










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Biológica Colombiana
Print version ISSN 0120-548X
Acta biol.Colomb. vol.20 no.3 Bogotá Sept./Dec. 2015
https://doi.org/10.15446/abc.v20n3.45421
Doi: http://dx.doi.org/10.15446/abc.v20n3.45421.
Artículo de investigación/Original research paper
TRANSPORTE DE MATERIA ORGÁNICA A LO LARGO DE UN RÍO TROPICAL DE MONTAÑA EN LA SIERRA NEVADA DE SANTA MARTA (COLOMBIA)
Transport of Organic Matter Across a Tropical Mountain Stream of Sierra Nevada De Santa Marta (Colombia)
Cesar Enrique TAMARIS TURIZO1,2, Javier RODRÍGUEZ BARRIOS1
1 Grupo de Investigación en Ecología Neotropical - GIEN. Cra 32 n°. 22-08 Universidad del Magdalena, Santa Marta, Colombia.
2 Grupo de Investigación en Ecología y Biodiversidad Aplicada Cra 32 n°. 22-08 Universidad del Magdalena, Santa Marta, Colombia.
For correspondence. cesartamaris@yahoo.es.
Received: 4th September 2014, Returned for revision: 8th February 2015, Accepted: 21st March 2015.
Associate Editor: Santiago Gaviria Melo.
Citation / Citar este artículo como: Tamaris Turizo CE, Rodríguez Barrios J. Transporte de materia orgánica a lo largo de un río tropical de montaña en la Sierra Nevada de Santa Marta (Colombia). Acta biol. Colomb. 2015;20(3):209-216. doi: http://dx.doi.org/10.15446/abc.v20n3.45421.
RESUMEN
En los ríos tropicales, la materia orgánica es la principal fuente de energía para las comunidades que allí habitan. Las principales proporciones de materia orgánica se encuentra la materia orgánica particulada gruesa (MOPG) y la materia orgánica particulada fina (MOPF). Por lo anterior, en este trabajo se evaluó el transporte de la MOPG, la MOPF y su relación con algunas variables físicas y químicas en tres tramos de un gradiente altitudinal de los 50 msnm hasta los 1700 msnm durante un ciclo diario en el río Gaira (Sierra Nevada de Santa Marta). Para lo cual se realizaron muestreos mensuales entre los meses de marzo a junio del 2008. Se detectaron diferencias significativas en la deriva de MOPG entre las estaciones, siendo la estación 2 (parte media) durante el primer muestreo, la que registró los valores más altos reportados para todo el estudio (474 kg m-3) y la estación 3 mostró la menor biomasa derivante (0.26 kg m-3). La MOPF registró valores similares durante todos los muestreos (0.58 - 6.34 g m-3). Sin embargo, se detectaron diferencias significativas en los muestreos tres y cuatro. Con el aumento de las lluvias, descendió el transporte de MOPG. Una relación inversa entre la velocidad del agua y el transporte de MOPG (rs = -0,70; n = 12; p < 0,05) se registró, mientras que la MOPF, presentó valores constantes. Los resultados de este trabajo coinciden parcialmente con lo registrado en la literatura, lo cual indica que las dinámicas de transporte de MO están relacionadas con las variaciones ambientales locales.
Palabras clave: materia orgánica particulada gruesa, materia orgánica particulada fina, Río Gaira.
ABSTRACT
In tropical streams organic matter is the principal source of energy for the communities in the river. Main components of organic matter are coarse-particulate organic matter (CPOM) and fine particulate organic matter (FPOM). Therefore, transport of CPOM and FPOM and its relationship with some physical and chemical variables was evaluated, on three sections of an altitudinal gradient from 50 to 1700 masl during a daily-cycle at Gaira river. Samples were taken monthly from March to June 2008.-It was observed significant differences in drift of CPOM between stations. Station 2 (middle part), first sample, recorded the highest value (474 kg m-3) and station 3 showed the lowest lower value of drift biomass (0.26 kg m-3) during the study. FPOM showed similar values during all the sampling (0.58 - 6.34 g m-3). However, significant differences were detected between sampling three and four. Transport of FPOM decreased with the increase of precipitation. A inverse relationship between water velocity and transport of CPOM was observed, while the FPOM, showed constant values. These results are partiality similar to reported in the literature, because the dynamic of transport of OM is relationship with local variations environmental.
