











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Cucúrbita moschata (Duch. ex Lam.) Duch. ex Poir. es la especie domesticada del género Cucúrbita más cultivada y consumida en varios departamentos de Colombia. Esta especie es un cultivo importante para la seguridad alimentaria (Restrepo y Vallejo, 2008), pues posee entre otras bondades un alto valor nutritivo debido a la presencia de β-caroteno (provitamina A), carotenos totales, ácido ascórbico, carbohidratos, proteínas y minerales (Restrepo, 2015).
En la especie C. moschata no existe un consenso acerca del tipo de acción génica predominante en la expresión y control genético de la variable PFP en cruzamientos dialélicos. En cruzamientos entre progenitores S0, Espitia et al. (2006) y Ortiz et al. (2013) reportan acción génica aditiva; Ruiz et al. (2004) y Toro (2009) sustentan acción génica no aditiva; Ortiz et al. (2013) y Nisha y Veeraragavathatham (2014) documentaron que ambos efectos fueron importantes. En cruzamientos dialélicos entre líneas endogámicas S1 se ha reportado efectos aditivos y no aditivos (de los genes) como responsables de la expresión y control genético de PFP (Mohanty, 2000; Espitia et al., 2006; Jha et al., 2009; Pandey et al., 2010; Ortiz et al., 2013; El-Tahawey et al., 2015; Begum et al., 2016), mientras que Rana et al. (2015) sustenta sólo acción génica no aditiva. En cruzamientos dialélicos entre líneas endogámicas S2, también se ha registrado la importancia de ambos efectos en la expresión de PFP (Ortiz et al., 2013).
Para PPF en C. moschata tampoco existe un consenso sobre el tipo de acción génica predominante en la expresión y control genético de la variable. En cruzamientos entre progenitores S0, Gwanama et al. (2001) y Espitia (2004) hallaron efectos aditivos; Ruiz et al. (2004) y Nisha y Veeraragavathatham (2014) sustentaron efectos aditivos y no aditivos. En cruzamientos dialélicos entre líneas endogámicas S1, Espitia (2004), Jha et al. (2009), El-Tahawey et al. (2015) y Begum et al. (2016) detectaron efectos aditivos y no aditivos; mientras que Mohanty (2000) reportó solo efectos no aditivos de los genes responsables en la expresión de PPF.
Los objetivos específicos de este estudio fueron: a) estimar los efectos de habilidad combinatoria general y específica en la expresión de la producción por planta y el peso promedio del fruto, en tres cruzamientos dialélicos de Cucurbita moschata, cada uno con diferente grado de endogamia (S0, S1, S2). b) Identificar progenitores o híbridos F1 sobresalientes por sus efectos de habilidad combinatoria y por el comportamiento promedio de sus variables.
MATERIALES Y MÉTODOS
La investigación se realizó en el Centro Experimental de la Universidad Nacional de Colombia, sede Palmira (CEUNP), localizado en Candelaria, Valle del Cauca (Colombia), con coordenadas 3°25'34,42'' N y 76°25'47,57'' W 980 msnm, 26°C de temperatura promedia anual, 1.100 mm de precipitación promedia anual y 76% de humedad relativa media.
Se evaluaron tres cruzamientos dialélicos de zapallo C. moschata, conformados cada uno por seis progenitores con diferentes niveles de endogamia (progenitores a libre polinización S0, líneas endogámicas S1 y S2, derivadas de los progenitores S0). Los progenitores 50 fueron: UNAPAL-Abanico-75-1, UNAPAL-Abanico-75-2, UNAPAL-Dorado, IC3A, UNAPAL-Llano-grande-1 y UNAPAL-Llanogrande-2, los cuales fueron seleccionados por sus características sobresalientes tamaño y color externo del fruto, color y grosor de la pulpa y origen geográfico diverso. Los frutos fueron seleccionados utilizando un índice de selección ponderado, en el cual se incluyeron las variables peso promedio del fruto (2,0-4,0 kg), grosor de pulpa (3,5-5,0 cm) y color salmón de la pulpa. En cada cruzamiento dialélico se evaluaron 21 genotipos (seis progenitores y 15 cruzamientos directos).
