiospermas en floración y recolectando las mariposas. Cada mariposa recolectada se sacrificó, almacenó y transportó al laboratorio de Ecología Evolutiva y Conservación de la Pontificia Universidad Javeriana, sede Bogotá, para su posterior identificación. Los ejemplares se depositaron posteriormente en la Colección de Entomología del Instituto de Ciencias Naturales ICN-MHN. Para la identificación se emplearon las claves y libros de D'Abrera (1987), Salazar y Vélez (1991), Constantino et al. (2002),

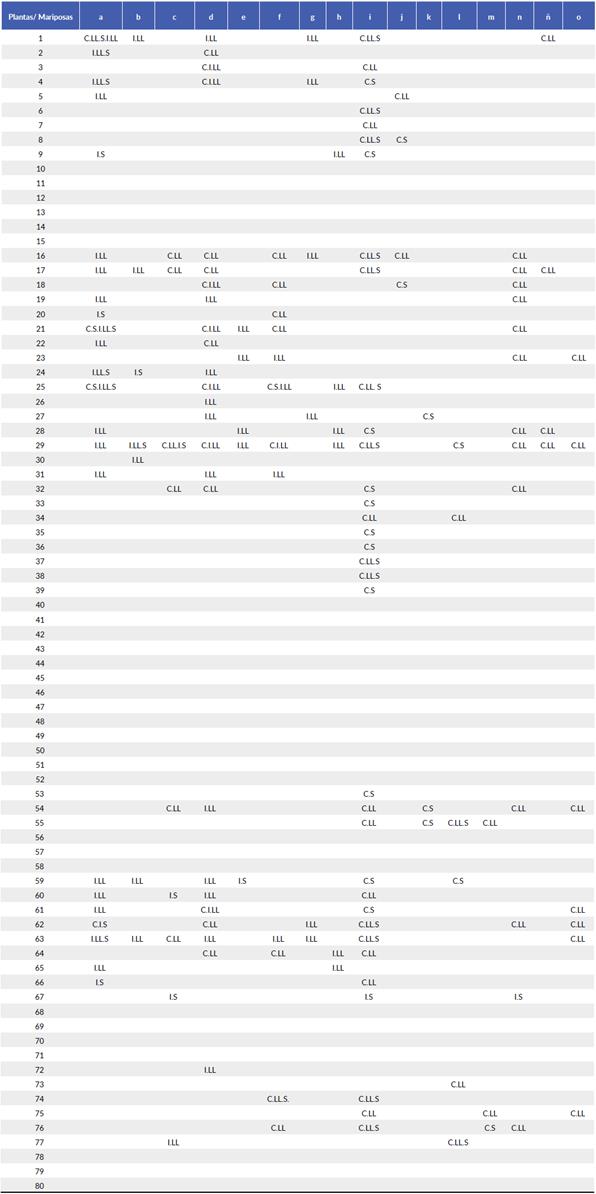
Figura 1 Redes ecológicas de Papilionoidea y Hesperoidea-Angiosper-mas en floración en una zona conservada y otra intervenida en época de lluvia y época seca, en el sitio de estudio. Los rectángulos blancos representan las especies de angiospermas y los negros las de mariposas. El ancho de cada rectángulo representa la abundancia de cada especie. El ancho de las franjas que conectan las plantas con las mariposas representa la abundancia de cada interacción. a. Red general para la zona conservada, b. Red general para la zona intervenida, c. Red para la zona conservada en época de lluvia, d. zona intervenida en época de lluvia, e. zona conservada en época seca, f. zona intervenida en época seca.
Andrade (2007) y Pulido (2010), y se contó con la ayuda de expertos como el curador de la colección de entomología de la Pontificia Universidad Javeriana, Giovanny Fagua quien facilitó claves para corroborar la identificación de las mariposas y Gonzalo Andrade-C quien permitió la entrada a la Colección de Entomología del Instituto de Ciencias Naturales ICN-MHN para corroborar la identificación de las mariposas por medio de la comparación con ejemplares. La identificación de las plantas en floración se realizó empleando el libro de Barrero et al. (2012) y mediante comparación con la colección de referencia del Herbario de la Pontificia Universidad Javeriana (HPUJ), sede Bogotá con ayuda del experto Carlos Andrés Ordoñez Parra.
