











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
Black bean (Phaseolus vulgaris L.) is a species produced and widely consumed worldwide. In Brazil, it is economically important and largely cultivated, being the basic food for the population, especially together with rice. Black beans are described for their high antioxidant capacity (Reverri et al., 2015), mainly due to the presence of phenolic compounds (Barreto et al., 2021; Fonseca-Hernández et al., 2021), in addition to being an excellent source of protein (Evangelho et al., 2017), and essential amino acids (Meenu et al., 2023).
The bean plant has a short-cycle C3 metabolism and is very sensitive to weed interference, especially in the early stages of development (Franceschetti et al., 2019; Viecelli et al., 2021). Among the weeds that infest the bean plant, alexandergrass (Urochloa plantaginea) stands out. Alexandergrass belongs to the Poaceae family, has C4 metabolism, rapid growth and development, high seed production with dormancy, and the ability to infest large areas, leading to local dominance and crop shading (Franceschetti et al., 2019; Frandoloso et al., 2019).
In addition, the faster germination of alexandergrass compared to bean results in a highly competitive ability, which can cause serious damage to productivity and crop quality in rice (Galon et al., 2014), maize (Frandoloso et al., 2019; Galon et al., 2019), and beans (Franceschetti et al., 2019; Galon et al., 2022). Weeds are known to compete with the crop for water, nutrients, and light, in addition to releasing allelopathic substances or even harboring insects and diseases that can significantly affect the growth and development of the bean plant, leading to a drastic reduction in productivity and grain quality Galon et al., 2018; Franceschetti et al., 2019; Viecelli et al., 2021; Galon et al., 2022).
The ability of a plant to compete with another plant depends on factors such as row spacing, plant density, seed rate, crop management, soil fertility, water availability, and others in addition to plant species (Mhlanga et al., 2016; Little et al., 2021; Singh et al., 2022). Because weeds occur spontaneously, they exhibit genetic variability, meaning they have greater environmental adaptability than human-selected species, resulting in these species being more competitive than crops (Mahaut et al., 2019; Clements & Jones, 2021). The competitive interactions between two species can be studied by experiments in a replacement series (Rizzardi et al., 2016). This method examines the effect of density and ratio between plants in a weed community, which allows for a better understanding of the competitive process between species. In this way, it can be determined whether the greater competitive ability of the crop or weed is due to the infestation level and overall density or due to greater individual competitive ability (Agostinetto et al., 2013; Rizzardi et al., 2016; Frandoloso et al., 2019).
Species living in community may depend on the same resources available in the environment. Therefore, understanding the mechanisms involved in competition in replacement series allows the development of more efficient and sustainable weed management practices. Therefore, avoiding competition is a strategy that allows stronger growth of one species without harming the neighboring species, or even allows stronger growth of both species. Nevertheless, selecting a cultivar with higher competitive ability may have the greatest crop suppression potential against weeds (Frandoloso et al., 2019).
Therefore, developing strategies that minimize the impacts of weed competitiveness against crops is extremely important. Moreover, identifying more competitive cultivars and knowing the weed densities that have greater impact on bean growth and development will allow for better decision making in weed management. In this context, the objective of this study was to evaluate the intra- and interspecific competition between black bean cultivars (BRS Campeiro and SCS Predileto) in the presence of alexandergrass (U. plantaginea) based on morphological and physiological traits, through the replacement series arrangement.
MATERIAL AND METHODS
Growth conditions, plant material, and experimental design. Five experiments were conducted in a greenhouse at the Federal University of Fronteira Sul (UFFS), Erechim campus, RS, Brazil (27ᵒ43’47”S latitude and 52ᵒ17’37”W longitude, altitude of 670m), during the 2018/19 crop year. The climate of the region is classified as Cfa (humid temperate with hot summer) according to the Köppen-Geiger classification, with rainfall well distributed throughout the year (Peel et al., 2007). Treatments were selected according to the methodology proposed by Radosevich et al. (2007), in which densities are arranged in 100, 75, 50, 25% and 0% of the crop after determining the constant final density in the preliminary experiment.
Beans of the BRS Campeiro (Embrapa Arroz e Feijão, Santo Antônio de Goiás, GO, Brazil) and SCS 204 Predileto (Epagri, Chapecó, SC, Brazil) cultivars and the alexandergrass biotype (Urochloa plantaginea (Link) R.D. Webster) were used. Alexandergrass seeds were obtained from the UFFS experimental area in the 2017/18 crop year and stored in a cold chamber until use. Bean cultivars were selected because they are most commonly used in the region, and alexandergrass because it is one of the most competitive weeds and is commonly found in bean crops.
