











 (pdf)
(pdf)




 SciELO
SciELO  Google
Google  SciELO
SciELO  Google
Google 
 Permalink
PermalinkActa Agronómica
ISSN 0120-2812
https://doi.org/10.15446/acag.v65n1.48161
http://dx.doi.org/10.15446/acag.v65n1.48161
Respuestas fisiológicas de Theobroma cacao L. en etapa de vivero a la disponibilidad de agua en el suelo
Physiological responses of Theobroma cacao L. to water soil available in nursery stage
Jairo García Lozano 1* y Liz Patricia Moreno Fonseca 2
1Facultad de Ingeniería Agronómica, Departamento de Producción y Sanidad Vegetal Universidad del Tolima, Colombia. 2Universidad Nacional de Colombia sede Bogotá, Colombia. *Autor para correspondencia: jairogl2004@gmail.com
Rec.:09.01.2015 Acep.: 18.03.2015
Resumen
En el estudio se evaluó el efecto del déficit de agua en el potencial hídrico foliar y el intercambio de gases de plantas de tres clones de cacao (Theobroma cacao L.) en etapa de vivero. El experimento se estableció en un diseño de parcela dividida con arreglo de bloques aleatorizados y cinco repeticiones. La parcela principal correspondió a cuatro niveles de agua disponible en el suelo y la subparcela a plántulas injertadas de tres meses de edad. Los resultados mostraron diferencias altamente significativas (P < 0.01) para los contenidos de agua en el suelo, pero no se encontraron diferencias entre los materiales evaluados. La pérdida de agua en el suelo disminuyó el potencial hídrico foliar (Ψf) y ocasionó el cierre de estomas alterando el intercambio de gases. El déficit de presión de vapor (DPV) se acentuó principalmente en horas del mediodía cuando aumentó la evapotranspiración. La magnitud del impacto del déficit hídrico depende de las variaciones climáticas a lo largo del día. Las variables climáticas que afectaron el desarrollo de la planta fueron la temperatura y la humedad relativa en forma de DPV. La fotosíntesis neta y el crecimiento de las plántulas de cacao son variables fisiológicas muy sensibles al exceso y especialmente al déficit de agua.
Palabras clave: Estrés hídrico, sequía, fotosíntesis, transpiración, potencial hídrico, cacao.
Abstract
In the locality of El Espinal, Tolima, the effect of water stress on leaf water potential and gas exchange of plants three clones of cacao (Theobroma cacao L) was evaluated. The experiment was established in a split plot design in randomized block arrangement. The main plot was four levels of available soil water, subplot grafted seedlings to three months of three clones with five repetitions. The results showed highly significant differences (P <0.01) in content of soil water, but no differences between the materials evaluated. The loss of water in the soil decreases leaf water potential (Ψf) and causes stomatal closure altering gas exchange and vapor pressure deficit (DPV) accentuates mainly at noon with increasing evapotranspiration. The magnitude of impact of water deficit depends on climatic variations throughout the day. The climatic variables that affect plant development, are temperature and relative humidity in the form of DPV. Net photosynthesis and growth of cocoa seedlings are physiological variables very sensitive to excess and especially to water deficit.
Keywords: Water stress, drought, photosynthesis, transpiration, water potential.
Introducción
En Colombia existen aproximadamente 100,000 ha cultivadas con cacao (Theobroma cacao L.) con una producción de 39,000 t anuales de almendra seca, que no abastecen la demanda nacional estimada en 47,000 t/año (Espinal, Martínez, Beltrán, y Ortiz, 2005). Los principales departamentos productores de cacao son Santander (46%), Huila (11.5%) y Tolima (5.2 %). Lo anterior indica que el cacao en Colombia se cultiva en diversos ambientes desde muy húmedos hasta secos. En la zona de topografía quebrada de Santander el cultivo está expuesto a situaciones extremas por prevalencia de enfermedades fungosas en ambientes húmedos y per–húmedos, y en el valle seco del rio Magdalena a problemas de déficit hídrico en alguna época del año (García, Romero, y Ortiz, 2007).