Durante cada salida, y con el fin de registrar las interacciones mariposa-planta descritas anteriormente, se hicieron tres recorridos por cada una de las franjas, con una duración de 90 minutos cada vez. Los recorridos se hicieron entre las 7:00 am y las 05:00 pm. Cada vez que se observaba una mariposa en una flor se identificó la mariposa y la planta en la que se posó. Dado que las comunidades e interacciones cambian con el tiempo, y con el fin de obtener resultados claros sobre un período particular, los muestreos se realizaron en el pico de cada estación climática. Para maximizar el esfuerzo de muestreo, dos personas estuvieron recorriendo las franjas.
La diversidad alfa para cada zona en cada época del año se estimó con el índice de Margalef, el cual tiene en cuenta que hay una relación entre el número de especies y su abundancia (Moreno 2001); para ello se empleó el programa de acceso libre Past (Hammer 2001). La representa-tividad del muestreo se evaluó con el estimador de Chao 1. Si los valores son iguales o mayores al 85%, significa que se obtuvo un muestreo representativo (Villarreal et al. 2004); para su cálculo se empleó la versión 9.1.0 del programa Estimates (Colwell c2019). Se empleó el índice de similitud de Sorensen para estimar la diversidad beta de las mariposas diurnas en las dos zonas, empleando la herramienta de Excel (Villarreal et al. 2004).
El análisis de las interacciones planta-mariposa, se realizó a partir de la elaboración de redes ecológicas bipartitas (Jordano 1987, Medel et al. 2009), para las cuales se estimaron cinco métricas: Anidamiento, que refleja el nivel de compartamentalización de la red, es decir, nodos genera-listas y especialistas con sus respectivos enlaces (Ings et al. 2009, Barbour et al. 2016); Conectancia, que muestra la proporción de interacciones y de nodos en la red comparado con el número posbile de todas la interacciones (Harvey et al. 1983, Ings et al. 2009); Generalidad, que muestra el número promedio de enlaces por visitante floral (Maunsell et al. 2015); Índice de Shannon, que mide la diversidad de interacciones en la red (Maunsell et al. 2015), y robustez, que muestra que tan estable o frágil es la red, teniendo en cuenta los efectos de la disminución y extinción de las especies (Pocock et al. 2012). Se hicieron en total seis redes: una con interacciones totales para cada zona y otras cuatro que corresponden a cada zona por época climática.
RESULTADOS
Diversidad de especies
En total se identificaron 80 especies de mariposas en la finca La Sultana (Tablas 1 y 2). La zona conservada obtuvo un 22 % más especies que la intervenida. Por otro lado, la zona intervenida en época de lluvia obtuvo un 25 % más de especies que en época seca. El estimador Chao 1 en la zona intervenida fue de 92 % en las dos épocas, mientras que en la zona conservada fue de 73 % (Tabla 3), lo que demuestra la representatividad del muestreo, mucho más precisa para la zona intervenida que para la conservada; de otra parte, la mayor diversidad y abundancia se encontraron en la zona conservada en época de lluvia (Tabla 3). Además, se encontró una alta similitud de riqueza entre las dos zonas de estudio con un 69 %.
Durante los muestreos, 16 especies de plantas estaban en floración y fueron visitadas por las mariposas adultas. De éstas, siete fueron exclusivas de la zona conservada, cuatro de la zona intervenida y cinco se encontraban en las dos zonas (Tablas 1 y 2).
Redes de interacción mariposa-planta
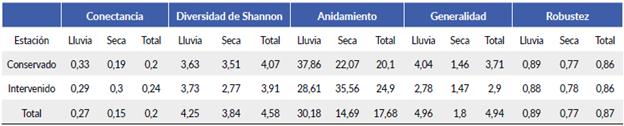
En general, la zona conservada obtuvo mayor cantidad de nodos y mayor diversidad de interacciones dentro de la red (diversidad de Shannon 4,07) en comparación con la zona intervenida (diversidad de Shannon 3,91). Además, esta última presentó mayor cantidad de mariposas especialistas, pues alcanzó una menor proporción de especies que interactúan con pocas especies de plantas, mientras que la zona conservada alcanzó una mayor proporción de especies que interactúan con muchas especies de plantas (generalidad: 3,71). La zona conservada obtuvo un mayor promedio de interacciones por nodo que la zona intervenida, además las dos zonas tuvieron la misma robustez (0,86), es decir, las dos tuvieron buena estabilidad de interacciones en la red dado que en los dos casos sus especies no disminuyeron en gran medida (Fig. 1a, Tabla 4).