For both studies, plants were grown in plastic pots (8 dm3) filled with soil collected from an agricultural field and characterized by humic Aluminiferic Red Latosol (Santos et al., 2018). The correction of the pH of the soil and the fertilization of the soil were carried out according to the physicochemical analysis (Table 1) and according to the technical recommendations for bean crop (CQFS - RS/SC 2016). The experimental design used was a randomized block experiment with four replicates.
Table 1 Chemical and physical characteristics of the soil.
| PH | OM | P | K | Al+3 | Ca+2 | Mg+2 | |
| (H2O) | % | ------ mg dm-3 ------ | ----------- cmolc dm-3 ----------- | ||||
| 4.8 | 35 | 4.0 | 117.0 | 0.6 | 4.7 | 1.8 | |
| CEC (t) | CEC (pH = 7.0) | H+Al | BS | V | clay | - | |
| -------------------- cmolc dm-3 -------------------- | % | % | - | ||||
| 7.4 | 16.5 | 9.7 | 6.8 | 41 | 60 | - | |
MO = organic matter; CEC = cation exchange capacity; BS = base saturation; V = current base saturation.
Study 1- Preliminary experiment to determine the plant density at which the final production remains constant. Three preliminary experiments were conducted in monoculture for both bean cultivars and alexandergrass at plant densities of 1, 2, 4, 8, 16, 24, 32, 40, 48, 56 and 64 plants per pot (equivalent to 25, 49, 98, 196, 392, 587, 784, 980, 1176, 1372 and 1568 plants m-2). At 55 days after emergence, the aerial part of the bean and alexandergrass plants were collected. The plant material was dried in a forced-air oven at 65±5°C until it reached a constant matter, to determine shoot dry matter (DM). From the average DM of the species, a constant DM production at a density of 20 plants pot-1 was determined for the bean cultivars and/or the weed, which corresponded to 463 plants m-2 (data not shown).
Study 2 - Evaluation of intra- and interspecific competitive interaction between bean cultivars and alexandergrass weed in replacement series experiments. Two experiments were conducted in replacement series with different ratios of bean cultivars BRS Campeiro and SCS 204 Predileto and the competitor alexandergrass weed, using 20:0, 15:5, 10:10, 5:15 and 0:20 plants per pot (crop versus weed), corresponding to a ratio of 100:0, 75:25, 50:50, 25:75, or 0:100%, maintaining the total density of 20 plants per pot, as determines in the preliminary study.
Seeds were initially germinated in trays, and size-uniformized seedlings were transplanted into 8-dm3 pots as described in the growth conditions topic. Physiological and morphological traits of the bean cultivars and alexandergrass were assessed 55 days after emergence (DAE). Photosynthetic rate (A, µmol m-2 s-1), stomatal conductance (gs, mol m-1 s-1), transpiration (E, mol m-2 s-1), and internal CO2 concentration (Ci, µmol mol-1) were measured between 8 and 11 am on a fully expanded leaf using an infrared gas analyzer (IRGA, LCA PRO, Analytical Development Co. Ltd, Hoddesdon, UK). Carboxylation efficiency (EC, mol CO2 m-2 s-1) and water use efficiency (WUE, mol CO2 mol H2O-1) were calculated from the ratio of A/Ci and A/E, respectively. After physiological analysis, plant height (PH), stem diameter (SD), leaf area (LA), and shoot dry matter (DM) of bean cultivars and alexandergrass plants were evaluated. Leaf area was determined using a portable AF meter (CI-203, CID Bio-Science, Inc., Camas, WA, USA). For DM determination, the aerial parts of the plants were placed in paper bags and dried in a forced-air oven at 65±5°C until they reached a constant matter.
Physiological and morphological data were analyzed using the graphical method of variation or relative productivity. This procedure, also known as the conventional method for replacement series experiments, consists in the construction of diagrams based on relative productivity (RP) or total relative productivity (TRP), as proposed by Bianchi et al., (2006) and Cousens (1991). the RP is a straight line, it means that the ability of the species is equivalent. If the RP results in a concave line, it means that the growth of one or both species is impaired. On the other hand, if the RP shows a convex line, the growth of one or both species is positive. When TRP is equal to 1 (straight line), competition for the same resources occurs; when it is greater than 1 (convex line), competition is avoided; and when TRP is less than 1 (concave line), mutual damage to species growth occurs (Cousens, 1991; Cousens & O´Neill 1993). The relative competitiveness indices (RC), relative clustering coefficient (K), and aggressiveness (AG) of the species were also determined. RC represents the comparative growth of thebean cultivars (x) relative to the alexandergrass competitor (y). K indicates the relative dominance of one species over the other, and AG indicates the most aggressive species. Thus, the indices RC, K, and AG indicate which species is more competitive, and their joint interpretation provides information about species' competitiveness with greater confidence (Cousens, 1991). Bean cultivar (x) is more competitive than alexandergrass (y) when RC > 1, Kx > Ky and AG > 0. On the other hand, alexandergrass (y) is more competitive than bean cultivar (x) when RC < 1, Kx < Ky and AG < 0 (Hoffman and Buhler, 2002). To calculate these indices, the 10:10 plants pot-1 (50:50 ratio) of the species involved in the experiment were used, using the equations RC= RPx/RPy; Kx= RPx/(1-RPx); Ky= RPy/(1-RPy); AG = RPx - RPy, according to Cousens and O´Neill (1993).