El conocimiento generado sobre la respuesta fisiológica de este cultivo a la irrigación, permite deducir que es posible establecer cultivos de cacao en ambientes que contrastan con su sitio de origen, en áreas secas, incluso en ambientes desérticos y/o con menor sombrío donde los rendimientos son aproximadamente de 3 t/ha de cacao seco por año (Carr y Lockwood, 2011; Leite et al, 2012). En los departamentos de Huila y Tolima la mayor limitante para la siembra de cacao es la necesidad de riego suplementario en la época seca, sin embargo, son zonas con disponibilidad de riego ubicadas estratégicamente en ambientes muy competitivos por la infraestructura de servicios y comunicaciones.
En ambientes secos y cálidos se consumen grandes volúmenes de agua para riego debido a la mayor cantidad de energía solar disponible y a la mayor tasa de evapotranspiración por las plantas; mientras que en ambientes más húmedos y cálidos el consumo de agua es menor (Torrán, 2007). La regulación de estomas es el proceso más importante de la planta para evitar la pérdida de agua y está fuertemente afectada por el poder evaporativo de la atmósfera (Doorenbos y Pruitt, 1984). El agua del suelo contenida entre el punto de marchitez permanente (PMP) y la capacidad de campo (CC) se le considera como el agua disponible para la planta (AD), y depende de la textura, la densidad aparente y la estructura y su movimiento ocurre principalmente por fuerzas capilares o matriciales (Allen, Pereira, Raes, y Smith , 2006; Azcón–Bieto y Talon, 2008; Martin, 2010).
El cultivo de cacao es considerado como hidroperiódico. Para Alvim (1981) la precipitación es el factor ambiental que más influye en la producción de este cultivo en condiciones de campo, siendo la periodicidad de las lluvias el principal factor climático que induce la apertura de brotes vegetativos y estimula la floración. Balasimha (1999) considera que la ocurrencia de amplios periodos secos afectan los procesos fisiológicos de esta planta en campo.
En zonas tropicales de Ecuador, Venezuela, Malasia, Ghana y Brasil se han adelantado trabajos a nivel experimental y comercial con cacao bajo riego. Los resultados en el incremento de producción han sido variables entre 40 y 100% (Freire, 1993; Siqueira, Sena, Dias y Souza, 1996). Por otra parte, la producción puede aumentar con la mayor radiación directa en el área de siembra (Dias, 2001; Edward, 1981). En Belice Khan, Patterson y Matlick (1987) evaluaron el comportamiento del cultivo de cacao a la aplicación de riego por goteo en la época seca y encontraron que cuando se restituyó entre 75 y 100% del agua perdida en los primeros 30 cm del suelo, el rendimiento fue 28% más alto que cuando se restituyó 0 y 50% de agua perdida por evapotranspiración.
De Almeida y Valle (2010) encontraron la existencia de genotipos de cacao que no muestran una alta resistencia estomática bajo déficit hídrico y baja humedad relativa, en contraste con otros genotipos que tienen un mecanismo eficiente de regulación estomática y limitan fuertemente la transpiración en condiciones de déficit hídrico. Mediante el estudio del potencial hídrico foliar (Ψf) durante la época seca en un periodo de 5 años, Balasimha y Daniel (1988) identificaron los híbridos 1–21 × NC42/94 y 1–29 × NC23/43 con características de tolerancia a la sequía debido a sus más altos potenciales hídricos foliares y resistencia estomática durante el periodo seco. En el sur de Bahía (Brasil) se compararon ocho clones por tolerancia a la deshidratación rápida, encontrando que los clones SPA5, SIAL70 y TSH516 fueron resistentes a la sequía y su ajuste osmótico fue asociado con la acumulación de iones de potasio y fósforo en la hoja (De Almeida, Brito, Aguilar y Valle, 2001). Araque et al. (2012) en Venezuela compararon las tasas de fotosíntesis, las relaciones hídricas y el crecimiento de plantas de cacao, durante las épocas secas y lluviosas, encontrando que los cultivares con mayor ajuste osmótico coincidieron con las mayores tasas de supervivencia.