Keywords: coarse particulate organic matter, fine particulate organic matter, Gaira River.
INTRODUCCIÓN
Las estimaciones del material transportado por la corriente, entre ellas la deriva del material orgánico particulado son un importante descriptor de la dinámica de un río (Wallace et al., 1991; Benfield et al., 2000), debido a que permiten conocer parte de la función del ecosistema acuático en el tramo a evaluar. Una de las variables en los ríos tropicales más influyentes en el transporte de la materia orgánica (MO), es la variación del caudal, la cual puede manifestarse a escala temporal, pero las variaciones periódicas, estacionales o de ciclo diario, se constituyen en condiciones permanentes a las que los organismos han debido responder adaptativamente (Rodríguez-Barrios et al., 2006).
La MO que es transportada a lo largo del río, es un importante recurso energético y de nutrientes, principalmente en la cabecera de los ríos (Mathuriau y Chauvet, 2002; Wallace et al., 2007). La materia orgánica particulada gruesa (MOPG) es generada principalmente por la vegetación ribereña, la cual está constituida principalmente por el aporte de hojas, flores, frutos y ramas (Wallace et al., 1991; Webster et al., 1999; Wantzen y Junk, 2000), también se puede generar en el propio río a través de procesos físicos como la floculación de la materia orgánica particulada fina (MOPF). La MOPG y MOPF transportada en el gradiente altitudinal de un río varía en sus proporciones (Vannote et al., 1980), donde los mayores valores de MOPG se producen en la cabecera y disminuye con el gradiente altitudinal, mientras que la MOPF muestra un comportamiento contrario. Sin embargo, se han desarrollado pocos trabajos en ríos tropicales no permitan validar este comportamiento. El entendimiento de las variables que puedan estar asociadas al transporte de la MO en los ríos tropicales puede ser útil para entender la importancia del aporte de energía de los sistemas autotróficos asociados relacionados con los ríos tropicales, de tal forma que la mayor proporción de MOPG en un sistema puede ser considerado como de fuente alóctona, mientras que la MOPF puede ser alóctona o autóctona.
El material transportado en la parte baja de un río sin llanura aluvial es en mayor proporción resultado de los aportes alóctonos de sus sectores previos (Naiman et al., 1987). Este hecho ha permitido que la cantidad y tipo de material transportado sea usado como un descriptor de procesos de la cuenca (Ramírez y Pringle, 2001). Sin embargo, son pocos los estudios de referencia en este tipo de ecosistemas tropicales, en los cuales se soportan estas hipótesis en condiciones naturales o ambientes no perturbados.
En los ríos de la Sierra Nevada de Santa Marta, el aporte de la MO es un importante suministro energético incluso en la zona baja (Deluque, 2005). Esta energía es aprovechada, en primera instancia, como MOPG por los trituradores, los cuales junto con la acción mecánica de la corriente, ayudan a su transformación a MOPF. Aquí, la materia orgánica es aprovechada principalmente por filtradores y colectores (Wantzen et al., 2008). Además, por ser un sistema montañoso separado de los andes, presenta patrones hidrológicos y climáticos diferentes al andino (Sarmiento, 1987; Prosierra, 1998). De acuerdo a Vannote et al., (1980), se espera que las fracciones de MOPG dominen en la parte alta y media, mientras que la MOPF sea más abundante en la parte baja del río. Caso contrario debe evidenciarse en las cantidades de MOPF detectadas en los diferentes tramos, lo cual debería concordar con los grupos funcionales de las comunidades de macroinvertebrados dominantes en estos sistemas. Por ello, en este estudio se pretende evaluar la dinámica de la MOPG y MOPF que es transportada en un gradiente altitudinal (50 - 1700 m s.n.m.) durante un periodo de bajas precipitaciones, en un río de la Sierra Nevada de Santa Marta.
