










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkActa Agronómica
Print version ISSN 0120-2812
Acta Agron. vol.65 no.1 Palmira Jan./Mar. 2016
https://doi.org/10.15446/acag.v65n1.47167
http://dx.doi.org/10.15446/acag.v65n1.47167
Hormigas asociadas a nectarios extraflorales de árboles de dos especies de Inga en cafetales de Cauca, Colombia
Ants asociated to extrafloral nectaries of two species of Inga trees in coffee plantations of Cauca, Colombia
Rosa Maria Sinisterra 1, María Cristina Gallego-Ropero *1 e Inge Armbrecht 2
1 Departamento de Biología Universidad del Cauca, Popayán, Colombia Grupo GEA y GEAHNA. 2 Departamento de Biología Universidad del Valle, Cali, Colombia Grupo GEAHNA. *Autora para correspondencia: mgallego@unicauca.edu.co
Rec.: 10.11.2014 Acep.: 01.07.2015
Resumen
En dos fincas cafeteras localizadas en la vereda Villanueva, municipio de Popayán; Colombia, se identificó la comunidad de hormigas presentes en los nectarios extraflorales de árboles de guamo (Inga edulis e I. densiflora) utilizados como sombra en cafetales, y sobre la hojarasca del suelo. Las recolecciones se realizaron con cebos, extracción de hojarasca con sacos mini-Winkler y captura manual. Se encontraron 43 especies, distribuidas en 17 géneros y siete subfamilias. Los resultados mostraron que las especies de hormigas que habitan en los árboles son diferentes a las que habitan en la hojarasca, pero gracias a sus hábitos forrajeros algunas especies se desplazan de un estrato al otro. Se concluye que los árboles de sombrío como el guamo son importantes tanto para el mantenimiento de las interacciones mutualistas hormiga–planta, como para la conservación de la diversidad; además, la presencia de árboles puede contribuir indirectamente a la prestación de servicios ecosistémicos fitosanitarios, ya que algunas especies de hormigas de los géneros Crematogaster y Camponotus pueden servir como potenciales agentes de control biológico para insectos plaga.
Palabras clave: Diversidad, interacción, hormigas, NEFs, sombra.
Abstract
The ant communities in the extrafloral nectaries of shade Inga edulis e I. densiflora (‘guamo’) trees and in the leaf litter of two coffee farms at Villanueva village, Popayán municipality, were identified. Bait collection, extraction of leaf litter by means of mini-Winkler and direct manual capture were carried out. A total of 43 ant species, distributed in 17 genera and seven subfamilies was found. Results show that ant species that inhabit trees are different from ant species than leaf litter dwelling ants. However, thanks to their foraging habits, some ant species may pass from one to the other stratum. It is concluded that shade trees, such as guamo trees, are important both to maintain mutualistic ant-plant interactions and to promote diversity conservation. Additionally, the presence of shade trees may indirectly contribute to provide phytosanitary ecosystem services given that some ant species, such as Crematogaster and Camponotus, might serve as potential biological control agents for pest insects.
Keywords: Interaction, ants, diversity, shade, EFNs.
Introducción
Las interacciones ecológicas entre plantas y hormigas (Hymenoptera: Formicidae) son resultado de procesos evolutivos que han modelado la historia de vida de estos organismos, permitiéndoles establecer relaciones específicas de diversa índole como depredación, competencia y simbiosis (Itino et al., 2001). Estas dinámicas ecológicas de las comunidades en interacción con factores biogeográficos, resultan en las distribuciones espaciales observadas en zonas tropicales (Davidson y Mckey, 1993) y por tanto, es interesante describir los mecanismos o relaciones interespecíficas que explican dichas distribuciones.