Statistical analyzes. Analysis of productivity data or relative variation was performed by calculating the differences for the RP values (DRP) obtained in the 25, 50 and 75% ratios (0.25, 0.50, and 0.75 for RP) relative to the values belonging to the hypothesized line in each ratio (Cousens, 1991; Bianchi et al., 2006). The t-test was used to test the differences in relation to the indices DRP, TRP, RC, K, and AG (Bianchi et al., 2006; Hoffman & Buhler, 2002). To test the differences of DRP and AG, the null hypothesis was made when the means were equal to zero (Ho = 0). For TRP and RC, the null hypothesis was when the means were equal to 1 (Ho = 1); and for K, the null hypothesis was when the means of the differences between Kx and Ky were equal to zero (Ho= (Kx - Ky) = 0). The criterion used to consider the RP and TRP curves as different from the hypothesized lines was that significant differences appeared in at least in two ratios by the t test (2). Similarly, for RC, K, and AG differences in competitiveness were considered if a significant difference was found by the t test in at least two of them (Bianchi et al., 2006). Physiological and morphological data were also subjected to analysis of variance by F-test, and when significant, means were compared with control at 5% probability level by Dunnett's test. Monocultures were used as control treatments. All graphs and coefficients were generated using R statistical software.
RESULTS AND DISCUSSION
Relative Productivities
Photosynthesis-related traits. The relative productivities (RP) for photosynthetic rate (A) showed greater photosynthesis of the alexandergrass with the increase of its ratio in the mixtures with bean cultivars BRS Campeiro and SCS Predileto, with convex concavities, i.e., above the estimated line (Figure 1A, B), contributing to the total relative productivities (TRP) with values greater than 1, in higher ratios than the competitors. Among the tested competitors, the bean cultivar BRS Campeiro showed an increase in photosynthetic rate, convex line, while the cultivar SCS Predileta showed a decrease in A, concave line, from a ratio of 50% (10 plants pot-1). This fact is due to genetic differentiation between bean cultivars (Delfini et al., 2017). However, when TRP is less than 1, it indicates mutual antagonism between species and competing species (Rubin et al., 2014), i.e., both species have a loss of growth and development as they compete for the same environmental resources.

Figure 1 Relative productivity (RP) for photosynthetic rate (a, b), internal CO2 concentration (c, d), and carboxilative efficiency (e, f) of bean (Phaseolus vulgaris) cultivars BRS Campeiro (a, c, e), and SCS Predileto (b, d, f) (●), alexandergrass weed (■), and total community relative productivity (TRP) (▲) as a function of the ratio of associated plants (black bean:alexandergrass).
Considering the TRPs, which represent the mixed density behavior of the crop and weed in their different associations, it was found that there were negative effects for the two bean cultivars at densities lower than 50:50, mainly due to the reduction of A caused by the presence of the alexandergrass (Figure 1A, B). In addition, higher carboxylative efficiency (CE) was observed in the alexandergrass weeds (Figure 1E, F). It has been described that competition between crops and weeds favors the growth of one of the species over the other, which can lead to water and nutrient deficiencies that can affect their metabolism (Manabe et al., 2014).

Figure 2 Relative productivity (RP) for stomatal conductance (a, b), transpiration (c, d), and water use efficiency (e, f) of bean (Phaseolus vulgaris) cultivars BRS Campeiro (a, c, e) and SCS Predileto (b, d, f) (●), alexandergrass (■), and total community relative productivity (TRP) (▲) as a function of the ratio of associated plants (black bean: alexandergrass).
The reduction in photosynthetic rates observed in bean cultivars BRS Campeiro up to a density of 10:10 plants pot-1 or 50:50%, and SCS Predileto in all ratios were accompanied by an increase in internal CO2 concentration (Ci) (Figure 1C, D), even when stomatal conductance (g S) was maintained (Figure 2A, B), indicating that there was a biochemical limitation to photosynthesis. Similar results were observed by Ferreira et al. (2015) when studying the effects of competition between soybean plants and Bidens pilosa and Urochloa brizantha.