La zona conservada en época de lluvia registró una menor cantidad de nodos y mayor diversidad de interacciones dentro de la red en comparación con la zona intervenida. Además, la zona conservada en época de lluvia obtuvo una proporción menor de interacciones que la zona intervenida y una proporción menor de especies de mariposas que interactúan poco con pocas especies de plantas; la zona intervenida obtuvo una mayor proporción de especies de mariposas que interactúan poco con pocas especies de plantas. Por otra parte, tanto la zona conservada como la zona intervenida en época de lluvia presentaron una robustez alta (0,89 y 0,88 respectivamente), es decir, las dos zonas presentan redes estables; sin embargo, la zona conservada presenta una estabilidad mayor reflejada en época de lluvia (0,89) que en época de sequía (0,77) (Figs. 2c-d, Tabla 4).
Para la zona intervenida en época seca, se obtuvo una menor cantidad de especies y de interacciones que en zona conservada. Por otro lado, la zona conservada en época seca obtuvo una proporción mayor de especies que interac-túan poco con pocas especies de plantas, lo cual representa una mayor cantidad de especies especialistas que en la zona conservada. La zona intervenida en época seca (0,78) presentó mayor robustez que la zona conservada (0,77); sin embargo, las dos redes son estables (Figs. 2e-f, Tabla 4).
DISCUSIÓN
En total, se registraron 80 especies de adultos de Papilio-noidea y Hesperoidea, asociadas con 16 especies de An-giospermas en floración, estableciendo un total de 591 interacciones (Tablas 1 y 2). De acuerdo con la literatura revisada, este constituye el primer estudio que establece asociaciones y redes de interacción entre la diversidad de estas superfamilias y Angiospermas en BST en el neotrópico comparando diferentes estados de perturbación en dos épocas del año. Algunos estudios previos, aunque son sobre redes ecológicas, se han concentrado en describir las redes entre mariposas adultas y angiospermas en floración en ecosistemas diferentes como el costero en donde comparan diferentes tipos de vegetación (Martínez-Adriano et al. 2018) o en zonas urbanas (Mukherjee et al. 2018), también en selvas tropicales comparando dos épocas del año (lluvia y sequía) (Mertens et al. 2021).
Tabla 1 Especies de mariposas adultas y angiospermas en floración en Bosque Seco Tropical Colombiano, Tolima-Carmen de Apicalá, finca La Sultana.

Tabla 2 Interacciones entre las especies de mariposas y angiospermas en zona conservada e intervenida en época de lluvia y sequía en Bosque Seco Tropical Colombiano, Tolima-Carmen de Apicalá, finca La Sultana. Las letras minúsculas representan las especies de angiospermas en floración y los números las especies de Papilionoidea y Hesperoidea que se denotan en la tabla 1. Las letras mayúsculas representan la zona y la época climática: Conservado (C) Intervenido (I) Lluvia (LL) Sequía (S). Las especies que no cuentan con interacciones, fueron encontradas en los primeros recorridos de reconocimiento.
En cuanto a la diversidad de especies, Mercado-Gómez et al. (2018) en un fragmento de Bosque Seco Tropical en María la Baja, departamento de Sucre, muestran una riqueza menor con 55 especies de Lepidoptera, con una duración mayor de muestreo (40 días y 120 horas). Casas-Pinilla et al. (2017) encontraron una diversidad de 121 especies de lepidópteros en La Mesa de los Santos, Santander con 36 días de muestreo y 360 horas de muestreo. Teniendo en cuenta el tiempo de muestreo de este estudio, podemos decir que 80 especies representan una muy buena aproximación a la diversidad de Papilionoidea y Hesperoidea de la región estudiada, datos confirmados por los resultados del estimador Chao1. 80 especies que interactuaron con 16 especies de plantas que en su momento estaban en floración, resultado por demás interesante en la medida en que Pizano y García (2014) registran para Colombia 2483 especies de angiospermas de BST y Villanueva et al. (2015) registran 1048 especies en los bosques secos del departamento del Tolima. Sería necesario un inventario de la diversidad de especies en el bosque de nuestro sitio de estudio con el fin de precisar la diversidad total de angiospermas y sus fluctuaciones temporales en floración y así establecer relaciones la forma en que las redes varían a lo largo de dichos periodos de floración. Sin embargo, a pesar de la aparente baja diversidad de especies de mariposas y plantas en floración, se registró una sorprendente cantidad de interacciones (591) lo que da cuenta de la importancia de evaluar además de la diversidad en términos de especies, la diversidad de interacciones, pues es esto lo que configura el uso que los organismos hacen de los recursos y en últimas la estructura de una comunidad.
