











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Trichoptera es un orden de insectos holometábolos compuesto por el superorden Amphiesmenoptera (Holzenthal, et al., 2007; Malm, et al., 2013), que a su vez se divide en tres subórdenes: Annulipalpia, Spicipalpia e Integripalpia (Wiggins, 1996). Los estados inmaduros (larvas y pupas) de estos organismos se desarrollan en ambientes acuáticos con excepción de Enoicyla pusilla (Trichoptera: Limnephilidae) (Burmeister, 1839), en tanto que los adultos son terrestres y habitan muy cerca del lugar de donde emergen (Martínez-Menéndez, 2014). Las larvas tienen un papel importante en la trasferencia de energía y en la dinámica de los nutrientes de los ecosistemas acuáticos, ya que explotan una variedad de recursos y son fuente de alimentos para peces, aves y otros vertebrados (Flint, et al., 1999; González & Cobo, 2004; Domínguez & Fernández, 2009). Además, se utilizan como bioindicadores de la calidad del agua, pues algunas familias son muy sensibles a los cambios en las variables fisicoquímicas (Springer, 2006; Stein, et al., 2008; Jaramillo-Londoño, 2006), por lo cual responden adecuadamente a la polución de los sistemas acuáticos (Roldán, 2003; 2016).
A pesar de que en la última década se han desarrollado en Colombia importantes trabajos sobre la biodiversidad del grupo, la información disponible aún es incipiente (Holzenthal & Calor, 2017). Actualmente se conocen cerca de 211 especies distribuidas en 15 familias y 45 géneros (Muñoz-Quesada, 2004; Spies, et al., 2006; Domínguez & Fernández, 2009), siendo la región andina colombiana la de mayor número de registros, con 14 familias y 35 géneros (Zúñiga, et al., 1993; Ballesteros, et al., 1997; Rincón, 1999; Posada & Roldán, 2003; Guevara, et al., 2005; Guevara-Cardona & Reinoso-Flórez, 2013, 2014; López, et al., 2015). En la región Caribe se conocen 11 familias y 26 géneros en los departamentos Magdalena, Cesar y La Guajira, con 22, 15 y 13 géneros, respectivamente (Rúa-García, et al., 2015, Serna, et al., 2015, Jaimes-Contreras & Granados-Martínez, 2016). En Magdalena la mayoría de los estudios se ha centrado en aspectos ecológicos como los hábitos alimentarios (Granados-Martínez, 2013, Guzmán-Soto & Tamaris-Turizo, 2014), la deriva (Tamaris-Turizo, et al., 2013), la bioindicación (Guerrero-Bolaño, et al., 2003) y la distribución (Serna, et al., 2015). En este último estudio se incluye un aspecto ecológico relacionado con el gradiente altitudinal, pero en un reducido rango espacial y temporal (entre 100 y 400 m y durante seis meses de muestreo), pero no hay otros estudios a mayor escala espacial o temporal.
Los sistemas fluviales de montaña pueden albergar una gran diversidad de microhábitats: grava, hojarasca, salpicadura, piedras y macrófitas, que ofrecen muchos recursos y favorecen el establecimiento de una amplia variedad de taxones de macroinvertebrados (Steven, 1992). Los tipos y la cantidad de microhábitats pueden variar según el gradiente altitudinal Rapoport, 1982; Stevens, 1992), pues la geomorfología y la heterogeneidad del río cambian a medida que aumenta o desciende el gradiente altitudinal. El objetivo de este estudio fue establecer cómo varía la diversidad de larvas de Trichoptera en un gradiente altitudinal del río Gaira y cuál es la relación entre la abundancia de los taxones y las principales variables ambientales.
Materiales y métodos
Area de estudio
El río Gaira se ubica en la vertiente noroccidental de la Sierra Nevada de Santa Marta. Es la principal fuente de agua potable para la población de los sectores turísticos de Gaira y El Rodadero en el distrito de Santa Marta. El río limita al norte con la cuenca del río Manzanares, al sur con la del río Toribio, al este con el río Guachaca y al oeste con el mar Caribe. Tiene una extensión aproximada de 10,464 ha y una longitud de 32,53 km desde su nacimiento hasta su desembocadura en el mar Caribe (Frayter, et al., 2000; Tamaris-Turizo, et al., 2013). La cuenca tiene un rango altitudinal entre los 0 y los 2,750 m del lugar de su nacimiento en la cuchilla de San Lorenzo, que es la parte más alta de la cuenca y registra una precipitación promedio multianual de 2.010 mm, de 1.800 mm en la parte media y de 470 mm en la parte baja (Pro-Sierra, 2017). A lo largo de la cuenca del río Gaira se determinaron cuatro sitios de muestreo (Figura 1) cuya localización y características ambientales se documentaron detalladamente (Tabla 1).