The A values were proportional to CE (Figure 1E, F). Both bean cultivars, BRS Campeiro and SCS Predileto, showed concave lines for CE values, regardless of their ratio of competitor plant. On the other hand, the weed CE was reduced when grown with the bean cultivar SCS Predileto up to a ratio of 10:10 plants pot-1, indicating resource competition with alexandergrass (Figure 1F). Therefore, adequate weed management is important to ensure crop growth and productivity (Franceschetti et al., 2019).
Plant dry matter (DM) was reduced by competition in both bean cultivars BRS Campeiro and SCS Predileto and in the competitor alexandergrass, with large losses observed even in TRP (Figure 3C, D). Regardless of bean cultivar, alexandergrass tended to be more affected than the competitor plant. This was evident from the larger differences between observed RP values (solid lines with confidence intervals) and expected values (dotted lines). The high competitive ability of bean cultivars was also observed when the crop was grown with species of the genus Amaranthus, where it is expected that the reduction of the weed DM will allow higher productivity of the crop (Carvalho & Christoffoleti, 2008).
The DM showed reductions of 55% and 70% in TRPs for the BRS Campeiro and SCS Predileto cultivars, respectively (Figure 3C, D). (Manabe et al., 2014). observed that growing field beans together with Amaranthus hybridus resulted in a reduction in crop DM, especially under water restriction at the end of the cycle. However, the bean cultivar IPR Eldorado proved to be more competitive than U. plantaginea due to higher crop DM compared to the weed DM (Cury et al., 2011).
Water use-related traits. Plots with convex lines were generated for leaf area (LA). They indicate higher deviations from the observed values than the estimated ones (Figure 3A, B), in this case there was an advantage for the growth of the crop. The greater relative growth of the bean cultivars compared to the alexandergrass could be related to the density of weed plants, since the competitive ability of weeds is reduced at lower population densities (Agostinetto et al., 2013; Galon et al., 2018).
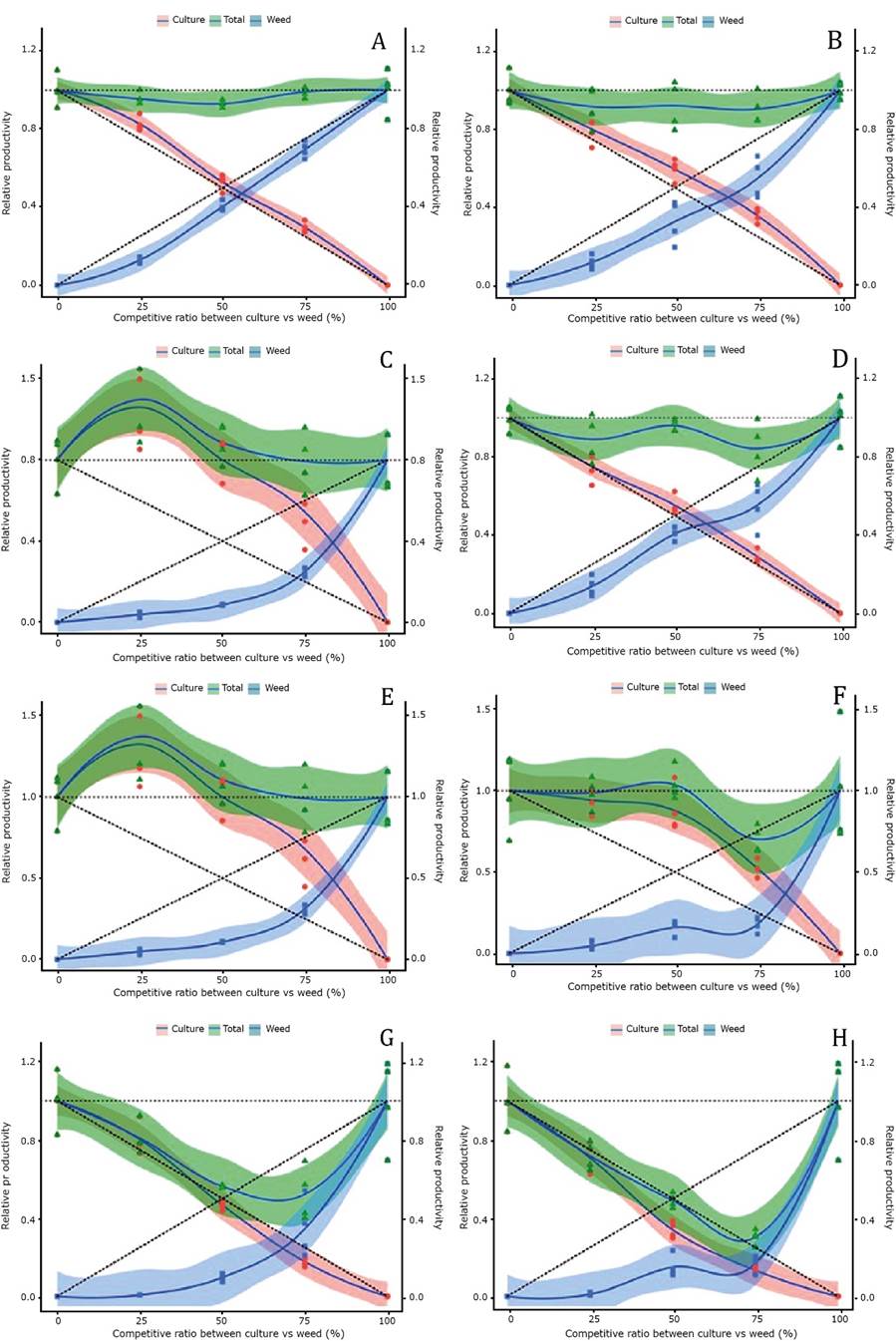
Figure 3 Relative productivity (RP) for plant height (a, b), stem diameter (c, d), leaf area (e, f), and shoot dry matter (g, h) of bean (Phaseolus vulgaris) cultivars, BRS Campeiro (a, c, e) and SCS Predileto (b, d, f) (●), alexandergrass (■), and total community relative productivity (TRP) (▲), as a function of the ratio of associated plants (black bean: alexandergrass).
Production of LA may favor light energy capture and also promote weed shading, as a competitive defense mechanism. The highest relative growth for LA was also observed in bean plants competing with B. pilosa and U. plantaginea (Lage et al., 2017), and in soybean plants when infested with B. pilosa and/or E. heterophylla (Forte et al., 2017).
