











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink1. Introduction
The Brazilian melon production in year 2015 was 521 596 metric tons and the planted area was 20 837 hectares [1]. The Northeastern region was responsible for 87% of that production, especially the state of Rio Grande do Norte, which is the main producer and exporter of this fruit. This region has large volumes of groundwater in the aquifers Calcário Jandaíra (medium and high salinity water) and Arenito-Açu (low salinity water), mainly in the Apodi Plateau region. The abundance of water favors the practice of irrigation, but in some irrigated areas problems of soil salinization and loss of crop productivity have been observed due to inadequate soil, water and crop management [2].
The use of irrigation water with high electrical conductivity causes a reduction in fresh and dry matter yield of plants and a higher accumulation of calcium, magnesium and potassium [3]. In addition, salinity may cause limitations in plant uptake of nutrients such as nitrogen and phosphorus early in the vegetative cycle. This can cause restrictions on the development of the plants’ shoots and roots, without later recovery, even if there is a new supply of these nutrients. Considering that the harmful effects of salinity depend on crop tolerance and irrigation management, a way of reducing the effect of high water salinity on plants is to irrigate with saline water mixed with low salinity water at the most tolerant stages of the crop [4].
Among the environmental factors that affect crops, the soil class deserves to be highlighted, since the soil’s physical and chemical characteristics imply more or less difficulty in absorbing water and nutrients by plants. In this respect, [3] evaluated the growth and nutrition of melon cultivated in soil classes with different textures and verified that the dry matter production and the accumulation of Ca, Mg, Na, K and Cl were different among the soil classes. The clay content of the soil was identified as the main factor determining the effects of soil salinity.
Given the need to use water with different salt levels to irrigate melon crops, this study evaluated how Galia melon plants responded in the nutritional aspect to the salinity of irrigation water in different soils of the semiarid region of Rio Grande do Norte state, Brazil.
2. Material and methods
2.1 Local, climate, and structure
The experiment was conducted in a protected environment of the Federal Rural University of the Semiarid, located in the municipality of Mossoró, RN, with geographic coordinates 5º11'31 S and 37º20'40" W, and average altitude of 18 m. The dominant climate of the region, according to the classification of Köppen, is BSwh', namely hot and dry, characterized as semiarid tropical climate, with irregular rainy season concentrating on the first months of the year, and lasting from summer to autumn.
The protected environment had a metal arched structure covered by a transparent plastic sheet treated against the action of ultraviolet rays. The place was closed laterally with a nylon net providing 50% shade. The ceiling height was 3.5 meters and the internal dimensions were 7 meters wide and 18 meters long.
2.2 Experimental design and soil classification
The experimental design was completely randomized in a factorial scheme (5 x 6), with three replications. The factors studied were five soil classes: Neossolo Quartzarênico - RQ (Typic Ustipsamments), Argissolo Vermelho-Amarelo - PVA (Typic Haplustults); Cambissolo Háplico - CX (Typic Haplustepts), Neossolo Flúvico - RY (Typic Ustifluvents) and Vertissolo Háplico - VX (Typic Haplusterts), and six water irrigation salinity levels (expressed as electrical conductivity): A1 = 0.5, A2 = 1.0, A3 = 2.0, A4 = 3.0, A5 = 4.0, and A6 = 5.0 dS m-1. Soil classification was carried out according to Brazilian Soil Classification System [5] and correlated with Soil Taxonomy [6]. The physical and chemical characteristics of the soils are presented in Table 1.
Table 1 Chemical and physical characteristics of soils cultivated with melon in the western region of the state of Rio Grande do Norte, Brazil

RQ - Typic Ustipsamments; PVA - Typic Haplustults; CX - Typic Haplustepts; RY - Typic Ustifluvents, and VX - Typic Haplusterts; SOM - soil organic matter; SB - sum of bases; T - cation exchange capacity at pH 7.0; CEC - cation exchange capacity; V - base saturation; ESP - exchangeable sodium percentage; SD - soil density; PD- particle density; Msat - saturation moisture; Ptot - total porosity.