No siempre las interacciones que involucran hormigas son de tipo mutualista, también pueden ser oportunistas o casuales, sin que esto signifique que sean menos importantes bajo la óptica humana. La asociación simbiótica mutualista se establece cuando tanto la hormiga como la planta se benefician. Las hormigas protegen las plantas contra herbívoros y plantas invasoras, brindan nutrientes esenciales, dispersan semillas, y en ocasiones, polinizan las plantas (Cushman y Beattie, 1991). Por su parte, las plantas ofrecen a las hormigas dos tipos de recompensa, un sitio de anidación y una fuente de alimento (Cushman y Beattie, 1991; Bronstein, 1998). Las hormigas desempeñan un papel muy importante en el mantenimiento de la dinámica natural de los ecosistemas y pueden ser usadas como agentes de control biológico de plagas (Vandermeer Perfecto, Ibarra-Nuñez, Philpott, y García-Ballinas, 2002, Ibarra – Núñez García, J., y Moreno, 1990, Perfecto Rice y Vandervoor, 1996; Barbera et al., 2004; Gallego-Ropero y Armbrecht, 2005).
Los nectarios extraflorales son glándulas que producen secreciones azucaradas que atraen hormigas, pero su función no se relaciona con la polinización sino más bien con el ofrecimiento de una recompensa a estos insectos a cambio de protección contra herbívoros (Cushman y Beattie. 1991; Bronstein, 1998). Estas estructuras poseen variaciones morfológicas y anatómicas y pueden ser encontradas sobre algunas partes de la planta tales como la lámina foliar, pecíolo, raquis, estípulas, tallo, brácteas, pétalos, sépalos, frutos y cotiledones (Delabie, Ospina y Zabala, 2003). Los nectarios extraflorales se presentan en 93 familias de angiospermas y 332 géneros. Estas plantas se pueden encontrar en zonas templadas y tropicales, siendo más comunes en estas últimas (Koptur, 1992).
El néctar que producen es rico en carbohidratos como fructuosa, sacarosa y glucosa, pero también tiene proteínas, aminoácidos, ácidos orgánicos, lípidos, alcaloides, fenoles, vitaminas y saponinas (Bentley, 1977). Las principales subfamilias de hormigas reconocidas que son atraídas por los nectarios extraflorales son Myrmicinae, Formicinae y Dolichoderinae (Del Claro, Bert y Reú , 1996).
Los árboles de Inga spp. (Tribu Ingeae, subfamilia Mimosoideae, Leguminosae), conocidos como ‘guamos’, son los más frecuentemente utilizados como sombrío en los cultivos de café en la zona andina colombiana, por tanto, es importante explorar los servicios ecológicos directos e indirectos que a través de las hormigas proveen estos árboles. Es posible que la tradición de sembrarlos tenga fundamento en ventajas todavía no conocidas claramente. En ocasiones el agricultor se ve beneficiado sin saberlo de varias formas, una es la protección microclimática para su cultivo, otras son la protección de la erosión del suelo y el enriquecimiento de nutrientes en el suelo a través del colchón de hojarasca. Quizás el beneficio más desapercibido es la presencia de biota benéfica como aves, murciélagos, arañas y hormigas, entre otros. Las hormigas constituyen uno de los elementos faunísticos más conspicuos y ubicuos en los agroecosistemas tropicales, y los árboles de Inga no son la excepción, muchas de estas hormigas, que anidan y se alimentan de nectarios extraflorales, son también depredadoras de otros artrópodos, por lo cual los arboles de Inga podrían estar prestando un beneficios adicional en los cafetales de sombra a través del mantenimiento de poblaciones viables de estas hormigas benéficas. El objetivo de esta investigación fue, por tanto, reconocer la comunidad de hormigas presentes en los nectarios extraflorales de dos especies de Inga (Fabaceae: Mimosoideae), examinar su composición y su relación con la diversidad de hormigas presentes en la hojarasca de cultivos de café, en la vereda Villanueva, Popayán, Colombia. Además se busca reconocer la importancia de las interacciones entre las hormigas y estos árboles en los agroecosistemas de café como potenciales agentes de control biológico.