Las condiciones vulnerables de la planta de cacao a fenómenos extremos de sequía se presentan en las etapas iniciales de establecimiento y afectan negativamente el crecimiento. Por otra parte, la respuesta de la planta en forma de ajuste osmótico y reducción en el potencial hídrico de la hoja, afectan la asimilación de carbono (De Almeida y Valle, 2010). En Colombia la información de los requerimientos hídricos, los mecanismos de tolerancia, aclimatación o adaptación a condiciones críticas de sequía temporal de clones de cacao es escasa. Por tanto, es necesario iniciar investigaciones que contribuyan al entendimiento de la dinámica del agua, los nutrientes y la luz en el crecimiento y desarrollo del cultivo en nuevos escenarios menos vulnerables a problemas fitosanitarios, contrastantes en luminosidad y disponibilidad hídrica.
Materiales y métodos
El trabajo se realizó en casa de malla del Centro de Investigación Nataima de la Corporación Colombiana de Investigación Agropecuaria (Corpoica) (4° 11’ 31.65” N, 74° 57’ 41.49” O), a 418 m.s.n.m., en la microrregión del valle cálido del Alto Magdalena, departamento del Tolima, municipio de El Espinal, en un bosque seco tropical (IAVH– Colombia, 1998). El promedio anual de temperatura es de 27 °C y la precipitación de 1500 mm, con distribución bimodal. Los suelos son de textura franco arenosa.
Material vegetal y establecimiento del ensayo. Se utilizaron plántulas de T. cacao clones ICS95, ICS60 y CCN51 injertados sobre patrón IMC67 (ICA, 2002). La siembra se hizo en un sustrato de suelo, cascarilla de arroz quemada y gallinaza compostada, en proporción 3:1:1, con fertilidad media a baja, sin problemas de salinidad (0.416 ds/m) y pH (6.43) adecuado para el desarrollo de plantas de cacao, muy bajos contenidos de potasio (0.11 cmol/kg), relación calcio:magnesio moderada (1.14), materia orgánica moderada a baja (2%), fósforo (18.2 mg/kg) y contenido moderado de elementos menores. La textura era franco arenoso (FA), la densidad aparente era baja (1.04 g/cc).
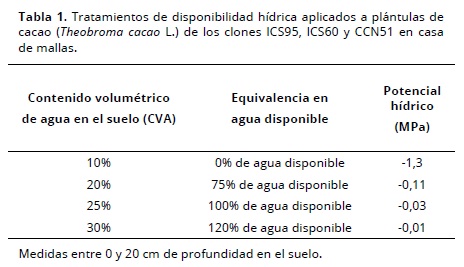
Las mediciones incluyeron: contenidos volumétricos de agua (% CVA) a capacidad de campo (24.75%) y punto de marchitez permanente (10.51%) obtenidos a partir de los datos de humedad gravimétrica mediante ollas de presión (Richards, 1949) y corregidos con la densidad aparente. Las plántulas se establecieron en bolsas de plástico negro calibre 3 de 40 cm de altura y 25 cm de diámetro, con cuatro perforaciones de 0.5 cm de diámetro en la base, se llenaron hasta 3 cm del borde para una altura final de 37 cm de sustrato, con un peso aproximado de 20 kg por cada bolsa. Para mejorar la fertilidad se aplicaron 5 g por bolsa de una mezcla de fertilizante compuesto (15 – 15 – 15) con 20% adicional de KCl. Las plántulas con 1 mes de injertadas se trasplantaron en cada bolsa, se regaron hasta saturación y para alcanzar los valores requeridos del contenido volumétrico de agua en el suelo (CVA) según el tratamiento (Tabla 1) se dejaron en condiciones de libre evaporación y drenaje. En los primeros 20 cm de cada bolsa se midió el CVA con una sonda TDR–100 (Spectrum technologies). Después de cada lectura se agregó la cantidad de agua requerida para mantener el CVA (%) según cada tratamiento.