Source: The Authors.
The soils at each sampling site were collected in trenches at depths of 0-20 and 20-40 and their physical and chemical characteristics are presented in Table 1.
2.3 Water characterization
The water used in the study was obtained from two aquifers: Arenito Açu, with depth of approximately 1,000 m and water with low salinity (electrical conductivity around 0.5 dS m-1); and Calcário Jandaíra, with depth of around 80 m and water with high salinity (electrical conductivity around 5.0 dS m-1), with the predominant cations of Na+ and Ca++ and anions of HCO3 - and Cl-. The chemical characterization of these waters is presented in Table 2. The water salinity levels of 1.0 dS m-1, 2.0 dS m-1, 3.0 dS m-1, and 4.0 dS m-1 were obtained from mixing water samples from the two aquifers in different proportions. For this, the electrical conductivity was measured with a commercial benchtop conductivity meter, which was calibrated using a KCl solution (1.0 mmol L-1) in a controlled temperature environment at 27.0 ± 1.0 ºC. During the experiment, the electrical conductivity was periodically measured with a pocket conductivity meter with automatic temperature compensation.
Table 2 Chemical characteristics of water used in different mixtures for irrigation of melon in different soils of the western region of the state of Rio Grande do Norte, Brazil

EC - electrical conductivity; SAR (sodium absorption ratio) = Na+ /[(Ca+2 + Mg+2)/2]0,5 ; Class - water classification of Richards (1954). AA -Arenito Açú Aquifer; CJ - Calcário Jandaíra Aquifer.
Source: The Authors
2.4 Experimental procedures
The crop was the Galia type melon (Cucumis melo L.) of the cultivar Néctar, which presents medium cycle and fruits of spherical shape, weighing between 0.8 and 1.2 kg. The skin has a dense netting of rough lines; the color of the pulp is green and its yield is around 22,000 kg ha-1. Pest and disease control was performed preventively with insecticides and fungicides specific for the crop, in order to avoid the presence of the pests and diseases most common in the region.
The experimental units consisted of polyvinyl chloride cylinders with controlled water drainage. They were 40 cm high and 20 cm in diameter and had capacity of 12 liters of soil. They were arranged in rows with spacing between them of 1.0 x 0.5 m. After being filled with soil, the cylinders were kept moist for incubation of the fertilizers, and then five melon seeds were sown in each one.
Soil moisture was monitored daily by tensiometers installed in each treatment for irrigation management purposes. Irrigation was carried out with test tubes containing the amount of water determined from the tensiometer readings to raise the soil moisture to field capacity. For this, soil water retention curves were plotted by the Tension Table and Richards Chamber methods (Fig. 1).

2.5 Measurements performed
The nutritional status of melon plants was evaluated by foliar diagnosis at 45 days after planting, when the plants were in the flowering phase. For this, the plants were cut at ground level and separated into leaves and stems, which were dried in an oven with forced air circulation at a temperature of 65 ºC until reaching constant weight. Next, the dry leaf material was ground in a Wiley mill for macronutrient and micronutrient measurement in the laboratory.
The N content of the leaves was determined after extraction by sulfur digestion. For this purpose, 0.2 g of the dry matter was digested in 3 mL of sulfuric acid, 1 mL of hydrogen peroxide and 1 g of a 10:1 mixture of potassium sulfate and copper sulfate. The samples remained in the digester block for half an hour until presenting a brown color, when the temperature was raised to 350 °C for two hours until the samples presented greenish color, when the extracts were completed to the volume of 25 mL. The method used to obtain extracts was steam distillation (Kjeldahl method), as described by [7].