MATERIALES Y MÉTODOS
Los muestreos se desarrollaron en tres tramos de la cuenca del río Gaira. La Estación 1 ubicada en la parte alta (sector de San Lorenzo) a 1700 m s.n.m., esta localidad se encuentra bien conservada, cerca de esta zona no se realizan actividades agropecuarias extensivas se localiza entre los 11°10'2" N y 74°10'41,5" W. En la vegetación ribereña abundan árboles de gran tamaño que generan una sombra permanente sobre el lecho del río. La Estación 2 (Hacienda La Victoria) a 900 m s.n.m. localizado entre los 11°07'44,2" N y 74°05'35,8" W. En esta zona hay grandes extensiones de cultivos de café. La vegetación ribereña se encuentra poco conservada y se limita a 20 m del río. La Estación 3 (Puerto Mosquito) localizada a 50 m s.n.m. entre los 11°10'26" N, 74°10'37" W. La cobertura de la vegetación es abierta y hay poco bosque ribereño. Cerca de esta estación (aguas arriba) se encuentra una planta de captación de aguas y río abajo hay varias zonas del balneario, convirtiendo al sector en el más intervenido entre las estaciones de estudio. Las características físicas y ambientales de cada estación se describen detalladamente en Tamaris-Turizo et al. (2013).
En cada tramo se realizaron cuatro muestreos, durante los meses de marzo a junio del 2008, lo cual coincidió con la época seca e inicio de las lluvias en la cuenca del río Gaira. En cada estación se realizó la evaluación del ciclo diario en las fechas: 19-23 de marzo (M1), 04-12 de abril (M2), 25 de abril al 3 de mayo (M3) y 11-20 de junio del 2008 (M4).
Recolecta y cuantificación de la MO transportada
Para la recolecta de la MOPG, en cada tramo del río se ubicaron tres trampas de deriva, cada una con un área de 0.045 m2 y un poro de red de 250 μm. Se ubicaron contra corriente permaneciendo sumergidas durante la exposición de las redes. Las trampas se desocuparon cada 12 horas (para evitar su colmatación), hasta completar el ciclo diario de 24 horas. En cada recolecta las muestras se preservaron con alcohol al 80 %, luego se rotularon por separado con información de la estación, fecha y hora.
Para la toma de muestras de la MOPF se llenaron paulatinamente tres botellas plásticas (500 ml) con agua del río con muestras parciales cada 12 horas (en horas del día y la noche). Las muestras se mantuvieron refrigeradas durante el transporte al laboratorio. Este proceso se realizó en todos los tramos.
Se considera MOPG a aquella materia orgánica particulada con un diámetro mayor a 1000 μm y la MOPF con un diámetro entre 1 μm y 1000 μm (Vannote et al., 1980). Sin embargo, en este estudio se evaluó la porción de la MOPF entre 45 μm y 1000 μm, debido a la disponibilidad de los filtros en laboratorio.
Para medir la MOPG (constituida por: hojas, frutos, ramas y fragmentos de organismos). Se retiró del alcohol, luego se enjuagó con abundante agua y se pasó por un tamiz de 1 mm de poro. Posteriormente se llevó a la estufa a 60 ± 1 °C, por 48 horas o hasta obtener peso constante. Finalmente, se calcinó en una mufla a 550 °C durante 2 horas para obtener el peso seco libre de cenizas (PSLC). El transporte de MOPG se calculó utilizando la ecuación de Smock (2007) como se describe a continuación.
D = g / (T V A)
Donde D es densidad de deriva de la MOPG (g PSLC de MOPG x m3), g son los gramos de PSLC de la MOPG, T es el tiempo de exposición de la red (segundos), V es la velocidad de la corriente en la boca de la red (m s-1), A es el área sumergida de la red (m2).
Para la medición de la MOPF, se fi ltraron 100 ml de agua de río por cada muestra mediante vacío utilizando filtros de fibra de vidrio (Whatman con poro de 45 μm). Los fi ltros se pesaron previamente en una balanza analítica (precisión ± 0,1 mg). El producto del filtrado se llevó a la estufa a 105 °C por 24 horas y posteriormente a la mufla a 550 °C durante 2 horas, para conocer su peso final. El transporte de MOPF se calculó de acuerdo a la ecuación de Smock (2007).