La evaluación agronómica de los tres cruzamientos dialélicos se realizó de manera conjunta en un solo experimento durante el segundo semestre de 2011. Se utilizó un diseño anidado de dos etapas con cuatro repeticiones. Se evaluaron dos factores: factor generaciones (factor A) y factor genotipos (factor B). El factor A conformado por tres niveles (generación S0, generación S1 y generación S2) y el factor B constituido por 63 niveles (21 genotipos S0 + 21 genotipos 51 + 21 genotipos S2). Los 21 genotipos S0 quedaron anidados bajo el nivel generación S0, 21 genotipos S1 quedaron anidados bajo el nivel generación S1 y 21 genotipos S2 quedaron anidados bajo el nivel generación S2. La distancia de siembra fue de 2,5X3,0 m entre surcos y entre plantas, respectivamente. La parcela experimental consistió de un surco de cinco plantas (37,5 m2). Como parcela útil se utilizaron las tres plantas centrales. Se evaluaron las variables producción de frutos por planta (PFP en kg) y peso promedio del fruto (PPF en kg).
El análisis genético y estadístico para estimar la habilidad combinatoria de los diferentes genotipos, se realizó empleando el método propuesto por Hallauer y Miranda (Hallauer et al., 2010). El modelo estadístico fijo asociado al diseño experimental fue:

Donde:
i, j |
: 1, 2....... p padres; p = 6; |
k |
: 1, 2......r repeticiones; r = 4; |
µ |
: efecto de la media general del experimento; |
g¡ |
: efecto de habilidad combinatoria general del progenitor "i"; |
gj |
: efecto de habilidad combinatoria general del progenitor "j"; |
Sij |
: efecto de habilidad combinatoria específica del híbrido entre los progenitores "i" x "j"; |
bk |
: efecto del bloque k; |
Dl |
: efecto de la generación de endogamia l; |
(Db)lk |
: efecto de la interacción de la generación de endogamia por bloques; |
(1/rn) ΣΣ eijkl |
: error experimental asociado con la observación Yijkl; |
Yijkl |
: valor promedio fenotípico observado de la variable en estudio para el cruzamiento (ij) en la generación de endogamia l. |
El análisis de varianza y la estimación de los efectos genéticos para las variables de interés, se realizaron mediante el uso de los programas computacionales SAS® (paquete SAS/STAT®, versión 9.4 del sistema SAS® para Windows©) de SAS Institute Inc.© (2012) y GENES (versión Windows©, 2004.2.1) de Cruz (2013). Para algunas fuentes de variación del análisis de varianza se utilizó la prueba F y para los efectos genéticos la prueba de T de Student.
RESULTADOS Y DISCUSIÓN
Análisis de varianza
Se observaron diferencias significativas entre las generaciones de endogamia S0, S1 y S2 de C. moschata para las variables producción por planta (PFP) y peso promedio del fruto (PPF), indicando que al menos una de las generaciones resultó ser significativamente diferente del resto (Tab. 1). Resultados similares fueron publicados en generaciones de endogamia S0, S1 y S2 de C. moschata para PFP (Ortiz et al., 2013). Igualmente, se detectaron diferencias significativas en la fuente de variación genotipos en cada una de las tres generaciones endogámicas evaluadas, y por ende, se puede inferir que existe al menos un padre o un híbrido que presentó un promedio de PFP y PPF diferente estadísticamente del resto de promedios en cada una de dichas generaciones (Tab. 1). Resultados similares fueron reportados en C. moschata para PFP (Espitia et al., 2006) y PPF (Espitia, 2004) en generaciones de endogamia S0 y S1.
Tabla 1 Cuadrados medios (CM) del ANDEVA para las variables producción de frutos por planta (PFP) y peso promedio del fruto (PPF) en tres generaciones de endogamia de C. moschata, según el método de Hallauer y Miranda (Hallauer et al., 2010).

*, **: Significativos al 5 y 1% de probabilidad, respectivamente, según la prueba de F. HCG: habilidad combinatoria general; HCE: habilidad combinatoria específica; CV: coeficiente de variación.
En la fuente de variación progenitores no se encontraron diferencias estadísticas en la generación de endogamia S0 para PFP y PPF, lo cual se debió posiblemente a que en este trabajo se utilizó un índice de selección que incluyó entre otras variables el PPF, de tal manera que se seleccionaron frutos de los seis progenitores S0 con valores similares de esta variable (2,0-4,0 kg).