The contents of the other nutrients were determined after extraction by nitric digestion in a microwave oven. For this, 0.5 g of dry matter was used, which was digested in 5 mL of nitric acid. In the digested extract, the phosphorus content was determined by molybdate blue spectrophotometry, the K, Na and Ca contents were determined by flame emission photometry, and the Mg, Cu, Mn, Fe and Zn were determined by atomic absorption spectrophotometry, according to [9].
2.6 Statistical analysis
The data were submitted to analysis of variance with the aid of the “Sistema para Análises Estatísticas” software [10]. The effect of salinity levels was evaluated by means of regression analysis, for which the coefficients of the models were tested based on the mean square residual of the ANOVA.
3. Results and discussion
A significant effect was observed of the interaction between the factors soil class and irrigation water salinity level for macronutrient and micronutrient contents. The nutrient content in the leaves, in addition to reflecting the plants’ nutritional status, shows the effect of irrigation water salinity on nutrient uptake by plants.
The increase in the irrigation water salinity from 0.5 to 5.0 dS m-1 promoted a linear increase in the contents of phosphorus, potassium, and sodium in the soils studied (Fig. 2). The exceptions were the contents of phosphorus and of potassium in the Typic Haplustepts, which presented quadratic increases.

Source: The Authors.
Figure 2 Contents of phosphorus (A), potassium (B) and sodium (C) in leaf blades of flowering melon irrigated with water of different salinities and in different soil classes: RQ - Typic Ustipsamments; PVA - Typic Haplustults; CX - Typic Haplustepts; RY - Typic Ustifluvents, and VX - Typic Haplusterts.
3.1 Macronutrients
Phosphorus, besides being in the composition of adenosine triphosphate, acts to promote the storage and transport of energy for endergonic processes, such as the synthesis of organic compounds and the active absorption of nutrients. Potassium is important for plants under stress conditions, with influence on osmotic properties, opening and closing of stomata, photosynthesis, enzymatic activation, protein synthesis and carbohydrate transport [11]. Sodium can stimulate the growth of C4 plants, aiding the movement of substrates between the mesophyll and the bundle sheath. It can also replace potassium in its osmotic role [12]. However, the increase in sodium concentration in leaf chloroplasts causes osmotic, ionic and oxidative stresses that result in low photosynthetic performance and reduced growth and yield [13].
The phosphorus contents in the melon leaf tissue (Fig. 2A) increased with the increase in irrigation water salinity in all soils studied and were below the values considered adequate by [14], which are between 3 and 7 g kg-1. The exception was the Typic Haplustepts, in which phosphorus levels were within the range mentioned, at all salinities. The waters with highest salinity provided contents of 2.21, 1.59 and 1.08 g kg-1 of phosphorus for the soils Typic Haplusterts, Typic Ustifluvents, and Typic Ustipsamments, respectively. At the lowest levels of water salinity, the lowest phosphorus content (0.53 g kg-1) was observed in the Typic Ustipsamments. High levels of P were observed in Haplustepts by [15], when studying the phosphate fertilization of melon plants. The authors explained that these contents are due to the soil parent material, namely limestone of the Jandaíra formation.
The accumulation of phosphorus in the plants grown in Typic Haplustepts was greater than in the other soils, and showed an increase of 60% for the water of highest salinity in relation to the lowest salinity. The higher amount of this element occurred due to the high natural fertility of this the soil (Table 1). Phosphorus content in the plant leaf blades in the present study was within the average found in other melon cultivars, which varied between 0.4 and 6 g kg-1 in the works of [16] and [17].
The foliar potassium contents increased by 77, 45, 43, 64 and 116% for the Typic Ustipsamments, Typic Haplustults, Typic Haplustepts, Typic Ustifluvents, and Typic Haplusterts, respectively, when irrigation water salinity increased from 0.5 to 5.0 dS m-1 (Fig. 2B). The potassium contents in the melon leaf tissue in most soils and irrigation water salinities were within the range considered adequate by [12], which is between 25 and 40 g kg-1, indicating that the plants were satisfactorily nourished at this stage of development. The exception was the Typic Haplusterts, in which potassium levels were below the range mentioned until the salinity of 3 dS m-1. Above this point, the contents of potassium were within the adequate range, which according to [17] is of 24 g kg-1.
