











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
The continuous increase in demand for food requires intensification of land use systems in the agricultural sector and achieving a compromise between increases in production and ecological conservation constitutes an important challenge (Davies et al. 2010; van Vuuren and Chilibroste 2013). Additionally, climate change may further increase the demands on management of resources and the capacity of livestock farmers and their rural communities to adapt (Herrero et al. 2017).
The watershed of the Orinoco River covers 35 Mha in Colombia (33% of the land area) and includes mountains, foothills and a variety of seasonally flooded and well drained savannas. The well drained savannas ("Altillanura") extend over 13.5 Mha (CONPES 2014), 35% of which is plain ("Plains") and 54% is slopes and hills (Rippstein et al. 2001). The latter include small valleys suitable for cropping, surrounded by pronounced slopes that have shallow, stony soils. By 2007, 23% of the Plains had been converted to crops, sown pastures, palm oil plantations (Romero-Ruiz et al. 2012; Rausch 2013) and a variety of tree plantations and reforested areas amounting to close to 100,000 ha (MADR 2015). Palm oil plantations have experienced a significant and ongoing increase in the savannas, and have reached 112,186 ha (FEDEPALMA 2015), including large areas in the Andean foothills. Overall, cropped areas represent 4.311.8%, whereas rangelands cover 72-89% of the Orinoco basin in Colombia (DANE 2016).
This expansion of crops and plantations has taken place at the expense of native savannas supporting beef cattle ranching. On areas closest to roads in the Plains, beef breeding herds have been replaced by yearlings brought in from the surrounding area for fattening, interspersed with crops and plantations (Romero-Ruiz et al. 2012; Huertas-Ramírez and Huertas-Herrera 2015). The cattle population of the region is estimated at 4.7 M head (DANE 2016), largely supported by native savannas that still constitute the main land use in the rest of the area, mainly dedicated to extensive beef breeding herds (Rausch 2013; Huertas-Ramírez and Huertas-Herrera 2015). The latter are low input-low output systems, frequently constrained by limited access, physiographic and water limitations, low soil fertility and often shallow soils that limit intensification (Seré and Vera 1983). On the other hand, these savannas are rich in plant and animal biodiversity (Rippstein et al. 2001; Lasso et al. 2011) and have varied landscapes and ecosystem services that attract rural tourism highlighted by educational and cultural values and traditions (Navas Ríos 1999; Molina and Triana 2011; Australian Government 2015).
Although the real impact of greenhouse gas emissions from cattle on neotropical savannas is still a challenge for scientists, it is likely that plant dynamics may mitigate the demanding effects of climate variability, reflecting the capacity of these plant-animal evolutionary systems to adapt to genetic, environmental and management stressors (O'Neill et al. 2010; Herrero et al. 2015; Ramírez-Restrepo and Charmley 2015). In this complex scenario, primary producers have to contend with low seasonal biomass production and nutritive value (Paladines and Leal 1979; Rippstein et al. 1996; Durmic et al. 2017). Together, these factors negatively impact productivity in terms of slow growth and fertility rates that seldom exceed 50%, yielding no more than 3 or 4 calves weaned over a cow's lifetime (Kleinheisterkamp and Habich 1985; Plessow 1985; Squires and Vera 1992).
These traits contrast with the relatively high performance reported when Brahman (Bos indicus) beef cows are grazed year-round on well managed tropical sown pastures (Vera et al. 2002). Nevertheless, expensive sown pastures are mostly reserved for fattening yearlings and steers (Vera and Seré 1985), and their year-round use by the breeding herd may represent an economically suboptimal use of an expensive resource. It is possible that strategic and seasonal grazing of improved pastures by suckling cows to complement native savanna grazing may significantly increase reproductive indexes (Vera and Seré 1989).
Reproductive rates may be boosted further by early weaning of calves, a technology that is 50 years old. In principle, early weaning can be performed at 45 days of age (Rasby 2007) and it is particularly useful in drought situations and to contend with the negative effects of climate change (FAO 2013). Cow-calf research in the savannas of northern Australia has been amply documented in over 100 references and reports summarized by Tyler (2012) and Tyler et al. (2012). Similarly, a large amount of research was carried out in the USA (Arthington et al. 2005; Vendramini et al. 2006). Much of this research has addressed the consequences of early weaning on the dams' reproductive performance (Fordyce et al. 1988; Schlink et al. 1992; Short et al. 1996; Tyler et al. 2012). In the majority of cases, weaners have either been raised in feedlots (Arthington et al. 2005) or supplemented with concentrates or crop by-products on pasture (Vendramini et al. 2007; Vendramini and Arthington 2008). Consequently, Tyler (2012) indicates that the effect of tropical pastures on the performance of early weaners should be prioritized in future pastoral research.