Este rango de selección corresponde a valores óptimos de PPF para la comercialización de frutos dirigidos al mercado de consumo en fresco conformado por consumidores que prefieren comprar el fruto entero y no en rodajas (Tab. 1). Estos resultados contrastan con los registrados en generaciones de endogamia S0 de C. moschata para PFP (Espitia et al., 2006; Ortiz et al., 2013) y PPF (Espitia, 2004). En la generación de endogamia S1 si se presentaron diferencias significativas entre progenitores para PFP y PPF (Tab. 1). Resultados similares fueron reportados en generaciones de endogamia S1 de C. moschata para PFP (Ortiz et al., 2013) y PPF (Espitia, 2004); mientras que diferencias no significativas fueron publicadas para PFP por Espitia et al. (2006). En la generación S2 solo se registraron diferencias para PPF (Tab. 1); mientras que Ortiz et al. (2013), si registraron diferencias para PFP.
La significancia estadística de los cuadrados medios de PFP y PPF para la fuente de variación cruzas, en las tres generaciones, demuestran que al menos uno de los cruzamientos entre progenitores S0 o entre líneas S1, o entre líneas S2, presentó comportamiento promedio de PFP o PPF diferente del resto (Tab. 1). De lo anterior se infiere que independientemente del nivel de endogamia, es probable encontrar al menos un híbrido con un promedio de PFP o PPF significativamente diferente del resto.
La habilidad combinatoria general (HCG) en la generación de endogamia S0 fue significativa para PFP y PPF; sin embargo, la habilidad combinatoria específica (HCE) no fue significativa. La HCG y HCE en la generación de endogamia S1, fueron altamente significativas para PFP y PPF. En la generación S2, la HCG fue altamente significativa para PFP y PPF; mientras que la HCE solo fue estadísticamente significativa para PPF. Para el contraste padres vs cruzas (P vs. C), en las tres generaciones de endogamia, se observaron diferencias altamente significativas en PFP y PPF, indicando que el comportamiento promedio de todos los cruzamientos F1 (entre progenitores S0 y entre líneas S1 o S2) fue superior al comportamiento promedio de los progenitores en conjunto, señalando efectos heteróticos significativos en los tres cruzamientos dialélicos para PFP y PPF (Tab. 1). Resultados similares fueron reportados en generaciones S0 y S1 de C. moschata para PFP y PPF (Espitia et al., 2006; Espitia, 2004) y en generaciones S0, S1 y S2 para PFP (Ortiz et al., 2013).
En el cruzamiento dialélico entre progenitores S0 solo los efectos aditivos (HCG) fueron importantes en la expresión genética de PFP (Tab. 1). Resultados similares fueron reportados en C. moschata por, Espitia et al. (2006) y Ortiz et al. (2013), quienes encontraron que la acción génica aditiva fue importante en la expresión genética de PFP en cruzamientos dialélicos entre progenitores S0. En contraste, en cruzamientos dialélicos entre progenitores S0 de C. moschata se ha publicado que sólo la acción génica no aditiva fue importante (Ruiz et al., 2004; Toro, 2009). Por otro lado, en esta especie en un cruzamiento dialélico entre progenitores S0 para PFP se ha registrado que ambos efectos fueron importantes (Ortiz et al., 2013; Nisha y Veeraragavathatham, 2014).
En el cruzamiento dialélico entre líneas endogámicas S1 la HCG y HCE, indicaron la importancia de la acción génica aditiva y no aditiva en la expresión y control genético de la PFP (Tab. 1). Resultados similares fueron registrados por Mohanty (2000), Espitia et al. (2006), Jha et al. (2009), Pandey et al. (2010), Ortiz et al. (2013), El-Tahawey et al. (2015) y Begum et al. (2016) en C. moschata, los cuales encontraron que los efectos aditivos y no aditivos de los genes fueron responsables de la expresión de PFP en cruzamientos dialélicos entre líneas endogámicas S1. Contrariamente, Rana et al. (2015) publicaron que sólo la acción génica no aditiva fue la importante en la expresión de PFP.
En el cruzamiento dialélico entre líneas endogámicas S2 solo los efectos aditivos (HCG) fueron importantes para PFP (Tab. 1). Estos resultados contrastan con los registrados por Ortiz et al. (2013), quienes observaron que ambos efectos fueron estadísticamente significativos para PFP en cruzamientos dialélicos entre líneas endogámicas S2 de C. moschata.
En el cruzamiento dialélico entre progenitores S0 de C. moschata para la expresión y control genético del PPF solo los efectos genéticos aditivos fueron importantes (Tab. 1). Resultados similares fueron reportados por Gwanama et al. (2001) y Espitia (2004) en C. moschata, quienes encontraron que solo los efectos aditivos fueron importantes para esta variable en el cruzamiento dialélico entre progenitores S0. En contraste, para PPF también se ha reportado la importancia de los efectos aditivos y no aditivos (Ruiz et al., 2004; Nisha y Veeraragavathatham, 2014).
