











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
Una de las principales consecuencias negativas de las actividades agropecuarias sobre los arroyos o quebradas de cabecera (1-2 orden) en la zona andina colombiana es la eliminación de los bosques ribereños (Chará, et al., 2007), los cuales cumplen funciones de gran importancia para la protección de los ambientes acuáticos. Los bosques ribereños reducen el deterioro de la calidad del agua a través de la filtración de sedimentos, pesticidas, nutrientes y otros elementos que provienen del área de captación de las microcuencas (Schilling & Jacobson, 2014; Sklodowski, et al., 2014) y contribuyen a reducir la erosión de los taludes y a mantener estable la temperatura del agua (Naiman, 2005; Rood, et al., 2014). Además, estos elementos del paisaje son reservorios importantes de biodiversidad terrestre, pues sirven de sitio de refugio, tránsito o reproducción a especies de aves, anfibios y mamíferos, entre otros (Saalfeld, et al., 2012; Tanaka, et al., 2016).
En paisajes ganaderos, cuando el bosque ribereño desaparece completamente, las gramíneas exóticas invaden la zona ribereña y el cauce, lo que constituye una barrera agresiva para la regeneración de especies leñosas y los procesos de restauración de la vegetación nativa en estas áreas (Richardson, et al., 2007; Galindo, et al., 2017). Esta situación, sumada a la actividad de los bovinos en los potreros y directamente en el cauce, provoca que la complejidad de los ambientes acuáticos en términos biofísicos se reduzca en gran medida, al igual que la calidad del agua y la biodiversidad acuática (Belsky, et al., 1999; Sovell, et al., 2000; Sweeney & Czapka, 2004; Braccia & Voshell, 2007).
Aunque el impacto de las actividades ganaderas en las fuentes de agua se ha estudiado a nivel mundial y se han demostrado sus consecuencias negativas (Piscart, et al., 2009; Mesa, 2010; Riseng, et al., 2011; Ramírez, et al., 2018), poco se conoce sobre el efecto que tiene la exclusión del ganado de la zona ribereña y los efectos iniciales de la restauración de los bosques en las comunidades biológicas y en la calidad de agua. En algunos estudios realizados en la zona cafetera colombiana se ha demostrado que el solo hecho de impedir el acceso del ganado a los cauces mejora la calidad del agua y del hábitat y provoca un cambio de las comunidades de macroinvertebrados en un tiempo relativamente corto, aunque esta recuperación es limitada si la rehabilitación de la cobertura arbórea es baja (Pedraza, et al., 2008; Chará, et al., 2011).
Según la 50, la restauración ecológica es el proceso de propiciar el restablecimiento de un ecosistema que se ha degradado, dañado o destruido como resultado directo o indirecto de las actividades humanas. Para el caso de los arroyos y ríos, la restauración de los bosques ribereños se considera una de las acciones prioritarias que repercute de manera positiva en el mejoramiento de las condiciones del cauce (Herbst, et al., 2012; Kuglerová, et al., 2014; de Mello, et al., 2017). Algunas características en el ambiente acuático pueden responder a la restauración, aunque es necesario evaluarlas por un periodo relativamente largo para notar los cambios que se producen (Herbst, et al., 2012).
En la cuenca media del río La Vieja, Colombia, a través de diferentes proyectos, algunos productores ganaderos han aceptado que las áreas aledañas a los cursos de agua sean separadas de la producción ganadera y se permita la recuperación de la vegetación nativa (Calle & Holl, 2019). Sin embargo, en la zona ribereña en la que predominan diversos géneros de gramíneas africanas como Cynodon, Penissetum, Echinochloa y Urochloa, entre otros, se presenta un crecimiento exagerado de las mismas, lo que impide el establecimiento de especies arbóreas (sembradas o que surgen por regeneración natural) y, por lo tanto, la recuperación del bosque es muy lenta o no puede lograrse fácilmente (Galindo, et al., 2017).
Para reducir el crecimiento de gramíneas en los corredores ribereños, se ha propuesto la siembra de plantas acompañantes, o nodriza, de crecimiento rápido que puedan competir con los pastos y, a la vez, permitir el crecimiento de especies arbóreas (Perkins, et al., 2014; Medeiros, et al., 2014). El presente estudio muestra los cambios ocurridos en la calidad del agua, en las características del hábitat y en la comunidad de macroinvertebrados acuáticos en quebradas sometidas a una estrategia de restauración de la vegetación ribereña a partir de plantas acompañantes de rápido crecimiento en una zona ganadera del Eje Cafetero de Colombia.