Domesticated species that compete with different ratios of individuals tend to have higher relative productivity than weeds (Woldeamlak et al., 2001). The crop is more competitive than the competitor because the effect of weeds is not only related to their greater individual competitiveness, but also to the high plant density on the area (Vilà et al., 2004). Agostinetto et al. (2013) reported that weeds generally occur in crops at higher densities, and can therefore be considered more competitive for environmental resources, although this effect is actually a consequence of higher plant density.
Using the PRs and PRTs of E, g S, and WUE, it was demonstrated that the higher stomatal conductance of bean crops (Figure 2A, B) allows for higher E (Figure 2C, D) and consequently lower WUE (Figure 2E, F). Water is an important limiting resource for crops. Thus, reduced water availability can negatively affect crop biomass when crops are in competition (Wanic et al., 2013). However, variables associated with water use indicate that water competition was not a predominant factor in bean cultivar growth and productivity.
Growth traits. The RP in relation to plant height (PH) show that the bean cultivars have convex lines in all ratios of plants, with BRS Campeiro having the most distant values and contributing to TRP values close to 1 (Figure 3A, B). The increase in PH indicates that plants were etiolated due to competition for light. The RP and TRP (~1) for stem diameter (SD) of all participating species (bean cultivars and alexandergrass) were slightly affected by competition, especially for bean cultivars (Figure 3C, D).
Although discrete, it was observed that both bean cultivars (BRS Campeiro and SCS Predileto) had convex lines with values above the expected value and the alexandergrass weed had concave lines below the expected value, showing that the bean cultivars excelled compared to the weed. The faster development of plant growth compared to the weed was also confirmed by Lage et al., (2017) when they analyzed the disturbance of the arrangement of bean plants (cultivar IPR Eldorado) in competition with B. pilosa and U. plantaginea.
Light competition can stimulate greater shoot development than the root system, as a strategy mechanism to increase and maximize light energy uptake. Light restriction is one of the most important limiting factors for initial plant growth in a community and can directly affect plant production (Page et al., 2010). In the present work, alexandergrass showed greater competitiveness than bean cultivars for photosynthetic activity, with lines of convex lines and concave culture, showing that the weed was more efficient in utilizing solar radiation.
It is important to highlight that the plants that establish first have a competitive advantage that can be facilitated by intrinsic characteristics of each cultivar or hybrid in terms of competitiveness, such as germination speed, rapid development, which reduce the available resources and cause damage to other species that can be the weed or crop (Agostinetto et al., 2013; Forte et al., 2017).
Competitiveness indices
Photosynthesis-related traits. Photosynthetic activity (A), carboxylation efficiency (EC), and internal CO2 concentration (Ci) did not differ between bean cultivars and alexandergrass with control treatments without competition in at least two ratios of plants in the crop versus competitor associations (Table 2).