Materiales y métodos
Área de estudio
El estudio se realizó en el corregimiento La Rejoya ubicado a 18 km al occidente del municipio de Popayán, en la vereda Villanueva, entre 1600 y 1800 m.s.n.m., con una temperatura media entre 18 y 24°C y un promedio anual de lluvias entre 2000 y 4000 mm, que corresponde a la zona de vida bosque húmedo y muy húmedo premontano (bh-PM y bmh-PM) (Holdridge, 1977). Esta zona se seleccionó por presentar un área de producción en café significativa para la región. La investigación se desarrolló en dos fincas que presentan las características siguientes:
- Finca El Ensueño. Se encuentra a 2° 32’ 19.9” N y 76° 36’ 49.2” O, a 1745 m.s.n.m. Tiene una extensión de 1.5 ha, cultivada con café variedad Caturra con una densidad de siembra de 3000 plantas/ha. Los árboles de sombrío son guamo (I. edulis e I. densiflora) con una densidad de 123 árboles/ha. Además, tiene cultivos de plátanos, bananos y una cerca viva de plantas de fique (Furcraea andina (Agavaceae)).
- Finca El Progreso. Se encuentra a 2° 32’ 04.8” N y 76° 36’ 55” O, a 1745 m.s.n.m. Tiene una extensión de 2 ha. Está cultivado con plantas de café variedad caturra con una densidad de siembra de 3000 plantas/ha. Los árboles utilizados para sombrío son guamos (I. edulis e I. densiflora) con una densidad de 100 árboles/ha. Tiene plantaciones de plátano, banano, chachafruto y como cerca viva botón de oro (Tithonia diversifolia). Ambas fincas han sido manejadas durante 10 años con sistemas de agricultura orgánica (Figura 1).

Muestreo de hormigas
En cada uno de los cultivos de café en ambas fincas fueron escogidos al azar cinco árboles de guamo, los cuales estaban sembrados a 10 m de distancia uno de otro. Por su escogencia aleatoria, este número de árboles representa una muestra válida para los cafetales de sombra en la región. En cada árbol se realizaron las labores y observaciones siguientes: (1) selección y demarcación de una rama para observación de los nectarios extraflorales (4 a 5 por rama) por 20 min y recolección de hormigas visitantes. Este procedimiento se realizó tres veces a diferentes horas cada día de muestreo con el fin de obtener una muestra representativa de la actividad de las hormigas durante el día. En total, fueron 5 horas de muestreo por día por cafetal x 10 muestreos. (2) instalación en los árboles de cebos con atún y miel (recursos atractivos para las hormigas) a una altura desde el suelo de 120 cm. Las hormigas atraídas fueron recolectadas 4 horas después (modificado Villareal et al., 2004). (3) recolección de hojarasca presente en 1 m2, utilizando un marco colocado a 50 cm de la base de cada árbol. La hojarasca recolectada fue pasada a través de un saco cernidor y procesada en sacos mini-Winkler que extraen los insectos por gravedad y geotaxismo, por 48 horas (Bestelmeyer et al., 2002). Las recolecciones fueron realizadas cada 15 días entre julio y noviembre de 2009. Las hormigas fueron identificadas con la ayuda de claves e ilustraciones de revisiones taxonómicas de Palacio y Fernández (2003), Mackay y Mackay (2002) y por comparación con ejemplares de la colección del Museo de Entomología de la Universidad del Valle (MEUV) y del Museo de Historia Natural de la Universidad del Cauca (MHNUC).
Análisis de datos
Todos los datos registrados se organizaron en una matriz de abundancias y se generó un listado con todas las especies encontradas para cada salida de campo. Se realizó un análisis de la información utilizando el programa EstimateS 6.00 (Colwell, 1997) para producir curvas de acumulación de especies y determinar la confiabilidad del muestreo, a través de estimadores no paramétricos ampliamente utilizados como CHAO1 y ACE que analizan la riqueza específica cuando se obtienen valores de abundancia. Igualmente se calcularon los índices de diversidad y similaridad.