Variables de clima. Durante 60 días que duró el ensayo fueron registradas la temperatura y la humedad relativa al interior de la casa de malla utilizando Data Logger HOBO U23 Pro v2 (Onset Technologies) y la radiación solar mediante un piranómetro LI–200 acoplado a un Data Logger LI–1000 (LI–COR Bios). A partir de los datos de temperatura y humedad relativa se calculó el DPV. Para lecturas instantáneas se utilizó la fórmula de Rosenberg et al., (1983) citada por (Arellano et al., 2006). Para obtener registros diarios se utilizaron los valores máximos y mínimos de temperatura y humedad relativa según (Allen et al., 2006). El potencial hídrico de la atmósfera (Ψa) fue estimado con los datos de temperatura y humedad relativa, según Azcón–Bieto y Talon (2008).
Potencial hídrico de la hoja Ψf. Al final del experimento, 60 días después del transplante (ddt) se recolectaron tres hojas maduras del tercio superior del tallo principal de una planta por cada unidad experimental (UE), las cuales fueron depositadas en bolsas de aluminio para evitar el aumento de transpiración y se almacenaron temporalmente dentro de una nevera de icopor refrigerada. Antes de que transcurrieran 15 min se midió el potencial hídrico en la bomba de scholander (Soil moisture modelo 3000). Para observar las primeras gotas de agua sobre el corte realizado en el peciolo se utilizó una lupa y se anotó el valor de la presión en bar y MPa (Turner, 1981).
Intercambio de gases. Sesenta ddt se hicieron lecturas de fotosíntesis neta (Pn), transpiración (T), conductancia (gs) y resistencia estomática (rs) en dos plantas por UE. Las mediciones se hicieron con un analizador de gases infrarrojo LI–6200 (LI–COR Bioscenciences) con luz natural, sobre la tercera hoja madura contada desde el ápice hacia la base del injerto con luz ambiente entre las 10:00 y 11:00 a.m. (Rehem, 2006).
Diseño estadístico y análisis de datos. El diseño estadístico utilizado fue parcelas divididas en bloques al azar con cinco repeticiones. La parcela principal correspondió a los tratamientos de agua disponible (10, 20, 25 y 30%) y la subparcela a los clones (ICS95, ICS60 y CCN51) para un total de 60 UE. Los datos de clima fueron organizados en hojas electrónicas en Excel y posteriormente analizados mediante gráficas de tendencia y correlación. Las variables fisiológicas fueron analizadas mediante Anava con el programa SAS versión 9.0 (SAS Institute Inc. Cary, NC, USA. 2002). Para las variables significativas y altamente significativas se hicieron comparaciones de las diferencias entre las medias con base en la prueba de Tukey. Para variables continuas se ajustaron modelos polinómicos mediante el procedimiento GLM de SAS comparando los CVA y la variable respuesta. El modelo de regresión seleccionado se fundamentó en el ajuste estadístico (P < 0.05), la reducción de los cuadrados medios (CME) y el coeficiente de determinación (r2).
Resultados y discusión
Microclima y evapotranspiración. La intercepción de la cubierta disminuyó en un 85% la radiación al interior de la casa de malla, presentando un rango de variación entre 600 y 1000 W/m2 como valores promedio diario. En la Figura 1A se observan los cambios desde las 06:00 h y durante el día de la radiación solar trasmitida hacia el interior a través de la cubierta de la malla, lo que proporcionó la energía suficiente para aumentar la temperatura en el ambiente. A medida que la temperatura ambiente aumentó, disminuyeron la humedad relativa y el potencial hídrico atmosférico al interior de la casa de malla (ΨaCM). El contenido de agua del sustrato (CVA) a 20 cm de profundidad aumentó proporcionalmente a la disminución del Ψa (Figura 1A). Esta variación en el CVA indica la perdida de agua del suelo por traspiración a través de las plántulas de cacao y por evaporación del sustrato expuesto en las bolsas que hacían parte de las UE. La estrecha relación inversa con la humedad relativa indica que la tasa de evapotranspiración en las UE evaluadas, depende del grado de saturación de humedad de la atmósfera interna ΨaCM (Figura 1B).