Early weaning has rarely been investigated in the neotropics (Moore and da Rocha 1983; Betancourt-López et al. 2012) and, as elsewhere, the practice has generally been coupled with regular supplementation of cows and calves with different combinations of concentrates and/or other feedstuffs in different settings. However, to the authors' knowledge, raising early weaned beef calves exclusively on sown pastures in the neotropics has not been investigated.
The objectives of this study were to assess the effects on cow and calf performance of early weaning of beef calves onto sown tropical pastures, while their dams were maintained on savanna, in comparison with cow-calf pairs grazing native savanna with weaning at the conventional age.
Materials and Methods
Experimental design
The study was conducted during the late 1980s over 6 consecutive years at Carimagua Research Station, located 87 km northeast of Puerto Gaitán in the Meta Department on the eastern plains of Colombia (4°36'44.6" N, 74°08'42.2" W). Monthly rainfall, ambient temperature and their annual variations were recorded during 13 consecutive years (Table 1).
Table 1 Monthly average climatic data over 13 years, and annual rainfall recorded during the 1984-1987 period at Carimagua Research Station, Meta Department, Colombia.
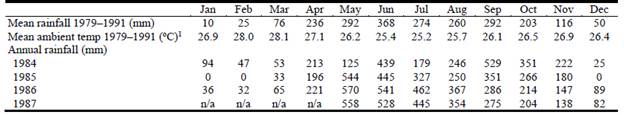
n/a - records not available.
1Mean monthly temperature for 1984-1987 not included as there is less than 1 °C variation between years.
Care of animals and experimental procedures were performed by accredited Doctors of Veterinary Medicine (DVM), including the second author, following national husbandry and animal welfare regulations.
During the first 2 years (Years 1 and 2) exploratory trials were conducted to assess the feasibility of raising early weaned beef calves on sown pastures alone, and to monitor the associated health and mortality risks. At the end of the experimental phase of Year 2, the weaners were transferred to a savanna paddock and their weight was monitored until the savanna controls were weaned at the age of 266 ± 7 days. A further 18 months were required to monitor growth of these weaners on savanna during 537 days after the early weaning date, and of the performance of their dams until the subsequent calving event. In the following 2 years (Years 3 and 4), trials compared the performance of early weaned calves on a number of sown tropical pastures with that of normally weaned calves on savanna.
In all cases Brahman and crossbred [Brahman x San Martinero (native; B. taurus)] cows and calves were used. Calves were born on savanna with a mean live weight (LW) of 22.7 ± 3.4 kg. Body weight of calves during the experimental phase on sown pastures was recorded every 7 days. Cows remained on large savanna paddocks as part of a large herd and were weighed and rectally palpated at ~4-monthly intervals to determine if they were pregnant. Calving events were recorded daily, but mustering of the cows was avoided near calving time. Pregnancy rates and inter-calving intervals were calculated. Approximate date of conception was back calculated from the calving date. Internal and external parasite infestations were controlled throughout following commercial farming practices. Mortalities and incidents of ill health were recorded.
Cows and calves had free access to fresh water plus a commercial mineral supplement containing (as-fed): 17.5% Na, 26.9% Cl, 8.0% P, 13.7% Ca, 2.0% S, 0.104% Cu, 0.35% Zn, 0.001% Co and 0.008% I. Supplement consumption was recorded every 15 days.
Animals, forages and grazing management
In the first year (Year 1) 10 male calves [112 ± 16 kg initial LW (ILW); mean ± standard error of the mean (s.e.m.)] were weaned at 166 ± 10 days old on 11 November 1984, coinciding with the end of the rainy season. They were rotationally grazed (6 calves/ha; 7 days grazing, 21 days rest) for 146 days during the dry season using 4 paddocks of equal size of a 6-year-old mixture of Andropogon gayanus cv. Carimagua-1 and commercial Pueraria phaseoloides (AgPp). Fifteen cow-calf pairs grazing savanna as part of a larger herd served as controls (Control 1) and were monitored at intervals of ~120 days. Their calves were weaned at 280 ± 29 days of age, and the dams were monitored until the next calving event. Savanna cows were stocked at 0.2 cows/ha in large (>200 ha) paddocks managed with periodic fire following traditional and regional farming practices (Kleinheisterkamp and Habich 1985; Rippstein et al. 2001).