Resultados y discusión
En total fueron recolectados 2613 especímenes, distribuidos en 17 géneros, 43 especies y siete subfamilias: Myrmicinae, Ponerinae, Dolichoderinae, Formicinae, Pseudomyrmicinae, Ectatomminae y Ecitoninae (Tabla 1), siendo la subfamilia Myrmicinae la que registró el mayor número de géneros (9) y especies (29) con los tres métodos empleados. En la finca El Progreso se registró 60% del total de las especies encontradas y en la finca El Ensueño se encontró 76% del total, con superposición de varias especies en ambas fincas. Según los estimadores CHAO1 y ACE (Colwell, 1997) se obtuvo una eficiencia de muestreo entre el 77% y 85% con las tres técnicas empleadas.
En la finca el Ensueño se registró la presencia de Procryptocerus hyaleus, hormiga solitaria, oportunista, arbórea, asociada a sitios con un buen nivel de sombra, concordando con lo reportado por Gallego-Ropero, Montoya y Armbrecht, (2009) para las especies de este género presentes en cafetales con sombra. Estas hormigas no son abundantes ni dominantes, pero son habitantes del dosel de selva tropicales (Longino y Snelling, 2002). La especie se registró una vez forrajeando sobre la hojarasca, lo que sugiere que caen del follaje arbóreo o que bajan al suelo ocasionalmente. Sobre la hojarasca se registraron las hormigas Neivamyrmex sp. y Wasmannia auropuctata.
En la finca el Progreso se registraron como especies exclusivas de hojarasca Gnamptogenys striatula, Octostruma sp., Pyramica sp.2 y Odontomachus chelifer y en el estrato arbóreo la hormiga Temnothorax sp. que se caracteriza por poseer hábitos variados de forrajeo, arbóreas, con colonias pequeñas y crípticas (Longino, 2004).
Cuando se calculó el índice de disimilaridad se observa que las fincas se diferencian en índice de 65%, por tanto, se puede sugerir que por el tipo de manejo agrícola que se realiza en cada una de ellas, se presentan condiciones habitacionales diferentes que permiten albergar una particular diversidad de hormigas. Así mismo, el índice de Shannon fue significativamente diferente entre las fincas (p: 0.001), siendo mayor para la finca El Ensueño (Ensueño 2.28 vs. Progreso 1.45). Esta diferencia posiblemente fue debida a la complejidad estructural que ocasiona la mayor densidad de siembra de árboles de guamo, que aportan abundante materia orgánica y sitios de anidación, lo que genera condiciones ambientales que favorecen la disponibilidad de recursos para varias especies de hormigas. En cambio, la finca El Progreso posiblemente fue afectada por los cambios en la temperatura, ya durante el año del estudio disminuyeron las precipitaciones, es decir, se extendió la época seca (Fenómeno de El Niño) y el sombrío de árboles era menor.
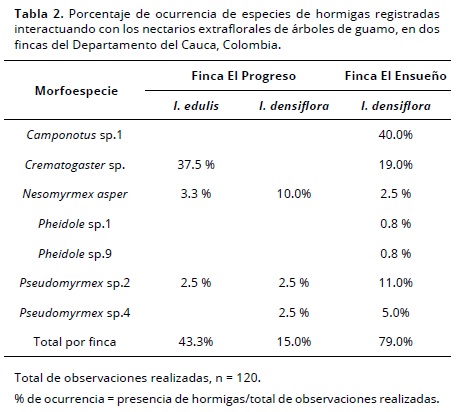
Sobre los nectarios extraflorales se encontraron géneros como Camponotus, Crematogaster y Pseudomyrmex que concuerdan con algunos géneros encontrados por varios autores sobre diferentes especies vegetales con nectarios extraflorales como Bixa orellana, Passiflora sp., Triumfeta semitriloba (Bentley, 1977; Apple y Feener, 2001; Sobrinho et al., 2002), y sobre la hojarasca los géneros como Hypoponera, Pyramica, Pheidole y Solenopsis.