Table 2 Physiological traits of Phaseolus vulgaris, cultivars BRS Campeiro and SCS Predileto, grown together with the weed alexandergrass in a replacement series arrangement, 55 days after plant emergence. UFFS, Erechim/RS.
| Plant ratio (%) Bean:weed | Gas exchange traits | ||||||
| A (µmol m-2 s-1) | gs (mol m-1 s-1) | Ci (µmol mol-1) | E (mol m-2 s-1) | CE | WUE | ||
| BRS Campeiro | |||||||
| 100:0 (C) | 4.69 | 0.23 | 324.25 | 3.59 | 0.04 | 2.24 | |
| 75:25 | 2.81 | 0.32 | 333.75 | 4.34 | 0.02 | 1.88 | |
| 50:50 | 4.72 | 0.18 | 309.25 | 3.21 | 0.02 | 1.88 | |
| 25:75 | 7.16 | 0.35 | 358.00 | 5.27 | 0.03 | 1.59 | |
| C.V (%) | 58.20 | 64.50 | 14.60 | 46.50 | 109.50 | 68.30 | |
| Alexandergrass | |||||||
| 0:100 (C) | 2.21 | 0.07 | 328.50 | 0.94 | 0.01 | 2.70 | |
| 25:75 | 2.52 | 0.04 | 302.25 | 0.72 | 0.01 | 3.49 | |
| 50:50 | 4.03 | 0.05 | 266.00 | 0.76 | 0.02* | 5.17* | |
| 75:25 | 3.43 | 0.05 | 278.00 | 0.71 | 0.01 | 4.89 | |
| C.V (%) | 43.50 | 40.50 | 25.60 | 46.9 | 53.70 | 37.90 | |
| SCS Predileto | |||||||
| 100:0 (C) | 8.94 | 0.36 | 333.00 | 4.72 | 0.03 | 1.93 | |
| 75:25 | 6.29 | 0.36 | 346.25 | 4.92 | 0.02 | 1.19 | |
| 50:50 | 9.02 | 0.42 | 334.50 | 5.09 | 0.03 | 2.06 | |
| 25:75 | 3.19 | 0.44 | 373.33 | 5.92 | 0.01 | 0.53 | |
| C.V (%) | 84.90 | 36.60 | 9.50 | 23.20 | 102.40 | 89.80 | |
| Alexandergrass | |||||||
| 0:100 (C) | 2.16 | 0.07 | 328.50 | 0.94 | 0.01 | 2.59 | |
| 25:75 | 3.44 | 0.04 | 275.50 | 0.75 | 0.01 | 4.66 | |
| 50:50 | 2.28 | 0.04 | 317.50 | 0.79 | 0.01 | 3.14 | |
| 75:25 | 2.28 | 0.05 | 356.25 | 0.76 | 0.00 | 2.79 | |
| C.V (%) | 65.60 | 46.5 | 29.10 | 50.10 | 120.80 | 61.10 | |
Asterisk (*) indicates significant difference compared to control (C) by Dunnett´s test (p < 0.05).
When the competitiveness indices (RC, K and AG) were evaluated, a significant effect was found only for CE of the BRS Campeiro cultivar in competition with alexandergrass (Table 4), where the weed was more competitive than the crop. The results show that although the species did not benefit from the competition, they were not affected either, proving that no species had an advantage over another. The LA of the two bean cultivars were higher than expected (Figure 3A, B) and lower for alexandergrass weed. However, the higher LA did not contribute to an increase in A in the present study.
The DM of the bean cultivars BRS Campeiro and SCS Predileto and also of the alexandergrass weed competitor differed significantly from the respective control (competition-free) treatment for all different associations between species (Table 3). The greater the ratio of competitors in the interaction with bean cultivars, the higher the observed damage to the crop. The effect of interspecific competition was more harmful to the accumulation of DM than intraspecific competition, i.e., the higher the plant density at association, the higher the damage. This fact was also confirmed by Galon et al. (2017), who studied the response of bean cultivars in competition with B. pilosa.
Table 3 Morphological traits of Phaseolus vulgaris, cultivars BRS Campeiro and SCS Predileto, grown with the weed alexandergrass in a replacement series arrangement, 55 days after plant emergence. UFFS, Erechim/RS.