In Year 2, the experiment was methodologically similar to Year 1, but 15 calves were weaned at 68 ± 9 kg ILW and 110 ± 8 days of age on 30 May 1985 at the beginning of the rainy season. They were rotationally grazed on 15 paddocks of AgPp and Ag plus Centrosema acutifolium cv. Vichada (accession CIAT 5277; AgCa; 6 calves/ha with 7 days grazing, 21 days rest) for 147 days. In parallel, 15 similar calves (Control 2) continued to suckle their dams as part of a large herd grazing native savanna, and were weighed at intervals of 21 days during the same period but weaned at 266 ± 7 days of age and 144 ± 20 kg LW. Thereafter, weaned control and early weaned calves were grazed on a native savanna paddock as a single group at 0.25 head/ha for 18 months, and were weighed about every 120 days.
During Year 3, the experiment compared the performance of groups of 10 weaned calves (68 ± 13 kg ILW, 93 ± 4 days of age) rotationally grazing 4 paddocks (6 calves/ha; 7 days grazing, 21 days rest) each of Ag, AgPp and AgCa for 123 days during the wet season, commencing on 25 June 1986 (mid rainy season).
Year 4 replicated the design of the third year, with the addition of a Brachiaria humidicola cv. Llanero (syn. B. dictyoneura)-Arachis pintoi (BhAp) pasture subjected to the same management and sampling practices previously described. The experiment began on 23 July 1987. Ten calves (81 ± 9 kg ILW, 86 ± 5 days of age) were weaned and placed on each improved pasture, while 10 unweaned calves grazed on savanna with their dams until weaning at 319 ± 29 days of age and 155 ± 25 kg live weight.
In Year 3 a pilot test of weaning was carried out starting on 28 July 1986 with one commercial herd at Carimagua. Forty-six calves born in 2 different savanna paddocks were weaned at a mean age of 186 ± 62 (range 54-285) days and LW 131 ± 26 kg, and were transferred to a 1-year-old A. gayanus-Stylosanthes capitata cv. Capica pasture stocked at 3 calves/ha for 99 days until the end of the rainy season.
Sample collection and lab analyses
Pasture. Pre-grazing herbage mass and botanical composition of the introduced pastures were estimated by the BOTANAL method (Tothill 1978; data partially shown). Botanical composition, growth rate and nutritive value of the savanna have been previously described by Rivera Sánchez (1988).
Blood. Jugular blood samples were collected weekly from each of the calves into two 10 ml BD Vacutainers® (Becton Dickinson, Franklin Lakes, NJ, USA) for hematology [hematocrit (g/100 ml)], serum enzymes [aspartate aminotransferase (AST, U/ml) and gamma-glutamyl transferase (GGT, U/ml)], total protein (g/100 ml), renal function [urea nitrogen (BUN, mg/100 ml)] and mineral [P, Ca and Mg (g/100 ml)] analyses. Hematology and serum biochemistry analyses were performed at the International Center for Tropical Agriculture, Cali, Colombia. Blood enzymes, protein and BUN were determined using standard kits (Sigma-Aldrich Corp., St. Louis, MO, USA), N by the micro-Kjeldahl method, P by colorimetry and the remaining minerals by atomic absorption.
Feces. Fecal grab samples were individually collected from the rectums of animals at weighing times to determine P, Ca, ash and N concentrations, with N expressed as percent of fecal organic matter.
Statistical analyses
Data were analyzed using the Statistical Analysis System, version 9.4 (SAS Institute, Cary, NC, USA). Results for the first 2 years were summarized using descriptive statistics (means ± s.e.). In Years 3 and 4, calves were balanced for LW and randomly allocated to sown pastures considering in all cases individual animals as the experimental unit. Data distribution from all variables examined (i.e. blood, feces and LW) was reviewed prior to additional analyses. Repeated-measures of blood, feces and LW for the same calf were analyzed with the GLIMMIX procedure, using a linear mixed model that included the fixed effect of pasture (i.e. sown forages and savanna), and the interaction between pasture and the random effects of year. All interactions were initially included, and those that were not significant were discarded for the final analysis (Gbur et al. 2012).
Final analyses were preceded by a study of the covariance structure (Gbur et al. 2012) to adjust the model specification as required. Differences were considered significant when P<0.05, and there was tendency to significance if P<0.10. Denominator degrees of freedom for the test of fixed effects were specified by the Kenward-Roger procedure. Multiple comparisons of least squares means used the Tukey procedure, complemented with graphical interpretation using SAS diffograms (not shown due to space limitations). Regressions were calculated with the GLMSELECT procedure.