El género Inga pertenece a la subfamilia Mimosoideae, conformado por aproximadamente 350 especies descritas. Muchas de estas especies tienen nectarios extraflorales. En este género, las hojas son compuestas y paripinnadas, con glándulas nectaríferas con forma de copa localizadas entre cada par de foliolos, frecuentemente con raquis alado. La inflorescencia puede variar desde umbela a capítulo y los frutos son legumbres indehiscentes alargados (Gentry, 1993). Las especies encontradas en el sitio de estudio fueron I. edulis Mart conocida como guamo santafereño o rabo de mono e I. densiflora Benth conocida como guaba salada (Nova-Serrano, 1992).
La hormiga Crematogaster sp. fue la que más interacciones registró con I. edulis y para el caso de I. densiflora fue Pseudomyrmex sp.2 en la finca El Progreso. En la finca El Ensueño la hormiga que frecuentemente visitó los nectarios fue Camponotus sp.1 y Crematogaster sp. para I. densiflora. Este hallazgo concuerda con lo observado por Koptur (1984), quien registra que hay más de una especie de hormiga asociada a cada especie de Inga y estas hormigas que visitan los nectarios se desplazan por el follaje. Así mismo, Marín-Cárdenas y García (2008) registran a Pseudomyrmex boopis y Cephalotes sp., hormigas de hábitos arbustivos, sobre nectarios de I. edulis en sistemas ganaderos.
El género Crematogaster fue el más abundante en las dos fincas estudiadas (40%) y su actividad de forrajeo alcanzó los dos estratos: arbóreo y hojarasca. Además los ejemplares fueron recolectados con frecuencia sobre los nectarios extraflorales (Tabla 2). Este género ha sido registrado como potencial agente de control biológico sobre la mayor plaga del café, un curculiónido escolítino conocido como ‘broca del café’ (Bustillo et al. 2002). Otros géneros encontrados en las fincas forrajeando en los diferentes estratos fueron Solenopsis, Pheidole, Odontomachus, Neivamyrmex, Hypoponera, Azteca, Gnamptogenys, Linepithema y Pseudomyrmex (Tabla 1)) que han sido reportados como potenciales agentes de control biológico (Van Mele,Cuc y Van Huis 2001; Rico-Gray y Oliveira, 2007). Se sabe que en cafetales las hormigas también pueden establecer relaciones con trofobiontes (Mera-Velasco, Gallego-Ropero y Armbrecht, 2010) que a su vez podrían llegar a ser plaga del cultivo. No obstante, existen dos argumentos que permiten discutir que los árboles de sombra tienden a proveer más beneficios que perjuicios en este sentido. Por un lado, cuando los nectarios extraflorales de Inga sp. proveen suficiente carbohidratos en forma de azúcares, las colonias se dedican a buscar más fuentes de proteína y se potencian como controladores biológicos (Carabalí-Banguero, Wyckhuys, Montoya-Lerma, Kondo y Lundgren 2013); por otro lado, la reducción de la sombra de árboles genera un desbalance en las comunidades de hormigas, que por lo general no favorecen a aquellas que son buenas controladoras de la broca del café, como lo demostró Philpott en México cuando las podas de árboles afectaron negativamente las hormigas benéficas (Philpott, 2005).