Materiales y métodos
Área de estudio
El estudio se llevó a cabo en cuatro quebradas de primer orden ubicadas en fincas ganaderas de la cuenca media del río la Vieja, en el centro-occidente de Colombia. Tres de las quebradas se localizan en el municipio de Alcalá, Valle del Cauca (quebrada 1: finca Palo Coposo, 4°39'32''N, 75°44'42'O; quebrada 2: finca El Rubí, 4°41'77''N, 75°47'59"O; quebrada 3: finca Villa Adriana, 4°41'24''N, 75°48'22''O) y un sitio en el municipio de Quimbaya, Quindío (quebrada 4: finca Tierra Labrantía, 4°38'59''N, 75°48'21''O), todas a alturas entre los 1.080 y 1.370 m s.n.m.
Para la selección de las fuentes de agua se tuvo en cuenta que el uso del suelo en el área de captación de las microcuencas tuviera predominio de ganadería bovina y que no fuera afectada por ningún tipo de contaminación puntual, como descargas de aguas residuales de origen doméstico o agropecuario. La zona ribereña estaba dominada por gramíneas exóticas de la familia Poaceae como Cenchrus purpureus (Schumach.) Morrone 2010; Paspalum paniculatum L, 1759; Megathyrsus maximus (Jacq.) B.K. Simon & S.W.L. Jacobs 1781; y Cynodon plectostachyus (K. Schum) Pilg. 1907 (Galindo, et al., 2017).
Métodos
En cada quebrada se seleccionó un tramo de 45 metros de longitud y se construyó una cerca al lado y lado del cauce, a una distancia de tres metros de cada orilla para evitar la entrada de ganado al ambiente en proceso de restauración. En esta área se sembraron especies de arbustos de rápido crecimiento pioneros en la zona (Tithonia diversifolia y Piper auritum) para inhibir el crecimiento de las gramíneas en el área ribereña, así como especies de árboles nativos propios de bosques ribereños de la región, entre ellos Inga edulis, Ampelocera albertiae, Brosimum alicastrum, Anacardium excelsum, Syagrus sancona y Aiphanes horrida (Galindo, et al., 2017).
Durante tres años (2010, 2011, 2012), con dos evaluaciones cada año, se tomaron muestras puntuales de agua para análisis fisicoquímico y bacteriológico que incluyeron los parámetros de alcalinidad, conductividad, pH, demanda bioquímica de oxígeno, dureza, fósforo total, nitratos, nitrógeno amoniacal, nitrógeno total, oxígeno disuelto, sólidos suspendidos totales, turbiedad, y coliformes fecales y totales.
Para evaluar la condición del hábitat en el tramo, se utilizó la metodología descrita por Barbour, et al., (1999), adaptada por Chará (2004), que permite obtener el índice de calidad de hábitat (ICH) mediante la sumatoria de valores dados a diferentes variables en el tramo y algunos aspectos de la microcuenca como el sustrato disponible para la colonización de organismos, la colmatación de espacios entre piedras, la caracterización del sustrato en piscinas, la combinación de velocidad y profundidad, la diversidad de piscinas, la deposición de sedimentos, el estado del nivel del agua, la perturbación del cauce, la frecuencia de turbulencias, la sinuosidad del cauce, la estabilidad de las orillas, la protección vegetal en orillas, y el ancho del bosque ribereño. A cada variable se le asignó un puntaje entre 0 (cero), es decir, el estado más afectado, y 20, que supone un estado óptimo.
Por otro lado, en cinco puntos del tramo se caracterizaron de manera visual algunos aspectos del hábitat, como la composición relativa del sustrato (piedras, grava, gravilla, arena, lodo) y los patrones de flujo (turbulencias, charcas, y tramos con corriente lenta y corriente rápida). Además, en los mismos puntos se midió el ancho y la profundidad del cauce.
Después de tomar las muestras fisicoquímicas y caracterizar el hábitat, se recolectaron macroinvertebrados acuáticos utilizando una red Surber de 0,09 m2 (500 |im de ojo de malla). En cada seguimiento se tomaron nueve muestras a lo largo del tramo en los microhábitats presentes. Las muestras de macroinvertebrados se preservaron en campo con alcohol etílico al 80 % y, posteriormente, se separaron e identificaron en el laboratorio hasta el menor nivel taxonómico posible con la ayuda de un estereoscopio (Nikon®, Modelo SMZ-645) con aumentos entre 10 y 40X y claves taxonómicas para los diferentes grupos (Posada & Roldán, 2003; Springer, 2006; Heckman, 2006; Domínguez & Fernández, 2009; Fiuza, 2012; Prat, 2012).