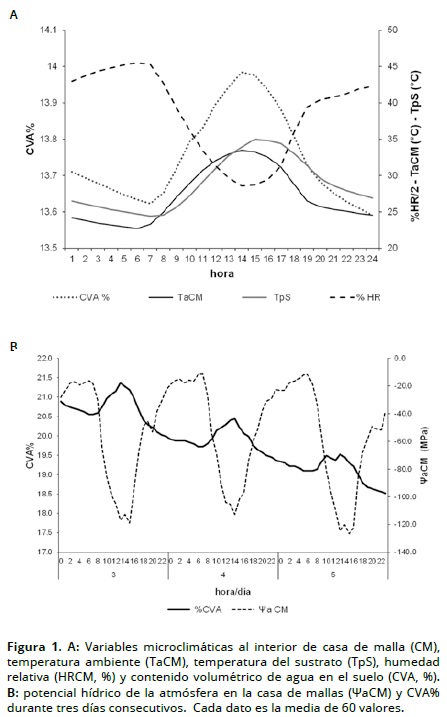
Durante el ensayo, las temperaturas del suelo superiores a 30 °C se presentaron entre las 12:00 y las 19:00 h, momento en el cual disminuyó la evapotranspiración. De acuerdo con lo encontrado por Amorim y Valle (1992) es de particular importancia para trabajos en casa de malla considerar la temperatura del suelo entre 30 y 40 °C, ya que ésta afecta la resistencia al flujo transpiratorio en todos los estados de desarrollo de hojas de plántulas de cacao, lo que altera de manera importante la transpiración.
Déficit de presión de vapor y potencial hídrico de la atmósfera. El DPV presentó un comportamiento similar y altamente correlacionado con la temperatura (r2 = 0.97) (P <0.01). El comportamiento del Ψa fue similar al de la HR, presentando una correlación de 0.99 (P < 0.01). Los valores máximos diurnos de DPV durante el experimento al interior de la casa de malla fueron, aproximadamente, de 2.2 kPa a las 14 horas. En días con temperaturas superiores a 30 °C y humedad relativa por debajo de 60%, el valor aumentó para llegar a un máximo entre 3.5 y 4.24 kPa con las temperaturas máximas alrededor de 40 °C. Estos valores inciden en el cierre normal de estomas ya que las plantas de cacao se saturan con muy baja radiación solar y a DPV entre el aire y la hoja cercanos a 1.8 kPa (De Almeida y Valle, 2007).
Intercambio de gases y Ψf. Se encontraron diferencias (P < 0.01) para el factor CVA. Los clones no se presentaron diferencias (P > 0.05) para las variables evaluadas. La Pn y la T aumentaron proporcionalmente con los tratamientos a medida que aumentó la cantidad de agua disponible en el suelo, siguiendo un modelo cuadrático creciente (Figura 2 A y B). Para Pn se presentó un aumento desde un valor mínimo 3.56 ± 0.71 µmol/m2 por segundo en condiciones cercanas a PMP (10% de CVA) hasta un valor máximo 6.43 ± 0.76 µmol/m2 por segundo con contenidos de humedad a capacidad de campo (25% CVA). A partir de este valor se observó que aumentos en la cantidad de agua disponible (30% CVA) no incrementaron la Pn, la cual se estabiliza o tiende a disminuir. El incremento final en Pn para el tratamiento con 25% de CVA fue de 80% en relación con el tratamiento de 10% de CVA, 34% en relación con 20% de CVA y 16% en relación con 30% de CVA.