Results
Forages and botanical composition
Pre-grazing herbage mass in all introduced pastures and across the experimental periods in Years 3 and 4 always exceeded 2,500 kg DM/ha, except in the Ag treatment in Year 3 for a brief period during temporary flooding of the paddock. Legume percentage in the forage on offer was higher for the AgPp (29%) mixture than for the BhAp (20%) association, with lower amounts (7-10%) in the AgCa sward. These legume percentages remained rela-tively stable throughout the experimental period as shown for Year 3 in Figure 1. The Ag pasture averaged 758 g/kg DM of neutral detergent fiber (NDF) and 80 g/kg DM crude protein (CP) during the rainy season, and 759 and 89 g/kg DM in the dry season, respectively. The AgPp and AgCa pastures had similar nutritive composition and averaged 756 g NDF and 115 g CP/kg DM and 757 g NDF and 110 g CP/kg DM for the rainy and dry seasons, respectively. Similarly, the BhAp pasture contained 716 g NDF and 84 g CP/kg DM in the wet season, and 691 g NDF and 21 g CP/kg DM in the dry season.

Figure 1 Forage on offer (kg DM/ha) in 3 Andropogon-based pastures and their respective botanical compositions in Year 3. Grass DM stands for total grass DM on offer in the respective pastures; Green DM is green grass DM; Leg DM is total legume DM. Ag = Andropogon gayanus; Pp = Pueraria phaseoloides; Ca = Centrosema acutifolium.
Mortalities
Over the 4 years of the study, 4 deaths out of 95 early weaned calves occurred in the AgPp paddocks in Year 3, which was related to a temporary flooding event. No calf mortality of control, suckling calves or their dams was recorded in the savanna paddocks.
Liveweight performance
Years 1 and 2. Data in Table 2 show the LWs and ages of calves during Years 1 and 2. Daily LW gains (LWG) of calves weaned at 166 days of age in Year 1 and grazed on the AgPp pasture averaged 0.10 ± 0.03 kg/head, whereas contemporary suckling calves on savanna gained 0.35 ± 0.19 kg/head. In Year 2, calves weaned at 110 days of age and grazed on AgPp and AgCa pastures gained 0.19 ± 0.06 kg/day, and control suckling calves gained 0.60 ± 0.13 kg/day.
Table 2 Mean (± s.e.) live weights and ages of early weaned Brahman and Brahman cross calves during the exploratory observations in Years 1 and 2.

n - number of animals.
Live weights of cows for Years 1 and 2 between consecutive calving seasons on savanna are shown in Figure 2. Year 1 dams of early weaned calves calved in the late rainy season, while control cows (normally weaned) calved at the end of the subsequent dry season (1985-1986). Early weaned dams in Year 2 calved at the end of the dry season, whereas control cows did so at the end of the following rainy season. Calculated LWs at conception for the 2 years were 303 ± 43 and 321 ± 45 kg, respectively.

Figure 2 Postpartum cows' live weights in relation to type of weaning. Early and normal refer to weaning treatment, followed by the calves' ages at weaning. Standard deviations are not shown for clarity, and ranged between 32 and 42 kg, and 24 and 55 kg for early and normal weaning, respectively. Vertical arrows indicate weaning times and ellipses indicate time and spread of calvings.
Years 3 and 4. Consistent with the preliminary observations, calves were weaned at average ages of 93 and 86 days in Years 3 and 4, respectively (Table 3). At the end of the 123 days of experimentation, LW was lower (P<0.0001) in calves on improved forages than in their suckling counterparts on savanna (Table 3). Differences in LW productivity amongst sown forages were small (P>0.05), with the exception of Ag in Year 3 (Table 3).
Table 3 Least squares means (± s.e.) of ages and live weights (LW) of early weaned calves at weaning and their final LW (FLW) after 123 days of experimental grazing on improved pastures (early weaned) and of normally weaned calves off savanna.

n - number of animals.
n.a. - not applicable.
NS - not significant.
1Within columns values followed by different letters differ significantly (P<0.05).
2Weights and ages of savanna calves were obtained on the same date (± 3 days) as those of the early weaners.
Data on daily LWG (DLWG, g/head) of calves for each individual grazing period and for each of the 3 Ag-based pastures in 1986 were pooled and regressed on the amount of green grass leaf on offer (GGL; kg DM/ha). The resulting linear regression equation was: DLWG = 0.702 ± 0.156 GGL - 166.667 ± 98.700; r2 = 0.63, P<0.01.
Pilot test
Despite the very low LWs of some of the early weaned calves, no deaths occurred. Weight gains during the sown pasture phase averaged 0.18 ± 0.10 kg/day, while the correlation between weaning weight and subsequent weight gain on the sown pasture was not significant (r2 = 0.24, P>0.05).