Análisis de datos
Las diferencias del promedio de las variables fisicoquímicas, del ICH, las características del hábitat, y la abundancia y diversidad de macroinvertebrados acuáticos entre los años de muestreo se determinaron mediante un análisis de varianza (ANOVA) de dos vías. Sin embargo, las variables que no presentaron normalidad se analizaron con una prueba estadística de tipo no paramétrico (Kruskal-Wallis). Estos análisis se hicieron con el programa Statistica 7.0 (StatSoft, Inc., 1984-2004).
La diversidad de macroinvertebrados entre los años de muestreo se calculó utilizando los números de Hill (Chao, et al., 2014), que tienen en cuenta el número efectivo de especies (Moreno, et al., 2011). En este caso se usaron tres valores 0 D (riqueza de especies), 1 D (diversidad de Shannon) y 2 D (diversidad de Simpson). Para la estimación y la comparación de q D se usó el mismo nivel de cobertura de muestreo entre comunidades (Chao, et al., 2014); la diversidad se calculó usando 100 aleatorizaciones con intervalos de confianza del 95 % en el programa SPADE (Chao, et al., 2010). Además, se estimó el índice biótico BMWP-Univalle, adaptado por Zúñiga & Cardona (2009) a condiciones regionales y cuerpos de agua del suroccidente colombiano y el índice de abundancia relativa de EPT (Ephemeroptera, Plecoptera y Trichoptera), según lo propuesto por Carrera & Fierro (2001).
Para evaluar la asociación de la comunidad de macroinvertebrados en las diferentes quebradas a lo largo del tiempo, se usó un análisis de escalamiento multidimensional no métrico (NMDS) y se probó el nivel de significación de los grupos formados mediante un análisis de similitud (ANOSIM). Estos análisis se hicieron con el programa Primer-E, versión 6.
Para determinar las asociaciones estadísticas entre la abundancia de cada género de macroinvertebrados y las variables ambientales, se hizo un análisis de correspondencia canónica (CCA: largo del gradiente > 4 desviaciones estándar), previa transformación a raíz cuadrada de la abundancia de macroinvertebrados. Las variables ambientales se escogieron (p<0,05) usando selección manual y 9.999 iteraciones de Monte Carlo. Estos análisis se realizaron con el programa CANOCO (TerBraak & Smilauer, 2002).
Para evaluar las relaciones existentes entre las variables fisicoquímicas y las variables de la comunidad de macroinvertebrados que presentaron cambios significativos a lo largo de los años de monitoreo, se hizo una correlación de Pearson usando el programa estadístico PAST, versión 3.21.
Resultados
Parámetros fisicoquímicas y bacteriológicos
En general, los parámetros de las fuentes de agua presentaron valores dentro del rango normal para aguas corrientes con impacto de la ganadería en la zona de estudio (Tabla 1). En los tres años de seguimiento, la DBO5 disminuyó significativamente, casi en un 80 % (F26=6,07, p<0,05), en tanto que el nitrógeno amoniacal aumentó significativamente (F26=17,08, p<0,05). Por otro lado, hubo un aumento leve de los valores del oxígeno disuelto y una disminución en la turbiedad y los coliformes fecales y totales a través del tiempo.

Características del hábitat
Los cambios en las variables correspondientes a las características físicas del hábitat, no mostraron diferencias estadísticas significativas a lo largo de los años de seguimiento. Sin embargo, se notó una recuperación leve de algunos parámetros relacionados con los sustratos y los tipos de flujo reflejada en la aparición de grava y corriente rápida en el año 2011 y el 2012 comparados con el 2010, cuando se presentaba solo un tipo de sustrato (lodo) y corriente laminar. Además, el ICH mostró aumento con el tiempo (Tabla 2).

Macroinvertebrados acuáticos
Durante los tres años de estudio se recolectaron 10.490 individuos distribuidos en 19 órdenes, 53 familias y 56 géneros. Insecta fue la clase más representativa (84 %), seguida por Gastropoda (9 %). El orden más abundante fue Diptera (56 %), seguido por Trichoptera (22 %) y Basommatophora (7 %). En cuanto a las familias, Chironomidae e Hydropsychidae fueron las más abundantes con el 53 y el 20 % de abundancia total, respectivamente.