Para la transpiración (T) los tratamientos con mayores contenidos de humedad en el suelo (25 y 30% de CVA) presentaron los mayores valores 7.84 ±1.31 y 6.91 ±1.07 mmol/m2 por segundo en relación con los valores más bajos (20 y 10% de CVA) 5.84 ± 1.21 y 4.48 ± 1.32 mmol/m2 por segundo. El tratamiento de 10% de CVA presentó una disminución final en la transpiración del 74% en relación con el tratamiento que no presentó deficiencia hídrica durante todo el experimento (25% de CVA). A media que se intensifica el estrés aumenta la rs (Figura 3 A).

El efecto del microclima en forma de mayores DPV indujo en los tratamientos de 10 y 20% de CVA, un mayor cierre estomático posiblemente vía síntesis de ABA (Davies y Zhang 1991; Moreno, 2009; Dodd, Egea, Watts, y Whalley, 2010), lo que disminuyó la disponibilidad hídrica en los tejidos y el flujo transpiratorio. A medida que las plantas estaban sometidos por mayor tiempo al estrés el efecto fue mayor; situación que indujo a cierres estomáticos tempranos y por lo tanto a la reducción de las tasas de intercambio de agua y dióxido de carbono (Figura 3B).
Las diferentes condiciones de disponibilidad hídrica impuesta tuvieron un efecto directo en la disminución del potencial hídrico de las hojas (Ψf). La tendencia de los valores de Ψf en relación con los CVA% fue de tipo cuadrático (Figura 4A). Los tratamientos tanto de déficit (10% y 20% de CVA) como de exceso hídrico (30% CVA) provocaron un Ψf menor (más negativo) que el obtenido en las plántulas con el tratamiento de 25% de CVA. La disminución del Ψf estuvo correlacionada con disminución en la Pn (Figura 4B). Joly y Hahn (1989) reportaron en cacaos amazónico y trinitario una reducción en las tasas de asimilación y transpiración significativas, cuando el potencial de agua de la hoja alcanzó –0.8 MPa.

En el caso del tratamiento con mayor estrés hídrico (10% de CVA) la perdida de asimilación neta de carbono fue mayor en comparación con los demás tratamientos. De igual manera, se infieren mecanismos de ajuste osmótico, adicionales al efecto del DPV, que disminuyeron el Ψf y a su vez la Pn. Baligar, Bunce, Machado, y Elson (2008) y Balasimha, Daniel, y Bhat (1991) encontraron una relación inversa entre DPV y Pn, de tal manera que cuando la demanda evaporativa fue menor (bajo DPV) la Pn fue mayor y al aumentar DPV, la Pn disminuyó. Paralelamente a la disminución de Pn, T y gs, el ajuste osmótico se considera un mecanismo útil para regular la perdida de agua por las plantas de cacao bajo déficit hídrico (Rada, Jaimez, García Núñez, Azócar, y Ramírez, 2005).
El potencial hídrico foliar, la fotosíntesis neta y el crecimiento de las plántulas de cacao, son variables fisiológicas muy sensibles al exceso y especialmente al déficit de agua. La pérdida de agua en el suelo ocasiona el cierre de estomas de plantas de cacao alterando el intercambio de gases y el déficit de presión de vapor se acentúa principalmente al mediodía cuando aumenta la evapotranspiración. La planta de cacao en niveles de contenido de humedad cercanos a capacidad de campo presenta buen desarrollo, por tanto, se sugieren estudios a nivel bioquímico que permitan conocer respuestas de aclimatación como el ajuste osmótico, ya que esto permitiría hacia futuro la búsqueda de ambientes más secos donde los problemas fitosanitarios tengan menor incidencia.