En los cruzamientos dialélicos entre líneas endogámicas S1 y S2 la HCG y HCE fueron importantes en PPF, indicando la importancia de la acción génica aditiva y no aditiva en su expresión genética (Tab. 1). Espitia (2004), Jha et al. (2009), El-Tahawey et al. (2015) y Begum et al. (2016), también encontraron en líneas endogámicas S1 de C. moschata la importancia de efectos aditivos y no aditivos de los genes en la expresión de PPF. Por otro lado, Mohanty (2000) al realizar un cruzamiento dialélico entre líneas S1 en C. moschata, reportó solo efectos no aditivos de los genes como los responsables de la expresión de PPF.
Al realizar un análisis global entre generaciones de endogamia se puede deducir, que los efectos aditivos fueron importantes en la expresión de PFP y PPF en todas las generaciones endogámicas evaluadas; mientras que los efectos no aditivos de los genes solo fueron importantes en los cruzamientos dialélicos entre líneas endogámicas en comparación con el cruzamiento dialélico entre progenitores S0 para estas variables. Este mismo comportamiento ha sido reportado por Espitia (2004) y Espitia et al. (2006) en C. moschata para esas variables, comparando el cruzamiento dialélico entre progenitores S0 con el cruzamiento dialélico entre líneas endogámicas S1. Resultados iguales han sido reportados en maíz por varios autores (Crossa et al., 1990; Rezende y Souza Junior, 2000), los cuales dan como una posible explicación a esto, la mayor divergencia genética que puede presentarse en los cruzamientos entre progenitores de base genética estrecha (S1 y S2) comparada con la que se presenta en cruzamientos entre progenitores de base amplia (S0).
Heterosis promedia para PFP y PPF
La heterosis promedia en las generaciones de endogamia S1 y S2 fue significativa para PFP y PPF; no así para la generación S0 (Tabs. 2 y 3). La mayor expresión de la heterosis promedia detectada en este estudio, en el dialelo entre líneas endogámicas S1 y S2 en comparación con el dialelo de progenitores S0, ha sido explicada por otros autores como Hallauer et al. (2010), los cuales han reportado que las progenies de líneas endogámicas pueden presentar una mayor heterosis con respecto a progenies de poblaciones de base genética amplia, cuando se utilizan como progenitores líneas genéticamente divergentes y se presentan niveles de dominancia unidireccional en la mayoría de loci que codifican por el carácter que se quiere mejorar.
Tabla 2 Producción por planta (PFP) para progenitores (diagonal), híbridos (arriba de la diagonal), efectos de habilidad combinatoria especifica (sij) (debajo de la diagonal), efectos de habilidad combinatoria general (gi) y promedio de los híbridos de zapallo C. moschata, en los cruzamientos dialélicos entre progenitores S0 y entre líneas endogámicas S1 y S2.

*, **: Significativos al 5% y 1 % de probabilidad, respectivamente, según la prueba de T de Student.
Tabla 3 Peso promedio del fruto (PPF) para progenitores (diagonal), híbridos (arriba de la diagonal), efectos de habilidad combinatoria específica (sij) (debajo de la diagonal), efectos de habilidad combinatoria general (gi) y promedio de los híbridos de zapallo C. moschata, en los cruzamientos dialélicos entre progenitores S0 y entre líneas endogámicas S1 y S2.
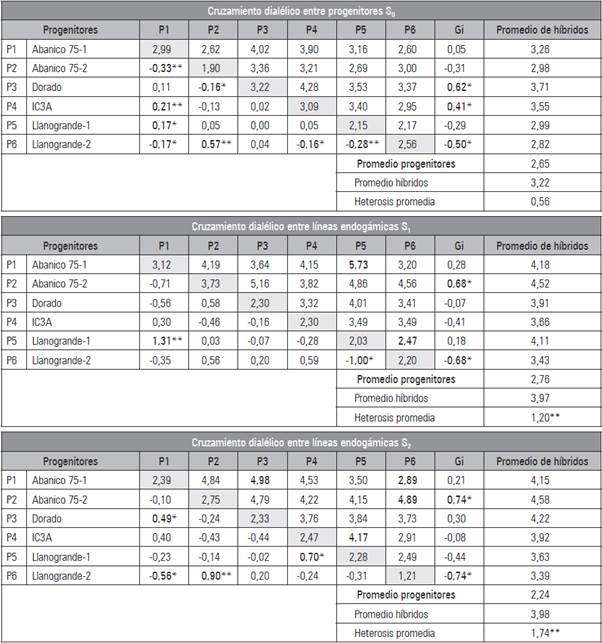
*, **: Significativos al 5% y 1 % de probabilidad, respectivamente, según la prueba de T de Student.