Chironomidae mostró diferencias significativas entre los años de monitoreo (F26=14,73, p<0,05). Esta familia fue muy abundante en el primer año de evaluación (promedio 81,89 ± 8,71%) y disminuyó en el 2011 (promedio 32,63 ± 24,45%) y el 2012 (promedio 28,68 ± 25,61%) (Figura 1). Aunque no fue significativo el cambio en la abundancia de la familia Hydropsychidae (F29=1,2, p=0,21), hubo un aumento en el número de individuos entre el 2010 y el 2012 (Figura 1).

Figura 1 Cambios en la abundancia relativa de las familias Chironomidae e Hydropsychidae durante la restauración temprana del corredor ribereño en quebradas del Eje Cafetero de Colombia. El punto representa el valor promedio; la línea representa la desviación estándar
La mayor riqueza de macroinvertebrados se registró durante el segundo año del proceso de restauración (2011) (Figura 2B), en tanto que la abundancia de individuos mostró una tendencia a disminuir en el segundo año y de nuevo aumentó en el tercero (2012) (Figura 2A). En cuanto al índice BMWP-Univalle (Figura 2E) y el porcentaje de Ephemeroptera, Plecoptera y Trichoptera- EPT (Figura 2F), aumentaron significativamente entre el primer año (2010) y el tercero (2012) del proceso.

Figura 2 Valores promedio en variables de la comunidad de macroinvertebrados A. Abundancia total. B. Riqueza de géneros. C. Diversidad q1. D. Diversidad q2. E. Índice BMWP-Univalle. F. Porcentaje de Ephemeroptera, Plecoptera y Trichoptera (EPT). Las líneas sobre las barras representan la desviación estándar
Los resultados del NMDS (stress=0,11) y el ANOSIM (R=0,52, p<0,001) fueron moderados en términos de formación de grupos entre años. Se obtuvieron dos grupos bien definidos (Figura 3): el primero comprendió la comunidad de macroinvertebrados muestreados en las cuatro quebradas en el primer año de restauración, y el segundo, la comunidad muestreada en las cuatro quebradas durante el segundo y el tercer año del proceso de restauración.

Figura 3 Ordenación del análisis de escalamiento multidimensional no métrico (NMDS) de la comunidad de macroinvertebrados registrados en las quebradas con procesos de restauración del corredor ribereño. El Rubí (R), Palo Coposo (P), Tierra Labrantía (T) y Villa Adriana (V) durante los años 2010, 2011 y 2012. Óvalo línea continua: primer año. Óvalo línea punteada: segundo y tercer años del proceso de restauración
El análisis de correspondencia canónica (CCA) explicó el 28,55 % de la abundancia de géneros de macroinvertebrados en las cuatro quebradas a lo largo del tiempo. Las variables de año y nitrógeno total explicaron significativamente (p<0,05) el 17,05 % y el 11,50 %, respectivamente, de la varianza de la comunidad de macroinvertebrados (Figura 4). Se observó la formación de tres grupos correspondientes a cada año de muestreo, entre los cuales uno de ellos se separó de los otros principalmente por el contenido de nitrógeno total en el agua.
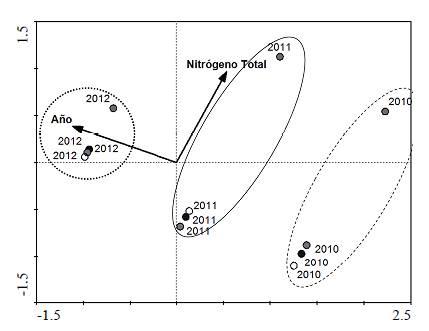
Figura 4 Análisis de correspondencia canónica para la comunidad de macroinvertebrados con variables ambientales (flechas) y quebradas (círculos). Círculo blanco: Tierra Labrantía; círculo gris claro: Palo Coloso; círculo negro: El Rubí; círculo gris oscuro: Villa Adriana
El análisis de correlación de Pearson evidenció una correlación positiva entre el porcentaje de Chironornidae y la DBO5 (R=0,63, p<0,05).
Discusión
Los resultados de este estudio demuestran que, en un tiempo relativamente corto, la recuperación de la vegetación ribereña ejerció un efecto muy importante en el mejoramiento de la calidad del agua, principalmente en la reducción significativa de los valores de DBO5, la disminución de la turbiedad y de los coliformes, y el aumento del oxígeno disuelto, lo que se reflejó en el cambio de la composición de la comunidad de macroinvertebrados acuáticos.