Tabla 1 Localización y algunas características ambientales de los sitios de muestreo a lo largo del río Gaira

SL-TA: tramo alto, San Lorenzo; LC-TA: tramo alto, La Cascada; LV-TM: tramo medio, La Victoria; PM-TB: tramo bajo, Puerto Mosquito. Bmh-ST: bosque muy húmedo subtropical; Bh-ST: bosque húmedo subtropical; Me -T: monte espinoso tropical (Espinal & Montenegro, 1963)
Toma de muestras
Entre los meses de abril y mayo de 2018 se hicieron dos muestreos intensivos en cada sitio durante el periodo seco. En cada sitio se seleccionó un tramo representativo del afluente de 50 m, aproximadamente. Teniendo en cuenta el criterio de heterogeneidad microgeomorfológica y el sistema de rápidos y remansos (Dunne & Leopold, 1978; Frissell, et al., 1986; Angrisano & Sganga, 2009). Las muestras se recolectaron en los microhábitats de grava, macrófitas, hojarascas, piedras y zonas de salpicaduras. La grava y las macrófitas se muestrearon con una red Surber de 0,09 m2 y 250 de abertura de malla. Para la recolección de los organismos asociados con la hojarasca se usó una red triangular (250 de abertura de malla). Se hizo una revisión manual de las zonas de salpicadura y piedras (aproximadamente 20 cm de diámetro) y estas últimas se rasparon cuidadosamente con cepillo de dientes durante 10 minutos. Las muestras tomadas de los diferentes microhábitats se unificaron formando una muestra compuesta, la cual se preservó con etanol al 96 % en un frasco de 800 ml previamente rotulado con los datos de cada sitio de muestreo. Por último, las muestras se almacenaron y se transportaron en neveras plásticas al Laboratorio de Limnología del Grupo de Investigación en Biodiversidad y Ecología Aplicada (GIBEA) de la Universidad del Magdalena.
Variables ambientales
En cada sitio de muestreo se utilizó una sonda multiparamétrica WTW35i0, para medir las siguientes variables ambientales: temperatura del agua (°C), conductividad (μS/cm), oxígeno disuelto (mg/L) y pH. Además, se tomaron 400 ml de agua del río y se almacenaron en botellas plásticas que luego se refrigeraron y se transportaron al Laboratorio de Calidad del Agua de la Universidad del Magdalena para estimar las concentraciones de nitratos (NO3 μg L-1), nitritos (NO2 μg L-1) y fosfatos (PO4: μg L-1) siguiendo los protocolos propuestos por la 1.
Identificación organismos
Las muestras de individuos inmaduros recolectadas en campo se analizaron en estereoscopios ZEISS Stemi 305 y, de ser necesario, en microscopios ZEISS Primo Star para observar algunas estructuras morfológicas relevantes en la identificación del organismo a nivel de género siguiendo las claves y guías taxonómicas para larvas neotropicales: Roback (1966), Holzenthal (1988), Wiggins (1996), Merrit & Cummins (1996), Angrisano & Sganga (2009), Posada-García & Roldán (2003) y Springer (2006).