Efectos de habilidad combinatoria para PFP
Para PFP ninguno de los progenitores S0 o de las líneas endogámicas S1 y S2, presentaron efectos de HCG significativos (Tab. 2), lo cual fue debido posiblemente a que en este trabajo de investigación la producción de las diferentes generaciones de endogamia estuvo acompañada de selección, donde los frutos seleccionados de los progenitores S0 o de las líneas endogámicas S1 y S2, tenían pesos promedios intermedios (2,0-4,0 kg), y por ende, no necesariamente correspondían a individuos con las mayores PFP. Algunas investigaciones reportaron que ningún progenitor S0 presentó efectos de HCG significativos para PFP en C. moschata (Ruiz et al., 2004; Toro, 2009) y en ninguna línea endogámica S1 (Rana et al., 2015). Otras investigaciones registraron al menos un progenitor S0 con efectos de HCG significativos (Espitia et al., 2006; Ortiz et al., 2013; Nisha y Veeraragavathatham, 2014). Contrariamente a lo observado en este estudio, Mohanty, (2000), Espitia et al. (2006), Jha et al. (2009), Pandey et al. (2010), Ortiz et al. (2013), El-Tahawey et al. (2015) y Begum et al. (2016) encontraron al menos una línea endogámica S1 con efectos de HCG estadísticamente significativos para PFP en C. moschata. Por otro lado, Ortiz et al. (2013) tampoco encontraron líneas endogámicas S2 con efectos de HCG significativos para dicha variable.
Para PFP ningún cruzamiento registró efectos de HCE significativos (Tab. 2). Las causas podrían ser la falta de complementación genética entre los progenitores que participaron en los cruzamientos o la carencia de niveles importantes de divergencia genética, aunque ésta última causa podría ser la menos posible, pues los progenitores S0 tienen un origen geográfico diferente (IC3A: Costa Rica-Centroamérica; UNAPAL-Abanico-75-1 y UNAPAL-Abanico-75-2: Magdalena; UNAPAL-Dorado: Valle del Cauca; UNAPAL-Llano-grande-1 y UNAPAL-Llanogrande-2: Cauca). En contraste a lo observado en este estudio, en progenitores S0 (Ruiz et al., 2004; Toro, 2009; Nisha y Veeraragavathatham, 2014) y en líneas endogámicas S1 (Mohanty, 2000; Espitia et al., 2006; Jha et al., 2009; Pandey et al., 2010; Ortiz et al., 2013; Rana et al., 2015; El-Tahawey et al. 2015; Begum et al., 2016), encontraron al menos un híbrido con efectos de HCE estadísticamente significativos para PFP. Por otro lado, Ortiz et al. (2013) encontraron un híbrido de líneas endogámicas S2 con efectos de HCE estadísticamente significativos para esta variable.
Efectos de habilidad combinatoria para PPF
El comportamiento diferencial de los progenitores para PPF en sus valores como progenitor (gi) (Tab. 3), confirman las diferencias altamente significativas detectadas para HCG en el ANDEVA en los tres cruzamientos dialélicos (Tab. 1) y favorece la selección de nuevos genotipos homocigotos o progenitores superiores, dado que los genes con efectos aditivos son los que responden a la selección. Gwanama et al. (2001), Ruiz et al. (2004), Espitia (2004) y Nisha y Veeraragavathatham (2014), también encontraron en C. moschata al menos un progenitor S0 con valores de gi (efectos de HCG) estadísticamente significativos para PPF. Igualmente, Espitia (2004), Jha et al. (2009) y El-Tahawey et al. (2015) reportaron en C. moschata varias líneas endogámicas S1 con efectos de HCG diferentes de cero para PPF. Por otro lado y con respecto a los cruzamientos dialélicos entre líneas endogámicas S1 y S2, los resultados de los efectos de HCE (Sij) para PPF (Tab. 3), también confirman los niveles de significancia estadística del ANDEVA (Tab. 1).
Similarmente, Mohanty (2000), Espitia (2004), Jha et al. (2009) y Begum et al. (2016) encontraron en C. moschata algunos híbridos entre líneas endogámicas S1 con efectos de HCE estadísticamente diferentes de cero para PPF.