Estos cambios son relevantes a nivel regional, debido a que en paisajes fragmentados por actividades agropecuarias, los bosques ribereños son los únicos relictos que pueden quedar y contribuyen a la protección de microcuencas (Ericsson & Stevens, 1996; Tanaka, et al., 2016) que prestan servicios ambientales de gran importancia en la red hídrica de la cual hacen parte (Palmer, et al., 2014). Además, su recuperación puede contribuir a los compromisos de restauración de paisajes rurales latinoamericanos (Meli, et al., 2019).
Es conocido que la cobertura vegetal ribereña tiene un efecto positivo sobre diferentes variables relacionadas con la calidad del agua y la biodiversidad acuática (Ding, et al., 2013; Kuglerová, et al., 2014; Ou, et al., 2016; Yang, et al., 2016). En el caso de este estudio, las respuestas en el ambiente acuático están dadas en gran medida por el efecto de los arbustos de rápido crecimiento establecidos en la zona ribereña que, a su vez, disminuyeron la cobertura de las gramíneas. En el tiempo de estudio Tithonia diversifolia y Piper auritum lograron tener una cobertura entre el 40 y el 80 %, lo que inhibió notablemente el crecimiento de las pasturas, en especial T. diversifolia. Además, estas especies contribuyeron a la supervivencia de árboles de sucesión intermedia a tardía plantados como parte del proceso de restauración (Galindo, et al., 2017).
A pesar de que en tres años de estudio la vegetación ribereña establecida no logró la composición y la estructura de especies de bosques avanzados, se demostró que los arbustos usados para disminuir el crecimiento de las pasturas prestan servicios importantes de protección del ambiente acuático. Calle & Holl (2019) encontraron en corredores ribereños separados de la producción ganadera entre 10 a 14 años en la misma zona de estudio (cuenca media del río La Vieja), una proporción similar de especies leñosas que en los bosques de referencia, además de la recuperación de la estructura vegetal, alta cobertura del dosel y baja cobertura de hierbas. Se espera, entonces, que con un mayor tiempo de recuperación, los corredores ribereños estudiados logren tener una complejidad estruc-tural similar a sitios de referencia de la zona.
Aunque los resultados evidencian el mejoramiento de varios parámetros de la calidad de agua, la poca reducción, o incluso el incremento de los valores de parámetros como el nitrógeno amoniacal, puede estar asociada al uso de fertilizantes en el área de captación de las microcuencas, ya que en los tres años de estudio los suelos aledaños de algunas quebradas pasaron de pasturas a sistemas agrícolas, lo que implica mayor uso de fertilizantes químicos en zonas de altas pendientes. Otros autores han encontrado pocos cambios en la concentración de nutrientes en ambientes con procesos pasivos de restauración (McKergow, et al., 2003; Hughes & Quinn, 2014; Muller, et al., 2016).
Sin embargo, los resultados mencionados contradicen los encontrados por de Mello, et al. (2017), quienes reportan una reducción importante del nitrógeno (22,6 %) en cuencas con procesos de restauración en Brasil. Herbst, et al. (2012) consideran que las quebradas pueden recobrar su salud ecológica, pero depende de varios factores como el uso del suelo en el área de captación y la reducción de la entrada de sedimentos al cauce. La falta de consenso en los estudios demuestra que la respuesta de cada sitio depende de varios factores particulares que intervienen en los procesos de recuperación del ambiente y que no todos los proyectos responden de igual manera (Teels, et al., 2006, Miller, et al., 2009), se requiere de más información para entender estas dinámicas, entre las que se encuentran el ancho mínimo y la densidad vegetal de los corredores ribereños para impedir la entrada de nutrientes al agua producto de la aplicación de fertilizantes en los terrenos aledaños a las quebradas, por ejemplo.
La situación en cuanto a características del hábitat como los sustratos y los tipos de corriente es similar a la de otros estudios realizados en la zona, en los que durante un tiempo similar de evaluación no hubo cambios significativos en la composición de estos elementos en el cauce (Pedraza, et al., 2008; Chará, et al., 2011). Los ambientes acuáticos con impacto directo de la ganadería pierden un alto porcentaje de los sustratos gruesos, como bloques, cantos y guijarros, debido al incremento de la erosión de los bancos y la colmatación del cauce con sedimentos (Riseng, et al., 2011; Giraldo, et al., 2014; Ramírez, et al, 2018). Se podría inferir que con la exclusión del ganado de la zona ribereña y un tiempo más prolongado de recuperación de la vegetación, las quebradas podrían recuperar dichas condiciones dentro del cauce, ya que son características determinantes para el establecimiento de la fauna acuática (Richards & Host, 1994; Skuja & Spungis, 2010).