Blood and feces profiles
Blood data showed that the concentrations of total protein, urea nitrogen and the AST and GGT enzymes in weaned calves differed significantly (P<0.0001) between years, but there were much smaller differences in concentrations of protein and enzymes amongst the pastures (Table 4). In Year 3, blood urea nitrogen levels on the AgPp pasture were 8-10 times those on the other pastures (P<0.0001), associated with a temporary increase in the percent legume. Over the same period, relative to calves grazing Ag as monoculture, hematocrit concentration was higher (P<0.001) in AgCa and AgPp mixtures by 31 and 19%, respectively. However, although within normal physiological values, a larger hematocrit difference (58%; P<0.001) was found with the unweaned calves on savanna (Table 4).
Table 4 Least squares means for total blood protein (TBP), hematocrit (HCT), urea nitrogen (BUN) and the body fluid enzymes aspartate aminotransferase (AST) and gamma-glutamyl transferase (GGT) of early weaned calves grazing sown pastures and un-weaned calves grazing savanna in Years 3 and 4.

n/a - not available.
Concentrations of minerals in serum and feces (Table 5) varied considerably between years (P<0.0001). However, there were small and mostly non-significant differences between pastures. There were no effects of nutritional treatment upon ash or fecal N, on either a DM or an organic matter basis (Table 5).
Table 5 Least squares means for blood serum phosphorus and calcium (mg/dl), fecal phosphorus, calcium and ash (% fecal DM, FDM) and fecal nitrogen (% fecal organic matter, FOM) concentrations (means ± s.e.) of early weaned calves grazing sown pastures and un-weaned calves grazing savanna in Years 3 and 4.

n/a - not analyzed.
NS - not significant.
1Letters compare pasture values within years. Differences were found between pastures in different years, but not within the same year.
Cow live weights and reproductive performance
Early weaned cows were significantly heavier at weaning than those weaned at normal times (Table 6; P<0.05). Inter-calving intervals increased significantly (P<0.05) with increasing calf weaning age, but were inversely related to cow weight at weaning (P<0.001), with the positive effect of cow weaning weight in reducing inter-calving interval being greater with older than with younger weaning ages (regression equation in Table 6).
Table 6 Reproductive performance and live weight (LW) of cows (n = 45) in relation to age and weight of calves at weaning.
| Weaning age (d) | Inter-calving interval (d)1,2 | Cow weaning LW (kg) | Calf weaning LW (kg) |
| 110 | 472d3 (454-491) | 339a ± 8 | 68a ± 4 |
| 166 | 514c (504-557) | 308b ± 11 | 112b ± 5 |
| 198 | 625a (594-656) | 304b ± 11 | 114b ± 5 |
| 266 | 642a (616-669) | 306b ± 9 | 143c ± 4 |
1Regression equation:
Inter-calving interval (d) = 394 (± 38) + 2.76 (± 0.59) * calf weaning age - 0.00588 (± 0.0019) * calf weaning age * cow weaning LW; adj R2 = 0.45; P<0.001.
2Confidence interval in parentheses.
3Within columns values followed by different letters differ significantly (P<0.05).
Calf compensatory body growth
The possible carry-over effects of low calf weaning LW were examined by monitoring subsequent performance of early weaned calves compared with normally weaned calves, for a total of 414 days after the end of the early weaning experimental period (Table 7). At that time, there were no significant (P>0.05) differences in final LW between the early weaned and normally weaned calves (Table 7). Over the 414 days of common grazing in savanna, daily weight gains were inversely related to weaning weight, weight gains to weaning, and weight at end of the experimental phase (r2 = 0.98, P<0.05 in all cases).
Table 7 Final live weights (LW) of contemporary calves (n) weaned early and grazed on various sown pastures for 123 days and normally weaned calves, following 414 days grazing savanna as a single group. Data are least squares means ± s.e.m.
| Weaning pasture | n | LW (kg) |
| A. gayanus | 10 | 187 ± 19 |
| A. gayanus plus C. acutifolium | 10 | 187 ± 16 |
| A. gayanus plus P. phaseoloides | 10 | 192 ± 13 |
| Traditional weaning on savanna | 10 | 205 ± 16 |
| Probability | NS |
n - number of animals.
NS - not significant.
Discussion
The primary objective of this study was to assess the growth of early weaned calves on sown tropical pastures, while keeping the breeding cows on savanna. A secondary objective was to document the effect of early weaning (3-4 months) on inter-calving interval in breeders compared with traditional rangeland cow-calf breeding, where calves are weaned at about 9-10 months old. The low overall mortality rate of calves of 1% over 4 consecutive years was a significant finding when compared with commercial, extensive tropical herds that normally exhibit death rates of 7-9% (Kleinheisterkamp and Habich 1985; Rivera Sánchez 1988). Reduced calf mortality would then complement the improved reproductive performance of the breeding cows in terms of reduced inter-calving intervals associated with early weaning.