D = g / Al
En la cual D es la densidad de MOPF (g PSLC de MOPF Transporte de MOPFx m3) transportada, g son los gramos de PSLC de la MOPF, En la estación 1 se registraron los mayores valores de Al es la alícuota de agua filtrada la cual fue de 0,0001 m-3 en transporte de MOPG, con excepción del M1, en el cual la todas las muestras.
Medición de las variables físicas y químicas
De forma simultánea a la recolección de las muestras de MOPG y MOPF empleando un multiparámetro WTW, se midieron las siguientes variables físicas y químicas: temperatura del agua (°C), pH (unidades), conductividad (μs cm-1) y oxígeno disuelto (mg L-1). Adicionalmente se calculó el caudal en cada tramo del río utilizando un objeto flotador (para calcular la velocidad de la corriente) y con el ancho de la sección. A partir de la composición del lecho en las tres estaciones se estimó que el coeficiente de rugosidad de Manning el cual fue 0,8 (Monsalve, 1999) para corregir los valores de caudal. Cada 12 horas se calculó la velocidad de la corriente en la boca de las redes con ayuda de un objeto flotador.
Análisis de datos
Para evaluar las diferencias estadísticas entre las estaciones de muestreo y entre muestreos, sobre el transporte de MOPG y MOPF, se aplicó la prueba no paramétrica de Kruskal y Wallis (K-W), debido a que los datos no cumplieron con los supuestos de normalidad y/o homogeneidad de varianzas. Las pruebas que presentaron diferencias significativas se evaluaron con la prueba post hoc no paramétrico de Student-Newman-Keuls (SNK). Para conocer las relaciones entre las variables físicas, químicas y el transporte de MOPG y MOPF se una correlación múltiple de Spearman. Todos los análisis se realizaron en los programas Statgraphics Centurion XV, para Windows. Se trabajó con un nivel de significación del 5 %.
RESULTADOS
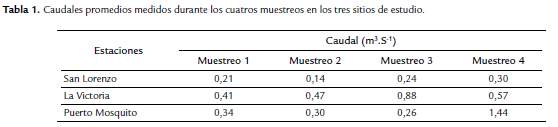
Durante el muestreo 1 (M1), los caudales fueron similares en las tres estaciones; sin embargo, la estación 2 en los tres primeros muestreos registró los mayores valores (0,41- 0,88 m3 s-1), seguido de la estación 3 (parte baja) (0,30- 0,4 m3 s-1), donde se registró el mayor caudal durante el muestreo 4 (M4) (1,44 m3 s-1). La estación 1 (parte alta), registró los menores valores durante el estudio (0,14 - 0,30 m3 s-1) (Tabla 1).
Transporte de MOPG
En la estación 1 se registraron los mayores valores de transporte de MOPG, con excepción del M1, en el cual la estación 2 los presentó. Durante todas las campañas de muestreo, las menores tasas de transporte se evidenciaron en la estación 3, con excepción del M3, donde la menor deriva de MOPG se registró en la estación 2 (Tabla 2). Durante el primer muestreo, la biomasa de MOPG que derivó durante el ciclo diario en la estación 1 fue de 4,10 ± 0,2 kg m-3. En la estación 2 (474,74 ± 92,5 kg m-3) y seguido de una fuerte disminución en la estación 3 (0,34 ± 0,15 kg m-3). Durante el M2 los mayores valores de transporte de la MOPG se presentaron en la estación 1 (140,36 ± 51,8 kg m-3), seguidos de la estación 2 y finalmente la estación 3 con 109,91 ± 35,7 kg m-3 y 18,33 ± 10,1 kg m-3 respectivamente. Este mismo comportamiento se observó durante el M4. Sin embargo, los valores de biomasa transportada fueron muy inferiores a los reportados en los anteriores muestreos (Tabla 2).