When evaluating the leaf content of sodium during the initial growth of the crop, increases of 399, 253, 214, 66, and 58% for the soils Typic Haplustepts, Typic Ustipsamments, Typic Haplustults, Typic Ustifluvents, and Typic Haplusterts were observed, respectively, as a function of the increase in irrigation water salinity from 0.5 to 5.0 dS m-1 (Fig. 2C). The foliar contents of sodium varied between 0.8 and 4.14 g kg-1 in the studied soils. The highest content of sodium was observed in the plants grown in Typic Haplusterts (2.57 to 4.14 g kg-1). The greater accumulation of this element at the highest irrigation water salinity level, regardless of the soil class, agrees with the results obtained by [18] for the melon ‘Piel de Sapo Sancho’. The percentages of increase in foliar sodium contents indicated that the use of saline water poses a risk to the growth and yield of melon plants.
The increase in the irrigation water salinity from 0.5 to 5.0 dS m-1 promoted a linear increase in the melon leaf contents of magnesium, nitrogen, and calcium in nearly all the soils studied (Fig. 3). The exceptions were the contents of magnesium and of nitrogen in the plants grown in Typic Haplustepts, which presented quadratic increases. Magnesium is part of chlorophyll, and it is necessary for photosynthesis and synthesis of proteins and several enzymes that act on phosphate transfer [10] and [9]. Nitrogen has a strong effect on the development of plants, and when supplied to plants, causes an increase in leaf area and plant biomass, as the leaves remain green longer, thus increasing the plant’s photosynthetic capacity [19]. Calcium is important for cell wall and membrane stabilization, osmoregulation, and as a secondary messenger, allowing plants to regulate developmental processes in response to environmental stimuli [10]. According to [20], the reduction in transpiration as a consequence of saline stress can cause imbalance in calcium nutrition in plants.
The magnesium content in the leaves of the melon plants varied from 1.24 g kg-1 in the Typic Ustipsamments when the water of lowest salinity was used, to 10.61 g kg-1 in the Typic Ustifluvents irrigated with water with salinity of 5.0 dS m-1 (Fig. 3A). The increases in these contents were 65, 118, 160, 233, and 331% for the soils Typic Ustipsamments, Typic Haplustults, Typic Haplusterts, Typic Ustifluvents, and Typic Haplustepts, respectively. Even so, the contents of this nutrient in the melon plant leaves grown in the Typic Ustipsamments did not reach the range considered adequate by [12], which is between 5 and 12 g kg-1. In the other soils, the adequate contents were reached from the salinity of 4.0 dS m-1 in the Typic Haplusterts, 3.0 dS m-1 in the Typic Ustifluvents, 2.0 dS m-1 in the Typic Haplustults, and only at the salinity of 5.0 dS m-1 in the Typic Haplustepts. The levels of these ions observed in this study were higher than those reported by [21], who observed a content of 0.97 g kg-1 in the shoots of ‘Piel de Sapo Sancho’ melon plants.
The nitrogen content in the leaves of the plants varied between 18.26 g kg-1 in the Typic Ustipsamments irrigated with water of lowest salinity (0.5 dS m-1) and 35.82 g kg-1 in the Typic Haplusterts irrigated with water with salinity of 5.0 dS m-1 (Fig. 3B). In comparison, the contents of nitrogen in net melon leaves observed by [22] ranged from 20.65 to 21.90 g kg-1.