There has been limited research on management of early weaners on supplemented tropical pastures (Vendramini et al. 2008; Vendramini and Arthington 2008), and even less if unsupplemented. Post-weaning LW gains by early weaned calves were low and similar across the 4 years of experimentation (Tables 2 and 3) allowing for the differences between years in age of calves at weaning. Schottler and Williams (1975) compared the performance of Brahman-Shorthorn crossbred calves weaned at 4, 5, 6 or 7 months of age on a Para grass (Brachiaria mutica)-Siratro (Macroptilium atropurpureum) sward for 2 months, followed by a buffel grass (Cenchrus ciliaris)-Siratro pasture. Regardless of weaning age, LWG ranged between 0.20 and 0.32 kg/day. Holroyd et al. (1990) weaned calves in northern Australia at 5 and 8 months of age, and after a 10-day period of supplementation with good quality pasture hay they were transferred to a savanna paddock. The LW difference at 8 months between early and late weaners of 54 kg for males was reduced to 13 kg at the age of 3.5 years due to compensatory growth in younger weaned calves, and the research indicated that pasture quality was probably the limiting factor for better animal performance.
This hypothesis is supported by the observation that the LW of early weaners (100 days) placed on a higher quality annual ryegrass (Lolium rigidum) temperate pasture did not differ from those of late weaners at 365 days of age (Potter et al. 2004). The close correlation between green leaf and LWG found in the present research supports the hypothesis that performance of early weaners on pastures may be limited by forage quality. In this context, Aguiar et al. (2015) advocated limited creep feeding with soybean meal to improve the performance of early weaners on limpograss (Hemarthria altíssima).
Casual visual observations showed highly selective grazing on the Ag-based pastures. In earlier studies (Bõhnert et al. 1985; 1986), young steers grazing AgPp mixtures selected diets much higher in N than when grazing Ag as a monoculture but differences in in vitro DM digestibility of the diets selected were not significant. Although some caution is required in extrapolating selective grazing behaviors between weaners and 1 -2 year old steers, the high BUN concentration on the AgPp pasture in Year 3 was likely due to temporary low availability of green leaf on Ag and high levels of Pp on offer, as Ag leaves were selectively grazed at the start of grazing in each paddock. However, as the availability of Ag leaves rapidly declined, calves were forced to consume the legume. The preliminary, positive relationship between green grass on offer on all Ag-based pastures and daily LWG points in the same general direction.
Overall, this study also showed that none of the early weaned calves demonstrated a deficiency of total protein, hematocrit, BUN and the sensitive enzyme marker of liver damage, AST (Table 4). However, compared with blood reference values, results from all pastures in Years 3 and 4 indicated low GGT activity, and high variability between animals (Table 4). Although the potential physiological effects of these values need to be clarified, it is reasonable to assume, as demonstrated by Stojevic et al. (2005) with healthy dairy cattle, that the observed GGT concentrations reflected a temporary acute situation (i.e. circadian changes) associated with age of the calves, rather than long-term detrimental metabolic effects. Furthermore, it is unfortunately impossible to define whether the low enzyme concentrations suggested adverse metabolic effects of secondary compounds in the legumes. Nevertheless, it is worth noting that recent studies (Ramírez-Restrepo et al. 2016) demonstrated that supplementation of animals with plant-derived compounds increases GGT blood serum values in Brahman cattle, which is contrary to the present results.
Despite differences between pastures and years in mineral concentrations in serum and feces (Table 4), all values fell within normal ranges (Doornenbal et al. 1988; Aiello and Moses 2016). The fairly large between-year differences, and the between-animal variation indicated by the relative magnitude of the standard error terms, question the reliability of single samplings within years and pastures, and by inference, between farming systems as frequently carried out in survey studies. Several mineral deficiencies in unsupplemented adult beef cattle grazing savanna, accompanied by low breeding cow LWs, have been reported (Lebdosoekojo et al. 1980). Subsequent and detailed analyses by Rivera Sánchez (1988) in controlled savanna experiments over several years showed adequate serum, liver and fecal concentrations of all minerals in cows, when complete mineral mixes were provided. Furthermore, Rivera Sánchez (1988) found evidence of an interaction of mineral supplementation with management strategies that allowed access to improved grass-dominated pastures by the breeding herd.
Although fecal N as an indicator of nutrition has sometimes been questioned (Hobbs 1987), it is generally regarded as appropriate for free-ranging herbivores in the absence of better, simple indices (Leslie Jr. et al. 2008). Similarly, N in feces has been found to be broadly and linearly related to N intake with a variety of forage diets (Nunez-Hernandez et al. 1992). Allden and Jennings (1969) proposed that fecal N levels of 1.4-1.6% in sheep are indicative of N-limited diets. If these values are applicable to calves, data in Table 5 would suggest that dietary N would not have been the limiting nutritional variable. This view accords with that expressed by Lascano (1991), who showed that digestible energy intake is the most limiting nutrient for yearlings and adult cattle in neotropical savannas.