Durante el M1 se detectaron diferencias significativas entre la biomasa transportada de MOPG entre las estaciones (p < 0,05; n = 17; K-W = 13,07). El análisis del diagrama de cajas y bigotes muestra que las estaciones 1 y 3 presentaron similares valores y la estación 2 presentó los valores más altos de biomasa y significativamente distintos a las otras dos estaciones (Fig. 1a). Este muestreo se desarrolló en época de sequía o aguas bajas en la zona. En el M2 se evidenciaron diferencias significativas del transporte de MOPG entre las estaciones evaluadas (p < 0.05; n =18; K-W = 7,52); durante el desarrollo de este trabajo en las estaciones 2 y 3 se presentaron algunas precipitaciones. En el diagrama de cajas y bigotes se observan similares valores de biomasa de MOPG en las estaciones 1 y 2, pero la estación 1 es significativamente distinta en un 95% de la estación 3 (Fig. 1b). En el tercer muestreo al igual que en el primero y el segundo, se encontraron diferencias significativas en la deriva de MOPG entre las estaciones (p < 0,05; n = 18; K-W = 9,08). Sin embargo, la estación 1 difirió significativamente de las estaciones 2 y 3 (Fig. 1c). Este mismo comportamiento se evidenció en el M2, en el cual se presentaron fuertes lluvias en la estación 2 (Fig. 1d).

En cuanto al ciclo diario, no se detectaron diferencias significativas entre el transporte de MOPG durante el día y noche (p > 0,05; n = 24; W=0,00). Esto evidencia que los procesos de deriva de MOPG fueron constantes durante los días monitoreados. Sin embargo, durante la mayoría de muestreos los valores de deriva de MOPG aumentaron durante las noches, excepto en las estaciones 2 y 3 en M3 y en la estación 1 durante M4 (Fig. 2).

Transporte de MOPF
El transporte de MOPF total que derivó durante todo el estudio en las estaciones 1, 2 y 3 fue respectivamente: 14,81, 9,32 y 15,11 g m-3. Los mayores valores se registraron en la estación 3, seguida de la estación 1, la estación con el menor valor fue la 2. Sin embargo, esta tendencia no fue constante durante el estudio (Fig. 3). Durante el M1, correspondió al momento de aguas bajas, el mayor transporte de MOPF se dio en la estación 2 con 4,78 g m-3 y el menor en la estación 1 con 1,90 g m-3. Durante el M2, la MOPF transportada en la estación 1 fue 3 veces mayor que en el muestreo anterior (M1) (6,36 g m-3) y 1.5 veces mayor que los muestreos siguientes (M3 y M4) con 3,17 y 3,37 g m-3 respectivamente. La tendencia registrada en la estación 2 durante los muestreos mostró un descenso en el transporte de la MOPF (hasta 0,93 g m-3), a su vez aumentaron las precipitaciones y los niveles de caudal del sector. En la estación 3, la MOPF aumentó gradualmente durante los tres primeros muestreos (2,15 a 6,34 g m-3). Sin embargo, durante el M4 se observó una disminución en este transporte (2,45 g m-3).

Durante los dos primeros muestreos no se detectaron diferencias significativas en el transporte de MOPF entre las estaciones. Este periodo correspondió a época de sequía (Tamaris-Turizo et al., 2013). Sin embargo, en M3 se encontraron diferencias significativas a lo largo del río (p < 0,05; n = 13; K-W = 10,51), con los mayores valores de en la estación 3 (3,19 g m-3), seguida por la estación 1 (3,17 g m-3); finalmente la estación 2 (1,71 g m-3). En M4, los valores más altos de biomasa de MOPF, se reportaron en la estación 1 con 3,37 g m-3, seguido por la estación 3 con 2,45 g m-3 y nuevamente la estación 2 presentó los menores valores (0,93 g m-3). Este periodo hidrológico correspondió al inicio de la época de estabilidad pluviométrica para la zona, en la cual las precipitaciones fueron constantes. La estación 2 presentó los valores más altos de MOPF (3,61 g m-3) durante el M1, hay que anotar que durante este muestreo se presentaron lluvias en esta estación y correspondió a las primeras precipitaciones de gran intensidad luego de la época de sequía.