The increases in the foliar content of nitrogen were 18, 28, 37, 38 and 75% in the soils Typic Ustipsamments, Typic Haplustepts, Typic Haplusterts, Typic Ustifluvents and Typic Haplustults, respectively. In the Typic Haplustepts, Typic Haplustults, Typic Haplusterts and Typic Ustipsamments, for all salinities the nitrogen content of melon leaves was within the range considered adequate by [12], between 25 and 40 g kg-1. In the Typic Ustifluvents, this content was below that range for the salinities of 0.5 and 1.0 dS m-1. The adequate contents of nitrogen also agree with [23] and [24]. Considering that, generally foliar analysis in melon plants is performed at the beginning of flowering [24], which occurs around 37 days after sowing. So, it can be stated that independently of the treatment, the plants did not present deficiency of N. On the other hand, the results obtained in this study were higher than those found by [15], who obtained levels of 9.71 and 6.81 g kg-1 of N for the ‘Trusty’ and ‘Orange Flesh’ melon hybrids, respectively.
The content of Calcium of melon leaves was below the range between 25 and 50 g kg-1, considered adequate by [12], in most soils and salinities. The exceptions were the Typic Haplustults, which presented contents within that range for all salinities, and the Typic Ustifluvents, which presented contents within the range from salinity of 4.0 dS m-1. The increases in the foliar content of this nutrient were 36, 47, 122, 151, and 526% in the Typic Haplustults, Typic Haplusterts, Typic Ustipsamments, Typic Haplustepts and Typic Ustifluvents, respectively, when the water salinity increased from 0.5 to 5.0 dS m-1 (Fig. 3C). A linear increase in the calcium content in pumpkin leaves as a function of the increase of the irrigation water salinity was also verified by [26]. Typic Haplustept soils, which contain a greater amount of clay, are subject to higher accumulation of this ion. The large increase in calcium content in the Typic Ustifluvents can be explained by that soil class’s physical and chemical characteristics, which make it more susceptible to salinization and sodification processes [27].

Source: The Authors.
Figure 3 Contents of magnesium (A), nitrogen (B) and calcium (C) in leaf blades of flowering melon irrigated with water of different salinities and in different soil classes: RQ - Typic Ustipsamments; PVA - Typic Haplustults; CX - Typic Haplustepts; RY - Typic Ustifluvents, and VX - Typic Haplusterts.
3.2 Micronutrients
The micronutrient contents in the leaves of the melon plants presented the greatest variations when the irrigation water salinity increased from 0.5 to 5.0 dS m-1. Only the zinc concentration presented a linear increase in all soils (Fig. 4C). The contents of this nutrient in all soils were within the range considered adequate by [12], which is between 20 and 100 mg kg-1. The lowest content of zinc (35.56 mg kg-1) was found in plants irrigated with water having lowest electrical conductivity (0.5 dS m-1) and cultivated in Typic Haplustepts, while the highest content (104.74 mg kg-1) was observed in plants irrigated with water of highest salinity in Typic Ustipsamments. The increase in water salinity from 0.5 to 5.0 dS m-1 caused increases in foliar zinc contents of 46, 51, 65, 69, and 134% for Typic Ustipsamments, Typic Ustifluvents, Typic Haplustults, Typic Haplusterts and Typic Haplustepts, respectively.
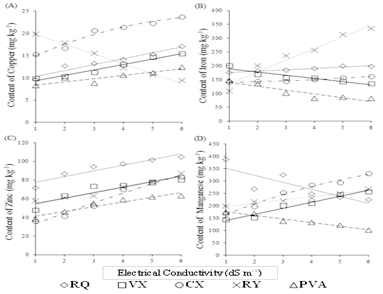
Source: The Authors.
Figure 4 Contents of copper (A), iron (B), zinc (C) and manganese (D) in the leaf blades of flowering melon irrigated with water of different salinities and in different soil classes: RQ - Typic Ustipsamments; PVA - Typic Haplustults; CX - Typic Haplustepts; RY - Typic Ustifluvents, and VX - Typic Haplusterts.