The low calf LWs at the end of the experimental periods, coupled with absence of indicators of specific nutritional deficiencies shown by the blood and fecal analyses, are indicative of a general condition of under-nutrition, a hypothesis supported by the in-depth analyses of metabolic profiles of early weaned calves in tropical northeast Argentina. There, Coppo (2003; 2007a; 2007b) assessed the stress produced by early weaning at 60 days of age in crossbred Zebu cattle supplemented with concentrates, but could not relate it to a large set of blood parameters and suggested that there was no evidence of specific metabolic stresses. Arthington et al. (2005) studied the dynamics of acute-phase proteins in beef calves weaned at 89 days of age onto pasture and fed a supplement, and found them to rise in the first few days following separation from the dams, decreasing subsequently to normal values. While early weaned calves were lighter than contemporary suckling calves at 120 days of age, and had lower concentrations of BUN, total proteins, triglycerides, P, Mg, Fe and Cu, indicators of stress such as cortisol, aldosterone and AST did not differ between the 2 groups of calves. The authors concluded that early weaning does not produce clinical stress in crossbred Zebu calves, despite a general condition of under-nutrition.
One measure of the sustainability of beef herds is absence of animal stress, and the present results, supported by the literature, would suggest that stress in early weaned beef calves would have been short-term only. Furthermore, and despite the condition of generalized under-nutrition, there was no evidence of negative carry-over effects. In fact, by the end of the observation period, there were no significant differences in the final LWs (Table 7) despite the early advantage of animals allowed to suckle for the normal time, a finding confirmed by the longer-term on-farm study of Mejía et al. (2009), who weaned female and male calves at 4 months of age on a rotation of B. decumbens, B. humidicola and savanna pastures plus a medium quality concentrate until reaching 8 months of age. Thereafter these and normally weaned calves (8 months old) were raised on the same pastures as above without supplementation. No significant differences were found in age at first conception (35 vs. 33.4 months for early and traditional weaning, respectively). The inter-calving interval of early weaned dams was 141 days shorter than those weaned late. The corresponding males reached slaughter weight (450 kg) with a non-significant difference of 3.2 months and average monthly LWGs of 11.5 vs. 10.6 kg for late and early weaners, respectively. Under more severe pasture conditions, Holroyd et al. (1990) noted that the early weaners (5 months old at weaning) grazing savanna in northern Australia were still 13 kg lighter than late weaners (8 months old at weaning) at 3.5 years of age.
In view of the compensatory growth experienced by the early weaners in the present study, the low LWG recorded on pasture would be acceptable if, as is generally the case, weaners are not sold immediately, but are kept as young steers for an additional 12-18 months. This trade-off between calf LWG and cow reproductive performance is an important consideration for farm managers who need to balance different forage resources and the nutritional needs of stock and prioritize their use. The new 'crop' of improved, high-yielding and leafier Brachiaria cultivars (Pizarro et al. 2013), together with higher quality grasses such as Panicum maximum, may help resolve the above issues to some extent, if the Brachiaria cultivars do not lead to subclinical and clinical photosensitization (Lima et al. 2012).
Notwithstanding few to no weight differences in later life, possible negative effects of low early growth rate on lifetime beef production of early weaned females through epigenetic effects affecting them and their progeny cannot be ignored (Martin et al. 2007; Funston et al. 2012;Wathes et al. 2014), although this would probably be a minor concern in extensive systems.
In our study, early weaned male calves attained weights at 18 months of age similar to those of control animals of equivalent ages. Vera (1991) and Vera et al. (1993) showed that, despite long periods of sustained under-nutrition, heifers and cows could achieve mature body sizes and inter-calving intervals similar to those of better-fed animals, when given the opportunity to make moderate compensatory gains. In the present study, weaning calves early removed lactation stress on cows, and allowed them to attain plateau weights of 350-400 kg, confounded with pregnancy, even during the dry season. While LWs of this magnitude are seldom observed in traditional savanna-based grazing systems (Rivera Sánchez 1988), weights of Zebu cows above 320 kg do not appear to limit re-conception (Mukasa-Mugerwa 1989; Vera et al. 1993). In fact, cows weaned at 166 days in the present research conceived after weaning with an average weight of 303 ± 43 kg.