Los resultados de este estudio evidenciaron que la composición de macroinvertebrados acuáticos era diferente en el primer año de monitoreo comparada con los siguientes según la formación de los grupos diferenciados (Figura 3). Esta condición está relacionada con el crecimiento de vegetación leñosa en el área ribereña, el mejoramiento de las condiciones del hábitat, tal como lo demuestra la calificación del ICH, y el mejoramiento de parámetros como el oxígeno disuelto y la menor concentración de materia orgánica y sedimentos, lo cual propicia condiciones para la colonización de nuevos organismos acuáticos asociados con ambientes menos perturbados.
Así como para algunas características del hábitat y algunos parámetros fisicoquímicos, también para los macroinverteibrados acuáticos se han presentado resultados variables en otros estudios. Por ejemplo, Louhi, et al. (2011) registraron pocos cambios en la comunidad de macroinvertebrados acuáticos a pesar de la recuperación de hábitats en el cauce, en tanto que Westveer, et al. (2018) encontraron una rápida colonización del ambiente y el aumento de la riqueza de macroinvertebrados en sitios restaurados.
La abundancia de Chironomidae se redujo significativamente a medida que avanzó el proceso de restauración. Esta familia comúnmente se asocia con ambientes perturbados y es muy abundante en sitos con altas concentraciones de materia orgánica (Barbour, et aL, 1999; Chará, et al., 2009). Su reducción se correlacionó con la disminución en la concentración de DBO5, que es el principal indicador de contaminación orgánica en estos cauces.
Por otro lado, aunque no fue significativo el cambio, la familia Hydropsychidae mostró una tendencia al aumento a través del tiempo. Además, aparecieron otras familias del orden Trichoptera como Helicopsychidae, Leptoceridae y Philopotamidae, lo que evidencia que hay condiciones en las quebradas para el establecimiento de este grupo, considerado sensible a la perturbación del hábitat (Bonada, et al., 2006; Chakona, et al., 2009).
La sumatoria de Ephemeroptera, Plecoptera y Trichoptera (EPT) es un índice biótico comúnmente empleado como indicador biológico debido a que los tres órdenes que lo componen se consideran sensibles a las perturbaciones del ambiente (Rosemberg & Resh, 1996; Gamboa, et al., 2008). En el presente estudio se evidenció un aumento significativo en la importancia relativa de estos tres órdenes al pasar de 1,9 a 24,5 % del total de los organismos recolectados. Este patrón es similar a lo reportado por Herbst, et al. (2012), quienes encontraron un cambio significativo en la riqueza de EPT después de cuatro años de impedir el pastoreo de ganado en pequeñas quebradas en Estados Unidos y sus zonas de captación, cambio relacionado con una mejora en la cobertura ribereña y las características del cauce. Según estos resultados, EPT podría proponerse como un indicador de la recuperación de ambientes acuáticos en proceso de restauración, pues, además, su recolección e identificación a nivel de familia son relativamente fáciles.
Dado que la conservación de los bosques y el agua debería ser una prioridad en paisajes agropecuarios, la restauración de los corredores ribereños constituye una de las estrategias más efectivas en este objetivo. Se demostró que una de las estrategias para iniciar estos procesos en zonas ganaderas es el establecimiento de arbustos de rápido crecimiento que propicien condiciones para la regeneración natural de la vegetación en la zona ribereña, lo que a su vez tiene efectos positivos sobre el ambiente acuático. Sin embargo, es muy importante que estas iniciativas estén acompañadas del compromiso de los productores, que son quienes finalmente inciden de manera concreta en el manejo y el cambio de uso de la tierra.
Conclusiones
Los resultados de este estudio demuestran que la exclusión de la ganadería del área ribereña y el inicio de procesos de restauración de la vegetación contribuyen a la recuperación de la calidad del agua y la comunidad de macroinvertebrados acuáticos. Se redujeron principalmente taxones asociados con ambientes perturbados y aparecieron grupos más sensibles, a pesar de que la recuperación de aspectos relacionados con características del hábitat como la diversidad de los tipos de sustratos y los patrones de flujo fue leve en tres años.
Es posible recuperar ambientes acuáticos con procesos de restauración de la vegetación ribereña mediante la siembra de arbustos de rápido crecimiento. Esto es especialmente útil en zonas aledañas al cauce invadidas por pasturas y que tienen dificultades para la conformación de un bosque ribereño.