Por otro lado, la estratificación vertical fue marcada, como lo demostró el hecho que la identidad de las hormigas que se encontraron en hojarasca fue diferente a aquellas forrajeando en los árboles de guamo y utilizando los nectarios extraflorales (Figura 2). Las diferencias en identidad se pueden atribuir, en parte a cierta fidelidad que varias especies de hormigas guardan al estrato donde anidan, sin descartar que puedan pasar a otro estrato. El resultado de las especies compartidas entre estratos es consistente con lo encontrado por Ramírez Herrera y Armbrecht (2010) en cafetales del Cauca, quienes observaron que L. neotropicum frecuentaba ambos estratos verticales. Sin embargo, no son consistentes con los hallazgos de Armbrecht (2003) en cafetales de Risaralda, donde el género Procryptocerus frecuenta árboles y anida en ramas. En ocasiones se registran especies arbóreas en hojarasca cuando han caído obreras o sus nidos. Las hormigas encontradas en hojarasca en este estudio han sido reportadas ampliamente en este estrato (Gallego-Ropero, 2005), por ej., W. auropunctata es muy abundante en este estrato, aunque puede anidar en arbustos y grietas de árboles (Arcila, Chacón de Ulloa y Achury, 2008; Achury, Chacón de Ulloa, y Arcila, 2012).

Conclusiones
- En este estudio, se encontró que las especies vegetales empleadas como árboles de sombrío en el cultivo de café, brindan recursos alimenticios y sitios de anidación para el establecimiento de las comunidades de hormigas, por lo cual pueden estar brindando servicios indirectos adicionales al agricultor.
- Las especies de hormigas que frecuentan los nectarios extraflorales tienden a ser diferentes de las que se encuentran en la hojarasca lo cual, posiblemente, obedece al estrato donde anidan; no obstante también ocurren especies que frecuentan el suelo y los árboles moviendo recursos a través de los estratos verticales.
- De acuerdo con los resultados, se sugiere la conservación de especies vegetales empleadas como árboles de sombrío en los cafetales pues son esenciales para el mantenimiento de un ensamblaje de insectos benéficos, el establecimiento de redes tróficas y el mantenimiento de la diversidad biológica.
Agradecimientos
A los dueños de las fincas El Progreso y El Ensueño y sus familias por permitirnos desarrollar el trabajo, al Laboratorio de Entomología de la Universidad del Valle dónde se corroboraron las identificaciones de las hormigas, a la Universidad del Cauca y a la Universidad del Valle.
Referencias
Achury, R., Chacón de Ulloa, P., y Arcila, A. (2012). Effects of the heterogeneity of the landscape and the abundance of Wasmannia auropunctata on ground ant assemblages in a Colombian tropical dry forest. Psyche: A Journal of Entomology. 1 - 12. [ Links ]
Apple, J., y Feener, D. H. (2001). Ants visitation of extrafloral nectaries of Passiflora: the effects of nectary attributes and ant behavior on patterns in facultative mutualism. Oecologia 127, 409 - 416. [ Links ]
Arcila, A., Chacón de Ulloa, P., y Achury, R. 2008. Composición de hormigas e interacciones competitivas con Wasmannia auropunctata en fragmentos de bosque seco tropical. Rev. Col. Entom. 34(2), 209 - 216. [ Links ]
Barbera, N.; Hilje, L.; Hanson, P.; Longino, J.; Carballo, M; y De Melo, E. 2004. Diversidad de especies de hormigas en un gradiente de cafetales orgánicos y convencionales. Rev. Manejo de Plagas y Agroecologíata 72:60 - 71. [ Links ]
Bentley, B. 1977. Extrafloral nectaries and protection by pugnacious bodyguards. Ann. Rev. Ecol. Syst. 8:407 - 427. [ Links ]
Bestelmeyer, B.; Angosti, D.; Alonso, L.; Brandão, C; Brown, W.; Delabie, J.; y Silvestre, R. 2002. Field techniques for the study of ground dewellings ants: An over view, description and evaluation. In: Ants, standard methods for measuring and monitoring biodiversity. Smithsonian Institution Press. Washington, USA. p. 122 - 269. [ Links ]
Brandão, C. (2007). Prólogo. In: Jiménez, E.,Fernández, F., Arias, T., y Lozano, F. (eds.). IX Sistemática, biogeografía y conservación de las hormigas cazadoras de Colombia Bogotá, Colombia: Instituto De Investigación de Recursos Biológicos Alexander von Humboldt. [ Links ]
Bronstein, J. (1998). The contribution of ant plant protection studies to our understanding of mutualism. Biotropica 30(2),150 - 161. [ Links ]
Bustillo, A., Cardenas, R., y Posada, F. (2002). Natural enemies and competitors of Hypothenemus hampei (Ferrari) coleóptera scolytidae in Colombia. Neotrop. Entom. 31(4), 635 - 639. [ Links ]
Carabalí-Banguero, D. J., Wyckhuys, K. A. G., Montoya-Lerma, J., Kondo, T., y Lundgren. (2013). Do additional sugar sources affect the degree of attendance of Dismicoccus brevipes by the fire ant Solenopsis geminata? Entom. Exp. et Appicata. 148, 65 - 73.