Early weaning is known to enhance reproductive performance of underfed beef cows (Moore and da Rocha 1983; Holroyd et al. 1990; Schlink et al. 1994; Coppo et al. 2002; Arthington et al. 2004), an effect shown also in Table 6 that demonstrates the trade-offs between weaning age and LWs of calves, LWs of their dams and inter-calving intervals. Moore and da Rocha (1983) investigated the effects of 2 levels of nutrition and 5 weaning ages in Zebu Gyr breed cows fed hay of B. decumbens in the Brazilian Cerrados and found that, irrespective of the cows' nutritional level, early weaning of calves improved cow weights at different stages throughout the reproductive cycle and subsequent reproductive performance. Weights of cows at weaning were 313 kg and 325 kg on low and high supplementary energy treatments, respectively, and the authors suggested that Zebu cows rarely conceive if suckling cows weigh less than 300 kg. Weights of cows at weaning decreased linearly from 352 kg, when calves were weaned at 1 month of age, to 294 kg if calves were weaned at 6 months of age, with LW losses during lactation increasing from 21 to 102 kg for the respective weaning ages (Moore and da Rocha 1983).
Mukasa-Mugerwa (1989) reviewed the literature regarding reproduction of Zebu cattle in the tropics, and noted that tropical cattle dependent on natural pastures most often calve in alternate years, but animals with access to good quality sown pastures have improved reproductive performance. For example, Rivera Sánchez (1988) reported inter-calving intervals of 618 days on well managed savannas over 4 years, whereas Vera et al. (2002) found an average interval of 445 days on well managed B. decumbens. Equally large differences have been reported by other authors (Arthington et al. 2004). Hale (cited by Mukasa-Mugerwa 1989) found that, when the LW of suckling Zebu cows fell from 390 to 320 kg, they stopped cycling, but they needed to reach weights in excess of 320 kg to start cycling again. As shown in Table 6, only cows, whose calves were weaned at 110 days, showed LWs above the purported lower critical weight of 320 kg. Inter-calving intervals increased significantly with increasing weaning age and were negatively related to cows' weights at weaning (P<0.001; footnote of Table 6), whereby the positive effect of cow weaning weight on inter-calving interval was greater with larger than with lower weaning ages (Table 5).
Lastly, early weaning of beef calves and the consequent changes in reproductive performance will likely lead to significant changes in herd dynamics (Turner et al. 2013), whose production, environmental and economic effects remain to be studied. These changes would significantly impact management decisions (Sullivan et al. 1997). The savannas of the Orinoco watershed are under transition as represented by: (i) increase in crop and tree plantation areas; (ii) oil exploration and extraction; (iii) mining; (iv) growing recognition of indigenous rights and lands; (v) and increasing appreciation of the savanna's relevance in terms of biodiversity and contribution to greenhouse gas emissions (Rausch 2013; CONPES 2014). This transition has been also noted in the Australian savannas (Holmes 2010). This implies that extensive beef cattle farming, even if it continues to represent an important land use system, will need to adapt and intensify to the extent possible. Further, farmers and their communities will need to modify their decision-making to take into account the multifunctional traits of these lands. The strategic use of areas of sown pasture could play an increasingly important role in management of breeding herds.
Conclusions
Results from this study suggest that there is considerable scope and flexibility in strategic use of small areas of sown pastures for weaners in combination with extensive savannas to improve productivity of beef breeding herds, if calves are given the opportunity to realize compensatory growth. Several authors have commented on the flexibility, adaptability and sustainability of extensive systems (Davies et al. 2010; Astigarraga and Ingrand 2011), while some of the environmental aspects have been also described (Ramírez-Restrepo and Charmley 2015) or are under scrutiny (Ramírez-Restrepo et al. unpublished data 2017). This study has demonstrated over 4 consecutive years, that early weaning of calves onto a variety of sown tropical pastures is technically feasible, resulting in improved LWGs and reproductive rates in their dams.
Adoption of such early weaning strategies in extensive systems, where mating generally extends over 6 or more months, would be constrained by the spread of the calvings over an extended period of time. A number of weaning events would be needed each year, resulting in increased labor and management requirements (Sullivan et al. 1997). On the other hand, with several batches of early weaned calves dispersed over several months, a smaller area of sown pastures would be needed than if all calvings were concentrated into a shorter period of time. From a management point of view, seasonal mating to facilitate early weaning would be desirable, but this would lead to a smaller calf crop in the initial year when the practice was first implemented. However, increased reproductive rates during the following years due to improved cow condition and LW, as demonstrated in this study, would soon compensate for the loss of production in the implementation year of seasonal mating. A possible added benefit would be a reduction in risks, which could be posed by climate change. Thus, productive and environmental trade-offs, as suggested above, would need better quality and intensity of animal and pasture management. Further savanna studies with larger numbers of calves and cows under commercial conditions would confirm these preliminary findings.