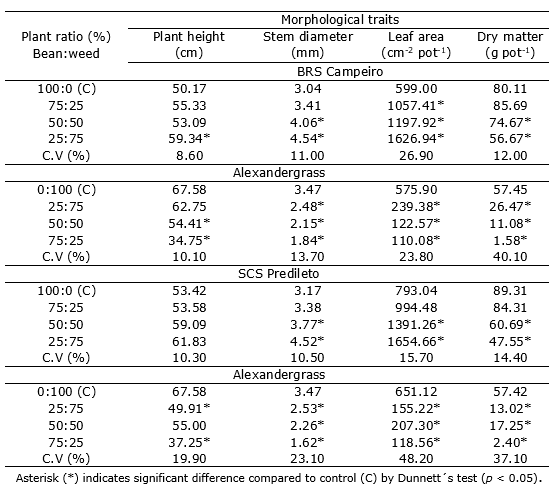
The relative competitiveness, the clustering coefficient (Kbean and Kalexandergrass) and the aggressiveness coefficient had significant effects for the bean cultivars in grown with alexandergrass for the DM, with the crop RC > 1, Kbean > Kalexandergrass and AG > 0 in all situations (Table 4). Thus, it was found that the bean cultivars have greater competitiveness than the alexandergrass at DM. It is likely that the higher LA observed in the bean cultivars (Figure 3A, B) favored a competitive advantage for the variable DM (Table 3). Other work also reported differences in competitive ability among bean cultivars infested with B. pilosa and U. plantaginea (Araújo et al., 2018) and/or with E. heterophylla (Galon et al., 2018).
Table 4 Competitive indices of Phaseolus vulgaris, cultivars BRS Campeiro and SCS Predileto, grown with the weed alexandergrass in an equal plant ratio (10:10 plants-1 pot-1, bean:weed) in a replacement series arrangement, 55 days after plant emergence. UFFS, Erechim/RS.
| Variable | Relative competitiveness1 | Kx2 (bean) | Ky (alexandergrass) | Aggressiveness3 | |
| Photosynthetic rate (A) | |||||
| BRS Campeiro x weed | 0.714 ± 0.342 | 1.513 ± 0.630 | 10.919 ± 21.124 | -0.411 ± 0.262 | |
| SCS Predileto x weed | 1.342 ± 0.568 | 1.862 ± 0.900 | 2.850 ± 1.897 | -0.025 ± 0.231 | |
| Stomatal conductance (g S) | |||||
| BRS Campeiro x weed | 1.228 ± 0.517 | 1.432 ± 0.995 | 0.535 ± 0.067 | 0.056 ± 0.177 | |
| SCS Predileto x weed | 1.922 ± 0.215* | 1.497 ± 0.243* | 0.454 ± 0.042 | 0.276 ± 0.056* | |
| Internal CO2 concentration (Ci) | |||||
| BRS Campeiro x weed | 1.195 ± 0,084 | 0.914 ± 0.039 | 0.693 ± 0.085 | 0.072 ± 0.029 | |
| SCS Predileto x weed | 1.075 ± 0.130 | 1.034 ± 0.134 | 0.988 ± 0.184 | 0.019 ± 0.061 | |
| Transpiration (E) | |||||
| BRS Campeiro x weed | 1.176 ± 0.411 | 1.121 ± 0.452 | 0.698 ± 0.102 | 0.043 ± 0.142 | |
| SCS Predileto x weed | 1.305 ± 0.187 | 1.369 ± 00.424 | 0.741 ± 0.098 | 0.118 ± 0.084 | |
| Carboxilative efficiency (CE) | |||||
| BRS Campeiro x weed | 0.229 ± 0.092* | 0.445 ± 0.273 | -2.500 ± 1.500 | -0.917 ± 0.194* | |
| SCS Predileto x weed | 0.180±0.083 | 3.310 ± 2.303 | -0.500 ± 1.258 | 0.000 ± 0.327 | |
| Water use efficiency (WUE) | |||||
| BRS Campeiro x weed | 0.446 ± 0.082* | 0.901 ± 0.391 | 0.347 ± 6.815 | -0.541 ± 0.123* | |
| SCS Predileto x weed | 1.475 ± 0.871 | 4.993 ± 3.055 | -0.755 ± 2.043 | -0.070 ± 0.294 | |
| Plant height (PH) | |||||
| BRS Campeiro x weed | 1.322 ± 0.086* | 1.135 ± 0.086* | 0.676 ± 0.036 | 0.126 ± 0.032* | |
| SCS Predileto x weed | 1.371 ± 0.111* | 1.259 ± 0.134* | 0.689 ± 0.043 | 0.146 ± 0.037* | |
| Stem diameter (SD) | |||||
| BRS Campeiro x weed | 2.184 ± 0.227* | 2.190 ± 0.450* | 0.451 ± 0.026 | 0.359 ± 0.054* | |
| SCS Predileto x weed | 2.030 ± 0.443 | 1.502 ± 0.158* | 0.512 ± 0.119 | 0.269 ± 0.062* | |
| Leaf area (LA) | |||||
| BRS Campeiro x weed | 9.385 ± 0.460* | 1.259 ± 8.252 | 0.119 ± 0.002 | 0.894 ± 0.057* | |
| SCS Predileto x weed | 6.159 ± 1.641* | 0.043 ± 4.554 | 0.192 ± 0.029 | 0.718 ± 0.091* | |
| Shoot dry matter (DM) | |||||
| BRS Campeiro x weed | 5.031 ± 0.641* | 0.875 ± 0.037* | 0.107 ± 0.120 | 0.370 ± 0.020* | |
| SCS Predileto x weed | 2.527 ± 0.485* | 0.519 ± 0.047* | 0.181 ± 0.043 | 0.190 ± 0.047* | |
Asterisk (*) indicates significant difference compared to control treatment (monoculture) by Dunnett´s test (p < 0.05). 1significant when RC differed from 1 by t test; 2difference between Kx and Ky at the same level of competition by Welch´s criterion t test; 3significant when AG differed from 0 by the t test.