Colwell, R. 1997. EstimateS: Statical Estimation of Species Richness and Shared Species from Samples. Version 6.1 User's Guide and application published at http://viceroy.eeb.uconn.edu/estimates. [ Links ]
Cushman, J. H. y Beattie, A. J. (1991). Mutualism: assessing the benefits to host and visitors. Tree 6(6). 193 - 195. [ Links ]
Davidson, D. W., y Mckey, D. (1993). The evolutionary Ecology of symbiotic ant – plant relationships. J. Hymen. Res. 2,13 – 83. [ Links ]
Delabie, J. C., Ospina, M., y Zabala, G. (2003). Relaciones entre las hormigas y las plantas: una introducción. En: Introducción a las hormigas de la región tropical. Fernández (ed.). Bogotá. Colombia: Instituto de Investigación Alexander Von Humboldt. 167 - 180.
Del claro, K., Bert, B., y Reú, W. (1996). Effect of of herbivore deterrence by ants on the fruit set ant extrafloral nectary plant, Qualea multiflora (Vocchysiaceae). J. Tropical Ecol. 12,887 - 892. [ Links ]
Gallego-Ropero, M. C., y Armbrecht, I. (2005). Depredación por hormigas sobre la broca del café Hypothenemus hampei (Curculionidae: Scolytinae) en cafetales cultivados bajo dos niveles de sombra en Colombia. Manejo Integrado de Plagas y Agroecología, (76) 32 - 40. [ Links ]
Gallego-Ropero, M. C., Montoya, J., y Armbrecht, I. (2009). ¿Es la sombra benéfica para la diversidad de hormigas y peso del café? Una experiencia en Pescador, Cauca, Colombia. Bol. Cient. Mus. Hist. Nat. 13(2),106 - 116. [ Links ]
Gentry, A. (1993). A field guide to the families and genera. Woody plants on Northwest south Americ. Conservation Intenational. Washington D.C. 8795 p. [ Links ]
Itino, T., Davies, S., Tada, H., Hieda, Y., Inoguchi, M., Itioka, T., Yamane, S., e Inouye.T. (2001). Cospeciation of ants and plants. Ecol. Res. 16,787 - 793. [ Links ]
Ibarra–Nuñez, G., García, J., y Moreno, M. (1990). Diferencias entre un café orgánico y uno convencional en cuanto a diversidad y abundancia de dos grupos insectos. En: Memorias primera conferencia internacional IFOAM sobre café orgánico. Universidad Autónoma de Chapingo. México, p. 115 - 129. [ Links ]
Koptur, S. (1984). Experimental evidence for defense of Inga (mimosoideae) saplings by ants. Ecology. 65(6),1787 - 1793. [ Links ]
Koptur, S. (1992). Extrafloral nectary â mediated interactions between insects and plants. In: EA. Bernays (ed.). insect's plant interactions. Boca Ratón. FL; CRC Press. 4:81- 129 [ Links ]
Longino, J. T.(2004). Picture guide to the ants genera of Costa Rica. 129 Recuperado de: http://academic.evergreen.edu/projects/ants/genusguide/poncommon.html [ Links ]
Longino, J. T., y Snelling. R. R. (2002). A taxonomic revision of the Procryptocerus (Hymenoptera: Formicidae) of Central America. Contributions in science, Natural History Museum of Los Angeles County. 495:1- 30. [ Links ]
Mackay, W. P., y Mackay, E. E. (2002). Ants of New Mexico; Hymenoptera: Formicidae. The Edwin Mellen Press, Lewiston, NY. p. 408. [ Links ]