Análisis y procesamiento de datos
Se analizó la contribución de los géneros de tricópteros del río Gaira en cada sitio mediante del cálculo de la abundancia relativa. Para conocer la composición del orden se estimaron los números efectivos de taxones a nivel de género con la serie de números de Hill (N0, N1 y N2) (Jost, 2006, 2010), en la cual la diversidad de orden cero (N0) está representada por la riqueza genérica (S), N1 por el número de géneros abundantes estimado con base en el exponente del índice de diversidad de Shannon-Weiner (H1), y N2 por el número de especies muy abundantes, el cual se estima con base en el inverso de Simpson (X) (Chao, et al., 2014). Cabe aclarar que a pesar de que la conversión del número efectivo está diseñada para especies, también se ha empleado a nivel de géneros (Barragán, et al., 2016), con el fin de dar una interpretación más adecuada de los datos de abundancia respondiendo al concepto de diversidad a partir de la conversión de los índices de diversidad a número efectivo de especies (géneros en este caso). Para conocer la similitud entre sitios a partir de las abundancias de los ensamblajes, se hizo un análisis de conglomerados según una matriz de Bray-Curtis (con medida de distancia euclidiana). Para conocer la relación entre las abundancias de las larvas de tricópteros en los sitios de muestreo se recurrió a un análisis de componentes principales (ACP). Por último, para explorar la relación entre las variables ambientales (temperatura, pH, conductividad, nitratos, nitritos, oxígeno disuelto y fosfatos) y la abundancia de larvas de Trichoptera por sitio se hizo un análisis de correspondencia canónica (ACC). Los análisis de diversidad alfa se realizaron en la plataforma iNext (https://chao.shinyapps.io/iNEXTOnline/). Todos los análisis estadísticos se hicieron en el programa PAST (3.x) (Hammer, et al., 2001).
Resultados
Caracterización ambiental
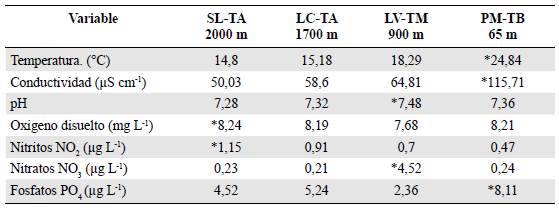
La mayor temperatura, conductividad y concentración de fosfatos se observó en Puerto Mosquito, con 24,84 °C; 115,71 cm-1 y 8,11 PO4 μg L-1, respectivamente, en tanto que la menor temperatura, conductividad y pH se registraron en San Lorenzo, pero también en este sitio se reportaron los valores más altos de nitritos y oxígeno disuelto (1,15 NO2 μg L-1 y 8,24 OD mg L-1, respetivamente). Por otro lado, la menor concentración de fosfatos se registró en La Victoria, con 2,36 PO4 μg L1. En La Cascada se presentaron las menores concentraciones de nitratos (0,21 NO3 μg L1) y los más bajos valores de nitritos se dieron en Puerto Mosquito, con 0,47 NO2 ug L1 (Tabla 2).
Estructura de comunidades
En total se recolectaron 443 larvas pertenecientes a 10 familias y 21 géneros, de los cuales 11 son nuevos registros para el río Gaira y uno es nuevo para Colombia (cf. Amazonatolica) (Anexo A,https://www.raccefyn.co/index.php/raccefyn/article/view/1148/2753). Las familias más diversas fueron Hydroptilidae (cinco géneros) y Leptoceridae (cuatro géneros), seguidas de Hydropsychidae, Phylopotamidae, Polycentropodidae y Glossosomatidae con dos, en tanto que las demás familias presentaron un solo género. Hydropsychidae fue la más abundante (40,6 %), seguida de Calamoceratidae (12,4 %), y Glossomatidae (1,6 %) y Helicosychidae (1,4 %) presentaron las menores abundancias (Anexo A, https://www.raccefyn.co/index.php/raccefyn/article/view/1148/2753). Entre los 21 géneros, Smicridea y Leptonema fueron los más frecuentes y abundantes, con 22,1 y 18,5 %, respectivamente (Figura 2).

Figura 2 Abundancia relativa de los géneros de las larvas del orden Trichoptera recolectadas en todos los sitios de muestreo del gradiente altitudinal en el río Gaira
cf. Amazonatolica, Cyrnellus, Nectopsyche y Polycentropus fueron los taxones exclusivos de la parte alta del río Gaira (San Lorenzo y La Cascada). En estos sitios se registraron abundancias de 97 y 112 y los menores valores de diversidad con 9 y 6 taxones, respectivamente (Tabla 3). Los taxones exclusivos encontrados en La Victoria (parte media) fueron Ochotrichia, Cerasmatrichia y Metrichia. Por su parte, en Puerto Mosquito (parte baja) se registró la mayor abundancia y diversidad de géneros (N0=13) (Tabla 3), siendo Protoptila, Mortoniella, Oecetis y Chimarra los taxones exclusivos de este sitio. En cuanto al rango de distribución altitudinal, los géneros que presentaron los rangos más amplios de distribución fueron Smicridea y Leptonema y Xiphocentron (Anexo A,https://www.raccefyn.co/index.php/raccefyn/article/view/1148/2753) (Figura 3). Los índices de completitud de los muestreos y de cobertura de las muestras (Anexo B,https://www.raccefyn.co/index.php/raccefyn/article/view/1148/2754) evidenciaron que el esfuerzo de muestreo estuvo cerca del 100 % en todos los sitios, con excepción de La Victoria (parte media del río).