The contents of copper in the leaf tissue of melon plants showed increasing linear behavior as a function of the increase in irrigation water salinity in the different soil classes, except for the Typic Haplustepts, which presented quadratic behavior (Fig. 4A). Among the studied soils, the highest increase (79%) occurred for Typic Ustipsamments, while for Typic Haplusterts, Typic Haplustepts and Typic Haplustults the increases were 56, 56 and 49%, respectively. Contrary behavior was observed in Typic Ustifluvents, in which a reduction of 53 % in the Cu content occurred when the irrigation water salinity increased from 0.5 to 5.0 dS m-1.
In most soils, the leaf contents of this nutrient were within or very close to the range considered adequate by [12], which is between 10 and 15 mg kg-1. However, in the Typic Haplustepts the foliar contents of copper were above the recommended range at all salinities. It should be noted that at the beginning of the experiment, the copper contents in this soil were much higher than in the other soils (Table 1).
The contents of iron in the leaf tissue of melon plants showed increasing linear behavior as a function of the increase in irrigation water salinity in Typic Ustipsamments, Typic Haplustepts and Typic Ustifluvents (Fig. 4B). The increases in these soils were 13, 15 and 204%, respectively. On the other hand, the foliar contents of iron decreased by 32% in the Typic Haplusterts and 44% in the Typic Haplustults. The contents of Fe observed when considering the Typic Ustipsamments, Typic Haplustults, Typic Haplustepts and Typic Ustifluvents are very close to those found by [15], who observed contents between 1 and 3 g kg-1. In most soils, the foliar contents of iron were within the range considered adequate by [12], which is between 50 and 300 mg kg-1. However, for Typic Ustifluvents, the contents found were within that range only up to the electrical conductivity of 3.0 dS m-1.
It is noteworthy that at the beginning of the experiment, the contents of iron in the Typic Ustipsamments and Typic Ustifluvents were higher than in the other soils, but the contents in the melon leaves presented a small increase in the Typic Ustipsamments and a three-fold increase in the Typic Ustifluvents. This is because the Typic Ustifluvents soil is clayey and has a higher natural fertility than the Typic Ustipsamments soil, causing the plants to absorb this ion better when subjected to saline stress.
The contents of manganese in the leaf tissue of melon plants showed increasing linear behavior as a function of the increase in irrigation water salinity in the Typic Haplusterts and Typic Ustifluvents (Fig. 4D). The increases in foliar contents of manganese in these soils were 78 and 35%, respectively. The foliar content of this nutrient in the Typic Haplustepts increased 97%, but in a quadratic pattern. On the contrary, the foliar contents of manganese decreased by 41% in the Typic Ustipsamments and Typic Haplustults. In most soils, the foliar contents of manganese were within the range considered adequate by [12], which is from 50 to 250 mg kg-1. However, in the Typic Haplustepts, the contents of manganese were within that range only up to the water electrical conductivity of 2.0 dS m-1, while in the Typic Ustipsamments the foliar contents of manganese were above that range from the irrigation water salinity of 2.0 dS m-1.
4. Conclusions
Increasing salinity of irrigation water caused a rise in leaf macronutrient content of melon plants in all soils. Increased sodium content indicates the risk to plants and soil of irrigation with saline water. Adequate leaf contents of N, K, and Mg were observed in all soils except K in Typic Haplusterts and Mg in Typic Ustipsamments. Appropriate P levels were observed only in Typic Haplustepts, and Ca only in Typic Haplustults and Typic Ustifluvents.
Increased salinity of irrigation water caused higher leaf contents of micronutrients in plants grown in nearly all soils, with the exception of copper in Typic Ustifluvents, iron in Typic Haplusterts and Typic Haplustults, and manganese in Typic Ustipsamments and Typic Haplustults. Micronutrient contents remained within the appropriate range, with the exception of iron in Typic Ustifluvents and manganese in Typic Haplustepts and Typic Ustipsamments, which had low levels at the highest water salinities.