The difference in competitive ability of common bean cultivars is related to sowing density and cultivar choice (Lage et al., 2017; Galon et al., 2018). However, competitive ability decreases when adequate weed management occurs, i.e., the crop is better under this condition (Andrew & Storkey, 2017). Therefore, it is important for organic farming systems to find out which cultivar is more competitive when infested with a particular weed species (Andrew & Storkey, 2017). This allows for the use of a lower amount of herbicides for weed control and thus greater economic and environmental benefits or even the production of safer food.
Water use-related traits. The LA differed for the BRS Campeiro and SCS Predileto cultivars in competition with the alexandergrass from the respective monoculture controls (Table 3). The greater the ratio of the competitor, the greater was the LA of the bean cultivars, and the opposite was observed for the weed. In this case, intraspecific competition was more harmful to the crop and interspecific competition was more harmful to the weed.
The LA-based RC was consistent, demonstrating the superiority of the crop to alexandergrass infestation regardless of bean cultivar. A similar result was observed for Kx and AG based on LA. Manabe et al. (2014) reported that weeds have a greater capacity for root development and consequently uptake more water from the soil, which is important in situations of water scarcity but may affect shoot growth under non-stressful conditions.
Growth traits. The PH and SD of bean cultivars BRS Campeiro and SCS Predileto in competition with alexandergrass differed from the isolated controls (without competition) (Table 3). The BRS Campeiro cultivar had the highest PH with the lowest bean plant density and the highest competitor plant density (Table 3), with significant effects in at least two situations. The weed competing with the cultivar BRS Campeiro showed an increase of PH when the association was higher weed density and lower bean crop density.
The cultivar SCS Predileto showed no significant effect on PH in at least two situations when the crop and weed competed (Table 3). Alexandergrass showed lower PH at lower density and the SCS Predileto cultivar at higher density. Passini et al. (2003) observed that bean plants emerged first and grew faster than Brachiaria plantaginea, which also had taller plants and higher LA, contributing to the better competitive performance of bean plants.
The SD of bean cultivars increased with decreasing bean plant density and increasing alexandergrass density. Bean cultivar plant density decreased with decreasing crop plant density and increasing alexandergrass density in the association. A similar response was observed for weed, where increasing bean plant density and decreasing weed density resulted in lower SD for alexandergrass plants. For both PH and SD, the interspecific competition was more harmful to bean cultivars than intraspecific competition. Similar results were previously observed for bean cultivars in competition with B. pilosa (Galon et al., 2018). Regarding the competitiveness indices between species, RC > 1, Kbean > Kalexandergrass, and AG > 0 of bean cultivars BRS Campeiro and SCS Predileto were detected in competition with alexandergrass (Table 4). Thus, considering the three competitiveness indices (RC, Kx and AG), the bean plant showed greater relative growth than the alexandergrass.
Studies report that the crop may have higher competitive potential compared to weeds when intraspecific competition (between individuals of the same species) is greater than interspecific competition (between individuals of different species) (Vilà et al., 2004). It is also noteworthy that usually the same ecological niche is used by plants of the same species, so that plants compete for the same resources in space and time (Bianchi et al., 2006; Forte et al., 2017; Baldessarini et al., 2020). Carvalho & Christoffoleti (2008) also found that the bean crop was more competitive with Amaranthus species, with benefits to the crop and losses to the weed.
CONCLUSIONS
Bean crops and alexandergrass compete with each other, resulting in mutual damage to the species involved in the community. Alexandergrass negatively affects the morphophysiological traits of bean cultivars BRS Campeiro and SCS Predileto, as it has a higher competitive ability than the crop plants. On the other hand, bean plants showed higher relative growth than alexandergrass, with both species responding similarly. Intraspecific competition was generally more harmful to both species than interspecific competition.