Marín-Gómez, O. H., y García-Cárdenas, R. (2008). Artropofauna asociada al guamo Inga edulis (Fabales: Mimosaceae) en un agroecosistema ganadero del Quindío, Colombia. Rev. Asoc. Col. Cien. Biol. 20,117 - 129. [ Links ]
Mera-Velasco, Y. A., Gallego-Ropero, M. C., y Armbrecht, I. (2010). Asociaciones entre hormigas y otros insectos en follaje de cafetales de sol y sombra, Cauca Colombia. Rev. Col. Entom. 36,116 - 126. [ Links ]
Nova-Serrano, O. (1992). Crecimiento inicial de guaba salada (Inga densiflora Benth) y guaba chillillo (Inga edulis) y guaba machete (Inga spectabilis) en dos sitios de Costa Rica. (Tesis de Maestria). CATIE, Turrialba, Costa Rica.119 p. [ Links ]
Palacio, E y Fernández, F. 2003. Claves para subfamilias y géneros. En: Fernández. F. (ed.). Introducción a las hormigas de la región Tropical. Bogotá, Colombia: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. p. 233 - 259. [ Links ]
Perfecto, I., Rice, R., Greenberg, R., y Van der Voort, M. E. 1996. Shade coffee: a disappearing refuge for biodiversity. Bioscience 46:598-608. [ Links ]
Philpott, S. M. (2005). Changes in arboreal ant populations following pruning of coffee shade-trees in Chiapas, Mexico. Agrof. Syst. 64, 219 - 224. [ Links ]
Ramírez, M., J. Herrera, y Armbrecht, I. (2010). Hormigas que depredan en potreros y cafetales colombianos: ¿bajan de los árboles? Rev. Col. Entom. 36 (1),106 - 115. [ Links ]
Rico-Gray, V., y Oliveira, P. (2007). The ecology and evolution of ant-plant interactions. Chicago, United States: The University of Chicago Press. Pp 331 . [ Links ]
Sobrinho, T.; Schoereder, J.; Rodrigues, L.; y Collevatti, R. 2002. Ant visitation (Hymenoptera) to extrafloral nectaries increases seed set and seed viability in the tropical weed Triumfetta semitriloba. Sociobiology, 39(2),353 - 368. [ Links ]
Vandermeer, I., Perfecto, I., Ibarra-Nuñez, G., Philpott, S., y García-Ballinas, J. A. (2002). Ants (Azteca sp.) as potencial biological control agents in shade coffee production in Chiapas, Mexico. Agrof. Syst, 56, 271 - 276. [ Links ]
Van Mele, P., y Nguyen, T. (2001). Farmers' perceptions and practices in use of Dolichoderus thoracicus (Smith) (Hymenoptera: Formicidae) for biological control of pests of sapodilla. Biological Control, 20(1), 23 - 29. [ Links ]
Van Mele, P., Cuc, N., y Van Huis, A. (2001). Farmers' knowledge, perceptions and practices in mango pest management in the Mekong Delta, Vietnam. Intern. J. Pest Manag. 47(1),7 - 16. [ Links ]
Villareal, H., Álvarez, S., Córdoba, F., Escobar, G., Fagua, F., Mendoza, H., Ospina, M., y Umaña, A. M. (2004). Manual de métodos para el desarrollo de inventarios de biodiversidad. Programa de inventarios de biodiversidad-Insectos. Bogotá, Colombia: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Ppp. 1541 - 183. [ Links ]