No se encontraron diferencias (p > 0,05; n = 30 W = 389,5) en el transporte de MOPF durante el ciclo diario, durante ninguno de los muestreos, lo que demuestra que la tasa de transporte de MOPG y MOPF fueron similares durante el día y la noche, y que el leve incremento del caudal que generalmente se presenta al inicio de las horas de la noche, no produce un efecto significativo sobre la deriva de estas fracciones de MO. De igual forma, las variables químicas evaluadas durante este trabajo, no tuvieron efectos significativos sobre el transporte de MOPG y (p > 0,05). Sólo la velocidad tuvo un efecto significativo sobre el transporte de la MOPG (rs = -0,70; n = 12; p < 0,05).
DISCUSIÓN
La velocidad tuvo un efecto significativo sobre el transporte de la MOPG (rs = -0,70; n = 12; p < 0,05), lo que sugiere que las variables hidráulicas influyen en el transporte de este material (Wilcox et al., 2008). Esto significa que tal como se evidenció en este estudio no es suficiente evaluar el aumento de los caudales en términos de cantidad de agua que pasa por una sección, si no de enfocar los estudios realizados sobre las comunidades biológicas en las fuerzas físicas que ejerce el fluido sobre los organismos y la capacidad que éstos tienen a resistir dichas fuerzas (Allan y Castillo, 2007). Las variables químicas no tuvieron efectos significativos sobre el transporte de MOPG (p > 0,05).
Transporte de MOPG y MOPF
La biomasa MOPG yMOPF detectadas en este estudio durante los dos primeros muestreos fue elevada, especialmente por la biomasa de MOPG derivada en la estación 2 durante M1. Esta deriva estuvo influenciada por las precipitaciones que se dieron durante el muestreo, ya que la MOPG que se encontraba almacenada en el lecho del río, piedras, ribera y deltas, fue arrastrada luego de unas lluvias finalizando la época bajas precipitaciones, lo que produce aumento en los niveles del caudal y arrastre del material, como lo sugieren Gonçalves et al. (2006), Ríos (2008) y Rodríguez-Barrios (2008). Así mismo, Cano (2003) en un estudio de trasvase de una quebrada en Medellín, encontró significancia en la variación del caudal sobre el transporte de MOPG y MOPF. Sin embargo, Ríos (2008), en la cabecera de un río andino ecuatoriano, reportó que los mayores valores de deriva de MOPG se dieron durante la época de aguas bajas, aunque los datos registrados no fueron estadísticamente diferentes a la época de lluvias.
Durante los siguientes muestreos la parte alta del río presentó la mayor biomasa derivante de MOPG, las cuales fueron significativamente diferentes a las demás estaciones, verificando de esta manera que las mayores fracciones de MOPG se dieron en la cabecera de los ríos y disminuyeron con el gradiente altitudinal (Vannote et al., 1980). Por otro lado, el transporte de MOPF no presentó esta tendencia clara, ya que el mayor transporte se dio en la estación 3, pero seguido de la estación 1 y finalmente las menores tasas en la estación 2. Sin embargo, durante M1 la mayor deriva de MOPF se dio en la estación 2; lo que puede significar que esta dinámica puede estar relacionada con otras variables no evaluadas en este estudio, tales como la pendiente, composición del suelo y las precipitaciones locales y puntuales como lo sugieren Bilby y Likens (1979) para un río en New Hampshire (USA). Aunque, Rodríguez-Barrios et al. (2008) encontraron que el transporte de la MOPF se encuentra asociado con la dinámica de las precipitaciones; por tanto, las precipitaciones no sincrónicas que se dan en el río Gaira (Sierra Nevada de Santa Marta) pueden influir sobre la dinámica de deriva de la MO.