Tabla 3 Abundancia e índices que valoran la estructura de los ensamblajes de larvas de tricópteros.

SL-TA: tramo alto, San Lorenzo; LC-TA: tramo alto, La Cascada; LV-TM: tramo medio, La Victoria; PM-TB: tramo bajo, Puerto Mosquito. N=número de individuos; H'=diversidad de Shannon-Weiner; λ=dominancia de Simpson; J'=equidad de Pielou; N0=riqueza; N1=taxones comunes; N2=taxones dominantes

El conglomerado de similitud mostró que Puerto Mosquito (parte baja) conformó un grupo separado de los demás sitios de muestreo por sus altas abundancias; en contraste, La Cascada (parte alta) y La Victoria (parte media) fueron los sitios más similares, conformando un grupo aparte del sitio San Lorenzo, que presentó las menores abundancias (Figura 4). Por otro lado, el análisis de componentes principales (ACP) presentó una varianza acumulada del 87,5 % en los dos primeros ejes. En este análisis se destacaron la gran abundancia de Chimarra y Smicridea, Wormaldia y Zumatrichia en Puerto Mosquito. La Victoria se destacó por la gran abundancia de Metrichia y Cerasmatrichia, San Lorenzo por la de Phylloicus y Polycentropus y La Cascada por la de Atopsyche (Figura 5).

Figura 4 Conglomerado de similitud de las abundancias de los géneros de tricópteros en los sitios de muestreos

Figura 5 Análisis de los componentes principales de los géneros de tricópteros con relación a los cuatro sitios de muestreo.
En el análisis de correspondencia canónica (ACC) en los dos primeros ejes la varianza acumulada fue del 99,9 %. Los altos valores de nitratos observados en La Victoria se asociaron con la gran abundancia de Metrichia, Ochrotrichia y Leucotrichia. En Puerto Mosquito la gran abundancia de Smicridea, Zumatrichia y Nectopsyche se relacionó con los altos valores de fosfatos y conductividad, en tanto que la mayor abundancia de Polycentropus y Cyrnellus se presentó en San Lorenzo, y La Cascada, con los mayores valores de oxígeno disuelto y nitritos, se caracterizó por la presencia de cf. Amazonatolica (Figura 6).

Discusión
En este estudio se observó que la estructura del río Gaira es la típica de los ríos tropicales de alta montaña, con una notable variación ecosistémica a lo largo de su gradiente alti-tudinal, similar a los ríos estudiados por Jaimes & Granados (2016), Serna, et al. (2015) y Rúa, et al. (2015), y una abundante vegetación ribereña en sus zonas más altas. A pesar de ser diferentes ríos, hacen parte de la Sierra Nevada de Santa Marta (SNSM) y mantienen el mismo patrón altitudinal en lo esencial. Los ríos estudiados en otras regiones del país (Lopez-Delgado, et al., 2015; Ballesteros, et al, 1997; Guevara-Cardona, et al, 2007; Posada-García & Roldán-Pérez, 2003; Zúñiga, et al, 1993), por ejemplo los de las zonas andinas, presentan un patrón altitudinal diferente, suelen ser caudalosos en gran parte de su cauce, de mayor tamaño y con pendientes más inclinadas que provocan corrientes de agua fuertes, tramos de rápidos y cascadas que contribuyen a la retención de oxígeno, a diferencia de los ríos que han sido estudiados en la Sierra Nevada de Santa Marta, donde las pendientes más inclinadas se encuentran en sus partes más altas y son más horizontales a lo largo de su cauce, por lo que se aprecian más sitios de remansos, así como fluctuación del factor antròpico. Sin embargo, la acumulación del material orgánico a lo largo del cauce, que corre hacia las partes bajas, da pie a la formación de ríos heterotróficos que favorecen el establecimiento de gran diversidad de organismos (Vannote, et al, 1980).