La diversidad y la abundancia de mariposas más alta se dio en la zona conservada en época de lluvia (Tabla 3). Estos resultados concuerdan con los del estudio de Prince-Chacón et al. (2011) y Vargas-Zapata et al. (2011). Se registró alta similitud de riqueza de mariposas diurnas entre la zona conservada y la zona intervenida, es decir gran cantidad de especies compartidas en las dos zonas, este resultado concuerda con el estudio de Gallego-López y Gallego-Ropero (2019).
Tabla 3 Valores de diversidad y abundancia de especies de mariposas, y porcentaje de representatividad (Chao 1), para zona conservada e intervenida en época de lluvia y sequía, en la zona de estudio.
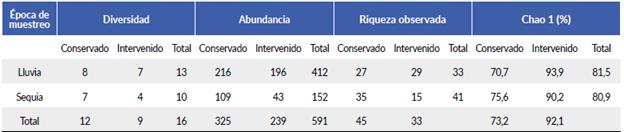
Tabla 4 Valores de conectancia, diversidad de Shannon, anidamiento, generalidad y robustez, de las redes ecológicas de Papilionoidea-Angiosper-mas en zona conservada e intervenida en época de lluvia y época seca, en Bosque Seco Tropical Colombiano, Carmen de Apicalá, Tolima.
En cuanto a las redes ecológicas, la mayor abundancia se evidenció en la zona conservada. En la zona intervenida de nuestra área de estudio, la extracción de madera y la disminución de fragmentos de bosque para generar pasturas ganaderas son factores que posiblemente contribuyen al patrón de interacciones observado. Así nuestros resultados concuerdan con Medel et al. (2009) quienes explican que las zonas conservadas presentan una mayor proporción de interacciones por nodo y una diversidad de interacción mayor que la de las zonas intervenidas.
En la época de lluvia tanto en la zona conservada como en la intervenida, la abundancia de interacciones fue más alta. Como afirman León-Camargo y Rangel-Ch (2015), es en la época de lluvia cuando se presentan los máximos picos de floración, lo que genera un efecto positivo en las interacciones. La zona conservada en época de lluvia obtuvo menor cantidad de nodos mostrando una proporción mayor de especialistas y mayor diversidad de interacciones, en contraste con la zona intervenida y con la época seca (Fig. 1c-f, Tabla 4). Lo anterior se debe a que la diversidad floral fue mayor en zona conservada y aumentó en épocas de lluvia lo cual proporciona cambios en la diversidad y abundancia de insectos que se alimentan de las angiosper-mas (Genung et al. 2010).
En conclusión, las redes variaron en función de la época y estado de conservación del ecosistema, influyendo esto directamente en su estructura, tal como se evidencia por ejemplo en la baja diversidad de Shannon y en el alto nivel de ani-damiento en la red de la zona intervenida en época seca, en donde las características ambientales son menos favorables por lo que la diversidad floral disminuye en comparación con la de la zona conservada en época de lluvia.
Estudios como este, además de contribuir al registro taxonómico de las especies de mariposas asociadas al Bosque Seco Tropical colombiano, brindan información valiosa sobre sus interacciones con angiospermas, contribuyendo de esta forma al cuerpo de conocimientos sobre las dinámicas de interacción que favorecen tanto a los poliniza-dores como a sus recursos y la forma en que varían tanto estacionalmente como en respuesta a factores de transformación antrópica. Adicionalmente, al complementar los análisis tradicionales de diversidad con el análisis de las interacciones mediante el establecimiento de redes ecológicas, se identifican los cambios en la configuración de las comunidades y sus implicaciones para los ecosistemas, brindando nuevos elementos para establecer estrategias de conservación de un ecosistema altamente degrado como es el Bosque Seco Tropical.