Para conocer la dinámica del transporte de MO, es necesario cubrir los meses correspondiente a la temporada de lluvias en la zona (septiembre - noviembre), puesto que en este estudio se muestreo sólo en la época de aguas bajas e inicio de las precipitaciones en el sector. Los datos de MOPG reportados en este estudio fueron muy elevados comparados con los reportados por Ríos (2008), Rodríguez-Barrios (2008). Sin embargo, fueron tres veces más altos a la mayor tasa de deriva reportada por Deluque (2005), la cual fue de 117,7 kg d-1 en la parte baja del río Gaira, durante el mes de enero; a pesar de todo, en este mes se presentaron fuertes precipitaciones en este año (Tabla 3). El transporte de MOPF fue un poco mayor al registrado por Rodríguez-Barrios et al. (2008). Pero en este estudio se evidencian altas tasa de deriva de MOPF en todas las estaciones y tiempos de muestreo (marzo-junio).

CONCLUSIONES
De acuerdo a lo encontrado en este estudio, el río Gaira se ajusta a los criterios descrito por Vannote et al. (1980) en cuanto al transporte de MO a lo largo del río debido a: 1) La mayor cantidad de biomasa de MOPG transportada se presentó en la estación 1, ubicada en la cabecera del río. 2) La velocidad de la corriente presentó una relación inversa con el transporte de MOPG, lo cual implica que esta variable física es de mucha importancia en el transporte de MO y por ende de energía en los sistemas acuáticos. 3) la deriva de la MOPG y la MOPF disminuyó con el aumento de las precipitaciones, lo cual debe estar relacionado con el lavado de MO que ocurre con las primeras lluvias.
AGRADECIMIENTOS
A la División Financiera de la Universidad Nacional de Colombia y a la Universidad del Magdalena por la financiación del proyecto. A la Reserva Natural La Iguana Verde y La Hacienda Cafetera La Victoria por facilitar sus instalaciones como estaciones de muestreo. A los integrantes de los Grupos de Investigación en Limnología Neotropical y Ecología Neotropical por el apoyo en campo. A Gabriel Pinilla y John Jairo Ramírez por la revisión del documento.
REFERENCIAS
Allan JD, Castillo MM, editors. Stream Ecology-Structure and function of running waters. Second edition. Netherland: Springer; 2007. p. 388. [ Links ]
Benfield EF, Webster JR, Hutchens JJ, Tank JL, Turner PA. Organic matter dynamics along a stream order and elevational gradient in a Southern Appalachian stream. Verh Internat Verein Limnol. 2000;27(1):1341-1345. [ Links ]
Bilby RE, Likens GE. Effect of hydrologic fluctuation on the transport of fine particulate organic carbon in a small stream. Limnol Oceanogr. 1979;24(1):69-75. Doi: 10.4319/lo.1979.24.1.0069. [ Links ]
Cano W. Alteración del metabolismo lótico en la Quebrada Piedras Blancas (Medellín) debido al trasvase del caudal (Tesis de Maestría en Biología). Medellín: Instituto de Posgrados, Universidad de Antioquia; 2003. p. 140. [ Links ]
Deluque Y. Deriva de macroinvertebrados bentónicos y la materia orgánica gruesa en la cuenca baja del río Gaira (Sierra Nevada de Santa Marta, Colombia) (Tesis de pregrado). Santa Marta: Universidad del Magdalena, Facultad de Ciencias Básicas, Programa de Biología; 2005. p. 36. [ Links ]
Gonçalves JF, Silva J, Callisto M. Dynamics of allochthonous organic matter in a tropical Brazilian headstream. Braz Arch Biol Technol. 2006;49(6):967-973. Doi: 10.1590/S1516-89132006000700014. [ Links ]
Mathuriau C, Chauvet E. Breakdown of leaf litter in a neotropical stream. J N Am Benthol Soc. 2002;21(3):384-396. Doi: 10.2307/1468477. [ Links ]
Monsalve G. Hidrología en la Ingeniería, Escuela Colombiana de Ingeniería. 2nd Edition. Bogotá: Escuela Colombiana de Ingeniería; 2004. p. 382. [ Links ]