Según Vásquez-Ramos, et al. (2010, 2014) existen organismos que se han adaptado a vivir en tramos específicos de los ríos, en este caso, cf. Amazonatolìca, Cyrnellus, Nectopsyche y Polycentropus, taxones exclusivos de las partes altas del río (San Lorenzo y La Cascada), donde las aguas son muy oxigenadas, las temperaturas bajas y la contaminación de origen antrópico, poca. En la parte media del río fueron Ochotrichia, Cerasmatrichia y Metrichia, tramo donde se presentó la menor abundancia probablemente porque en esta zona existen grandes extensiones de cultivos de café y otros cultivos transitorios como maíz y frutales, los cuales pueden superar las 300 ha, y en algunos sectores se desarrollan actividades ganaderas de baja intensidad. En la estación ubicada en la parte baja (Puerto Mosquito) se encontraron los taxones exclusivos Protoptila, Mortoniella, Oecetis y Chimarra, además de la mayor abundancia y diversidad de géneros. Este fenómeno se sustenta en lo expuesto por Vannote, et al. (1980) en el sentido de que las partículas finas de materia producto de la fragmentación resultante de los procesos físicos, químicos y biológicos que terminan en la parte baja del rio, así como la luz que promueve la proliferación de algas y la alta carga de minerales acumulados a lo largo del cauce, generan microhábitats propicios para la proliferación y diversidad de tricópteros.
En cuanto a los atributos ambientales, se logró establecer su relación con la abundancia de los géneros. El pH se mantuvo dentro del rango de la neutralidad (7,28 - 7,48), una de las características propias de los ecosistemas oligotróficos (Roldán, 1992), como es el caso de algunos ríos de la Sierra Nevada de Santa Marta (ProSierra, 1998). El oxígeno disuelto presentó valores entre 7,5 y 8,5 mg/L, lo que favorece el asentamiento y desarrollo de una gran densidad biológica de entomofauna acuática debido a la significativa oxigenación de los ríos (Guerrero, et al., 2003). Por otra parte, los nitratos, nitritos, fosfatos y la conductividad registraron diferencias según el gradiente altitudinal, lo que se reflejó en las zonas media y baja del río. El incremento de estas variables podría explicarse por la moderada actividad agropecuaria en proximidades de la estación Puerto Mosquito.
De los 53 géneros de tricópteros registrados para Colombia (García & Moreno, 2000; Muñoz-Quesada, 2000; Grimaldo, 2001; Fuentes & Carmona, 2001; Manjarrés & Manjarrés-Pinzon, 2004; Muñoz-Quesada, 2004; López-Delgado, et al., 2015), 20 se encuentran en el río Gaira. Se reporta aquí por primera vez cf. Amazonatolica para la región y para el país, por lo que el número de géneros asciende a 54 y sugiere que existe una gran diversidad del orden en un solo río de la Sierra Nevada de Santa Marta. En este estudio, la riqueza genérica fue mayor a la registrada por Serna, et al. (2015), quienes registraron 14 géneros en un gradiente altitudinal entre los 60 y 360 m del río Manzanares (río vecino del Gaira), y la reportada por Rúa, et al. (2015), quienes registran nueve géneros en un rango altitudinal de 0 a 900 m en el río Gaira. Uno de los factores determinantes de estas diferencias podría estar relacionado con el gradiente altitudinal más amplio considerado en el presente estudio, entre los 65 y los 2.000 m s.n.m.
La parte baja del río (Puerto Mosquito) fue la de mayor número de taxones, una gran diversidad comparada con los tramos medio y alto (Anexo B,https://www.raccefyn.co/index.php/raccefyn/article/view/1148/2754). Moreno, et al. (2011), lo que confirma que el uso del método de los números efectivos de especie promueve una mejor comprensión a la hora de interpretar la diversidad de las comunidades debido a que se unifican diferentes componentes que permiten establecer comparaciones entre sitios diferentes. La Victoria (parte media) presentó el mayor número de taxones en abundancia similar, lo que evidencia que la abundancia se repartió de forma similar entre los géneros y podría estar asociado con características propias de zonas intermedias, que usualmente poseen una gran diversidad y comparten taxones en la cabecera y la parte baja de los ríos (González & García, 1984; Posada & Roldán, 2003; Rivera, et al., 2008).