Agradecimientos
Los autores expresan su agradecimiento al Ministerio de Agricultura y Desarrollo Rural y a la Corporación Colombiana de Investigación (Corpoica) Centro de Investigación Nataima, quienes financiaron y orientaron la realización de este trabajo de investigación.
Referencias
Alvim, P. D. (1981). Recent studies on environmental physiology of cacao. En Proceedings of the 7th International Cacao Research Conference. Lagos, Kenia. p. 85 – 88. [ Links ]
Allen, R. G., Pereira, L. S., Raes, D., y Smith, M. (2006). Evapotranspiración del cultivo: Guías para la determinación de los requerimientos de agua de los cultivos. Food and Agriculture Organization (FAO). 56:323. [ Links ]
Amorim, S. M. C. y Valle, R. R. (1992). Radicular temperature effects in water movement resistence in cacao tree Theobroma cacao L.. Acta Bot. Bras., 6,55 – 64. [ Links ]
Araque, O., Jaimez, R. E., Tezara, W., Coronel, I., Urich, R., y Espinoza, W. (2012). Comparative photosynthesis, water relations, growth and survival rates in juvenile criollo cacao cultivars (Theobroma cacao) during dry and wet seasons. Exp. Agric., 48, 513 – 522. [ Links ]
Arellano García, M. A., Valera Martínez, D. L., Urrestarazu Gavilán, M., García Garza, S., Sánchez Salas, A., y Soria–Ruiz, J. (2006). Estudio del microclima en dos subtipos de invernaderos Almería. Agric. Téc. México, 32,225 – 234. [ Links ]
Azcón–Bieto, J., y Talon, M. (2008). Fundamentos de fisiología vegetal. Segunda Ed. Mc Graw Hill/Interamericana de España. [ Links ]
Balasimha, D. (1999). Stress physiology of cocoa. J. Plant. Crops, 27,1 – 8. [ Links ]
Balasimha, D., y Daniel, E. V. (1988). A screening method for drought tolerance in cocoa. Current Sci. 57:395. [ Links ]
Balasimha, D., Daniel, E. V., y Bhat, P. G. (1991). Influence of environmental factors on photosynthesis in cocoa trees. Agric. Forest Meteorol., 55, 15 – 21. [ Links ]
Baligar, V. C., Bunce, J. A., Machado, R. C,., y Elson, M. K. (2008). Photosynthetic photon flux density, carbon dioxide concentration, and vapor pressure deficit effects on photosynthesis in cacao seedlings. Photosynthetica, 46, 216 – 221. [ Links ]
Carr, M. K., y Lockwood, G. (2011). The water relations and irrigation requirements of cocoa (Theobroma cacao L.): A review. Exp. Agric. 47, 653 – 676. [ Links ]
Davies, W. J., y Zhang, J. (1991). Root signals and the regulation of growth and development of plants in drying soil. Ann. Rev. Plant Physiol. Plant Mol. Biol., 42, 55 – 76. [ Links ]
De Almeida, A. A., y Valle, R. R. (2007). Ecophysiology of the cacao tree. Braz. J. Plant Physiol., 19, 425 – 448. [ Links ]
De Almeida, A. A., y Valle, R. R. (2010). Cacao: ecophysiology of growth and production. En: Ecophysiology of Tropical Tree Crops. F. U. O. V. Fabio Damatta Dept. Of Plant Biology, Vicosa, Brazil, p. 37 – 70. [ Links ]
Dias, L. A. (2001). Melhoramento genético do cacaueiro. 578 p. [ Links ]
Dodd, I. C., Egea, G., Watts, C. W., y Whalley, W. R. (2010). Root water potential integrates discrete soil physical properties to influence ABA signalling during partial rootzone drying. J. Exp. Bot., 61, 3543 – 3551. [ Links ]
Doorenbos, J. y Pruitt, W. (1984). Las necesidades de agua de los cultivos. FAO Riego y Drenaje 24:194. [ Links ]
Edward, N. G. (1981). Potential cocoa photosynthetic productivity. En: 8a International Cocoa Research Conference. Cartagena, Colombia. p. 235 – 244. [ Links ]
Espinal, G. C., Martínez, C. H., Beltrán, L. L., y Ortiz H. L. (2005). La cadena del cacao en Colombia. Una mirada global de su estructura y dinámica. Ministerio de Agricultura y Desarrollo Rural. Observatorio agrocadenas de Colombia. 49 p. [ Links ]
Freire, M. D. (1993). Efeitos da irirgacao complemetar no rendimento do cacaueiro Theobroma cacao, USP Tese de mestrado. ESALQ, Piracicaba, São Paulo – Brasil. [ Links ]
García, L. J., Romero, C. M., y Ortiz, O. L. (2007). Evaluación edafoclimática de las tierras del trópico bajo colombiano para el cultivo de cacao. Corpoica–Produmedios. Bogotá [ Links ].