Genotipos recomendados para el mejoramiento genético de PPF
El progenitor más indicado para mejorar PPF con destino al mercado de consumo en fresco constituido por consumidores que prefieren comprar frutos enteros y no en rodajas, mediante el aprovechamiento de los efectos aditivos por selección recurrente intrapoblacional, es el progenitor S0 UNAPAL-Dorado, debido a que presentó un valor en los efectos de HCG significativo (0,62**), un promedio en sus híbridos de PPF con un valor óptimo para ser comercializado como fruto entero (3,71 kg) (Tab. 3) y un promedio en sus híbridos de PFP aceptable (14,05 kg/planta) (Tab. 2). Por otro lado, el progenitor recomendado para mejorar PPF, mediante el aprovechamiento de los efectos aditivos y no aditivos por selección recurrente intrapoblacional, dirigido al mercado de consumo en fresco y uso agroindustrial, conformado por aquellos consumidores en los cuales el peso del fruto no es una característica limitante para su adquisición, es la línea endogámica S2 de UNAPAL-Abanico-75-2, pues registró un valor en los efectos de HCG significativo (0,74*), un promedio en sus híbridos de PPF de 4,58 kg (Tab. 3) y el promedio en sus híbridos de PFP más alto de todos los progenitores evaluados (16,93 kg/ planta) (Tab. 2).
El híbrido más indicado para mejorar PPF mediante el aprovechamiento de los efectos aditivos y no aditivos por selección recurrente recíproca, dirigido al mercado de consumo en fresco conformado por consumidores que prefieren comprar frutos enteros y no en rodajas, es el híbrido entre líneas endogámicas S2 UNAPAL-Abanico-75-1 x UNAPAL-Llanogrande-2, debido a que presentó un valor en los efectos de HCE significativo (-0,56*), un valor óptimo de PPF para ser comercializado como fruto entero (2,89 kg) (Tab. 3) y un valor de PFP aceptable (14,00 kg/planta) (Tab. 2). Por otro lado, los híbridos recomendados para el mejoramiento de PPF a través de la explotación de los efectos aditivos y no aditivos por selección recurrente recíproca, con destino al mercado de consumo en fresco y uso agroindustrial, constituido por aquellos consumidores en los cuales el peso del fruto no es una característica limitante para su adquisición, son el híbrido entre líneas endogámicas S1 UNAPAL-Abanico-75-1 X UNAPAL-Llanogrande-1 y el híbrido entre líneas endogámicas S2 UNAPAL-Abanico-75-1 X UNAPAL-Dorado. Ambos híbridos presentaron valores en los efectos de HCE significativos (1,31** y 0,49*, respectivamente), valores de PPF de 5,73 y 4,98 kg, respectivamente (Tab. 3); así como también los valores de PFP más altos entre todos los híbridos evaluados (20,50 y 20,87 kg/planta, respectivamente) (Tab. 2).
Los valores de los efectos de HCE negativos y significativos para algunos de los híbridos, sugiere que su promedio fue inferior al promedio esperado con base en la gi de sus progenitores y la media general, considerándose por ello como una alternativa genética viable para reducir el tamaño o peso promedio del fruto y así satisfacer las preferencias del mercado fresco de zapallo.
CONCLUSIONES
En la generación de endogamia S0 solo la habilidad combinatoria general fue importante en la expresión de PFP y PPF. En contraste, en la generación de endogamia S1 tanto la habilidad combinatoria general como la habilidad combinatoria específica, contribuyeron al control genético de PFP y PPF.
En la generación de endogamia S2, las habilidades combinatorias general y específica fueron responsables del control de la expresión del PPF; mientras que en PFP solo la habilidad combinatoria general fue importante.
Los genotipos más indicados para mejorar PPF con destino al mercado de consumo en fresco constituido por consumidores que prefieren comprar frutos enteros y no en rodajas, son el progenitor S0 UNA-PAL-Dorado y el híbrido entre líneas endogámicas S2 UNAPAL-Abanico-75-1 x UNAPAL-Llanogrande-2.
Los genotipos recomendados para mejorar PPF para el mercado de consumo en fresco y uso agroindustrial, conformado por aquellos consumidores en los cuales el peso del fruto no es una característica limitante para su adquisición, son la línea endogámica S2 de UNAPAL-Abanico-75-2, el híbrido entre líneas endogámicas S1 UNAPAL-Abanico-75-1 x UNAPAL-Llanogrande-1 y el híbrido entre líneas endogámicas S2 UNAPAL-Abanico-75-1 x UNAPAL-Dorado.