Naiman RJ, Melillo JM, Lock MA, Ford TE, Reice SR.Longitudinal patterns of ecosystem processes and community structure in a subarctic river continuum. Ecology. 1987;68(5):1139-1156. Doi: 10.2307/1939199. [ Links ]
Prosierra-Fundación Pro-Sierra Nevada de Santa Marta. Evaluación Ecológica Rápida de la Sierra Nevada de Santa Marta. Definición de Áreas Críticas para la Conservación de la Sierra Nevada de Santa Marta. Colombia: Ministerio del Medio Ambiente, UAESPPNN The Nature Conservacy -USAID- Embajada de Japón; 1998. p. 134. [ Links ]
Ramírez A, Pringle C. Spatial and temporal patterns of invertebrate drift in streams draining a Neotropical landscape. Freshw Biol. 2001;46(1):47-62. Doi: 10.1111/j.1365-2427.2001.00636.x. [ Links ]
Ríos B. Comunidades de macroinvertebrados en un río altoandino: Importancia del hábitat, dinámica de la deriva, papel de la materia orgánica y relevancia de la ovoposición. (Tesis Doctoral). Barcelona, España: Departament d'Ecología, Universitat de Barcelona; 2008. p. 154. [ Links ]
Rodríguez-Barrios J, Ospina-Torres R, Gutiérrez JD. Entradas y transporte de materia orgánica en una quebrada tropical de montaña. Bogotá, Colombia. Caldasia. 2008;30(2):349-367. [ Links ]
Rodríguez-Barrios J, Ospina-Torres R, Berrío M, Cepeda B, Castellanos G, Valencia M. Variación diaria de la deriva de macroinvertebrados acuáticos y de materias orgánica en la cabecera de un río tropical de montaña en el departamento de Nariño, Colombia. Acta Biol. Colomb. 2006;11(supl.1):47-53. [ Links ]
Sarmiento G. Los principales gradientes ecoclimáticos en los Andes tropicales. En: Universidad de Medellín, editor. Medellín: Anales del IV Congreso Latinoamericano de Botánica, Medellín; 1987. p. 47-64. [ Links ]
Smock L. Macroinvertebrate movements: drift, colonization, and emergence. In: Hauer, FR, Lamberti GA, editors. Methods in stream ecology. 2th edition. China: Academy Press; 2007. p. 466. [ Links ]
Tamaris-Turizo CE, Rodríguez-Barrios J, Ospina-Torres R. Deriva de macroinvertebrados acuáticos a lo largo del río Gaira, vertiente noroccidental de la Sierra Nevada de Santa Marta, Colombia. Caldasia. 2013;35(1):149-163. [ Links ]
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE. The River Continuum Concept. Can J Fish Aquat Sci. 1980;37(1):130-137. Doi: 10.1139/f80-017. [ Links ]
Wallace JB, Hutchens JJ, Grubaugh JW. Transport and storage of MOPF. In: Hauer FR, Lamberti GA, editors. Methods in stream ecology. China: Academy Press; 2007. p. 249-272. [ Links ]
Wallace JB, Cuffney TF, Webster JR, Lugthart GL, Chung K, Goldowitz BS. Export to fine organic particles from headwater streams: effects of season, extreme discharges and invertebrate manipulation. Limnol Oceanogr. 1991;36(4):670-682. Doi: 10.4319/ lo.1991.36.4.0670. [ Links ]
Wantzen KM, Yule CM, Mathooko JM. Organic matter processing in tropical streams. In: Dudgeon D editor. Tropical Stream Ecology. USA: Academic Press; 2008. p. 44-60. [ Links ]
Wantzen KM, Junk WJ. The importance of stream-wetland-systems for biodiversity: a tropical perspective. In: Gopal B, Junk WJ, Davies JA, editors. Biodiversity in wetlands: assessment, function and conservation. Leiden: Backhuys Publishers; 2000. p. 11-34. [ Links ]
Webster JR, Benfield EF, Ehrman TP, Schaeffer MA, Tank JL, Hutchens JJ, et al. What Happens to Allochthonous material that falls into streams? A synthesis of new and published information from coweeta. Fresh Biol. 1999;41(4):687-705. Doi: 10.1046/j.1365-2427.1999.00409.x. [ Links ]
Wilcox AC, Peckarsky BL, Taylor BW, Encalada AC. Hydraulic and geomorphic effect on mayfly drift in high-gradient streams at moderate discharges. Ecohydrol. 2008;1(2):176-186. Doi: 10.1002/eco.16. [ Links ]