La gran abundancia y distribución de la familia Hydropsychidae se debe a que su historia natural exhibe un amplio rango de distribución, ya que es una de las familias más diversas de los tricópteros del Neotrópico, con cerca de 500 especies (Holzenthal & Calor, 2017), y tiene la habilidad de colonizar diferentes tipos de sustratos como rocas, arena, grava, cieno y hojarasca (López, et al., 2015; Vásquez, et al., 2008). Otro factor está asociado con el hecho de que los géneros Smicridea y Leptonema pueden tolerar cambios en las condiciones ambientales y construir redes extensas de seda que les permiten fijarse al sustrato y capturar gran cantidad de alimento (García, et al., 2009; Vásquez-Ramos, et al., 2010; Zúñiga, et al., 2013; López-Delgado, et al., 2015).
Por otra parte, la poca abundancia de Helicopsychidae y Glossosomatidae coincide con lo hallado en otros estudios en ríos andinos (Guevara, et al., 2005, Vásquez-Ramos, et al., 2010), lo que se explica por el hecho de que estas larvas habitan sustratos específicos. La gran riqueza de las familias Hydroptilidae y Leptoceridae fue similar a la registrada por Rúa, et al. (2015), lo que se debe a que son grupos con un alto número de géneros y especies a nivel global (Holzenthal & Calor, 2017) y sus géneros son capaces de colonizar una gran diversidad de sustratos debido a sus diversos comportamientos tróficos (Posada & Roldán, 2003). Su amplio rango de distribución puede explicarse porque los géneros de estas familias están asociados con ríos y quebradas de diferentes corrientes y temperaturas (Rincón, 1999), por su gran tolerancia a cambios en las condiciones ambientales y su alimentación con perifiton, en especial las diatomeas, que proliferan en las rocas de las zonas de salpicadura y tienen la capacidad de construir refugios en forma cónica que les brindan protección y los hacen poco visibles a los depredadores (Correa, et al., 1981; Ballesteros, et al., 1997; Muñoz-Quesada, 2000; Posada, et al., 2000; Muñoz-Quesada, 2004; Guevara-Cardona, et al., 2007; García, et al., 2009; Vásquez-Ramos, et al., 2010; López-Delgado, et al., 2015).
En La Victoria (parte media) y Puerto Mosquito (parte baja) se encontró la mayor riqueza, en contraste con La Cascada y San Lorenzo (partes altas). Según Townsend & Hildrew (1994) y Rosenzweig (1995) son muchos los factores que pueden influir en la abundancia y la riqueza de taxones, incluidos la diversidad, la heterogeneidad del hábitat y el gradiente altitudinal. En estudios como los de Ballesteros, et al. (1997), Guevara-Cardona, et al. (2007), Posada-García & Roldán-Pérez, (2003), Zúñiga, et al. (1993) en rangos altitudinales de la región Andina similares a los del presente trabajo, se ha evidenciado una tendencia notoria a una mayor abundancia y riqueza de géneros en las cuencas de mayor tamaño y caudal, lo cual es consistente con una mayor disponibilidad de microhábitats que favorecen la diversidad.
Conclusión
La familia Hydropsychidae fue la más abundante y se encontró en cada sitio de muestreo, con dos géneros solamente: Leptonema y Smicridea, cuyas larvas fueron las más abundantes en el río Gaira (40,6 %) y las de mayor rango de distribución altitudinal, lo que evidencia su capacidad para colonizar diferentes tipos de sustratos y tolerar ambientes diversos.
A pesar de que la estructura de los ensamblajes de tricópteros fue similar en la mayoría de los tramos, se encontró un gran número de taxones exclusivos en el tramo bajo (Puerto Mosquito), el medio (La Victoria) y el alto (San Lorenzo). Esto evidencia que algunos géneros tienen limitada distribución y deben estudiarse con base en su autoecología. Además, la gran diversidad de tricópteros en el Gaira se refleja en el elevado número de géneros (21) encontrados en un solo río, de los cuales 11 son nuevos registros para el río y uno lo es para Colombia (cf. Amazonatolica).