IAVH–Colombia (1998). El Bosque seco Tropical Bs––T en Colombia. Instituto Alexander von Humboldt, Ministerio del Medio Ambiente. Bogotá. 24 p. [ Links ]
ICA (Instituto Colombiano Agropecuario). (2002). Acuerdo No. 003 Por el cual se define el material genético de cacao. ICA–Consejo Nacional Cacaotero, Bogotá [ Links ].
Joly, R. J.; y Hahn, D. T. (1989). Net CO2 assimilation of cacao seedlings during periods of plant water deficit. Photos. Res., 21, 151 – 159. [ Links ]
Khan, M. N., Patterson, G. R., y Matlick, B. K. (1987). Effect of supplemental water supplied trough drip irrigation on cocoa yield at hummingbird Hershey, Ltd., Belize, C.A. En: Proceedings 10th International Coca Research Conference. p. 185 – 191. [ Links ]
Leite V, J. B., Fonseca V, E., Sodré A, G., Valle, R. R., Nascimento N, M., y Marrocos L, P. C. (2012). Comportamento produtivo de cacau no semiárido do Brasil. Agrotrópica, 24, 85 – 90. [ Links ]
Martin, E. C. (2010). Métodos para medir la humedad del suelo para la programación del riego ¿Cuándo?. University of Arizona. College of agricultural and life sciences. Arizona Cooperative Extension. Arizona. 8 p. [ Links ]
Moreno, L. P. (2009). Respuesta de las plantas al estrés por déficit hídrico. Una revisión. Agronomía Colombiana, 27,179 – 191. [ Links ]
Rada, F., Jaimez, R. E., García Núñez, C., Azócar, A., y Ramírez, M. E. (2005). Relaciones hídricas e intercambio de gases en Theobroma cacao var. Guasare bajo períodos de déficit hídrico. Rev. Fac. Agron., 22,112 – 120. [ Links ]
Rehem, B. C. (2006). Repostas fisiologicas de clones de Theobroma cacao L. ao alagamento do substrato. Tese de Mestrado, Universidade Estadual de Santa Cruz, Ilehus, Bahia. Brasil. [ Links ]
Richards, L. A. (1949). Methods of Mounting Porous Plates Used in Soil Moisture Measurements. Agron. J., 41, 489 – 490. [ Links ]
Siqueira, P. R., Sena G., A., Dias, L. A., y Souza, C. A. (1996). Efeito da irrigação na produtividade do cacaueiro Theobroma cacao L. em Linhares Brasil. En: Proceedings 12th International Coca Research Conference. Salvador, Bahía, Brasil. p. 867 – 877. [ Links ]
Torrán, A. E. (2007). Impacto de las plantaciones de Eucalyptus grandis sobre el contenido de humedad del suelo. Análisis de un caso en el noreste de la provincia de Entrerríos. Tesis de Maestria, Universidad Tecnológica Nacional, Concepción del Uruguay. [ Links ]
Turner, N. C.(1981). Techniques and experimental approaches for the measurement of plant water status. Plant Soil, 58, 339 – 366. [ Links ]
