











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
El objetivo de los inventarios florísticos cuantitativos de la vegetación obtener información sobre la riqueza, diversidad y abundancia de las especies de un determinado sitio, lo cual permite comparar los resultados obtenidos en otras zonas usando la misma metodología, en aras de replicarla en otros ecosistemas naturales (Tirado, 2016).
Manzanilla et al. (2020) afirman que determinar la estructura florística vertical y horizontal y la densidad de individuos dentro de un área boscosa permite conocer la naturaleza de la masa forestal con base en su diversidad biológica y composición florística, así como la abundancia de las especies que forman comunidades vegetales en la distribución espacial y en los estratos altitudinales.
Lamentablemente, poco se conoce sobre la composición y estructura de los bosques húmedos tropicales; se hace indispensable realizar estudios cuantitativos que permitan conocer sus condiciones biológicas, en aras de establecer criterios de manejo y conservación a través del monitoreo de estos bosques naturales (Fabián, 2016).
Dicho lo anterior, la composición y diversidad florística arbórea a escala local de un bosque natural del Chocó biogeográfico (una de las regiones más lluviosas del mundo, que cuenta con alta biodiversidad y endemismo) y su relación con las características de los sitios y la diversidad o la distribución de las especies arbóreas de los bosques están determinadas parcialmente por variables ambientales como la topografía, la fertilidad y el tipo del suelo, entre otros (Mosquera & Hrtado, 2014).
En estudios realizados en el Chocó biogeográfico (Langendoen & Gentry, 1991; Galeano, 2002; Torres et al., 2016; Mosquera et al., 2007; Quinto & Mosquera (2014), se ha encontrado que los bosques tienen características particulares en términos de la estructura y composición florística de sus ecosistemas naturales. En particular, se presenta una alta densidad de árboles por ecosistemas y un número comparativamente elevado de especies. En este sentido, se ha registrado un promedio de entre 150 y 250 especies árboles por hectárea, lo cual sitúa a esta región en el segundo lugar en cuanto a riqueza florística y diversidad en Colombia (después de la Amazonía) (Casas et al., 2018).
Por lo tanto, el establecimiento y monitoreo de parcelas permanentes ha permitido entender la dinámica y biodiversidad de los bosques tropicales, así como su estrecha relación con el clima y el suelo (Álvarez et al., 2021).
Es probable que el área cubierta por bosques cambie a medida que varía el clima, como también es posible que haya cambios en los tipos de bosques debido a las modificaciones en las temperaturas y los regímenes de precipitación, donde se espera que la superficie forestal se expanda en la zona templada y se contraiga en las regiones boreales, tropicales y montañosas. Es por ello que el cambio climático puede tener impactos profundos en la salud y vitalidad de los bosques del mundo. Dichos impactos se determinan considerando factores como edad, estructura, composición, función, vigor, niveles inusuales de insectos o enfermedades y resistencia a las perturbaciones (FAO, 2012).
Asimismo, el papel de la conservación, el manejo sostenible de los bosques y la mejora de las reservas de carbono forestal (conocido como REDD +) son vitales en el marco de los esfuerzos mundiales para combatir el cambio climático. Producto del Acuerdo de París sobre el cambio climático, celebrado en diciembre de 2015 en la CMNUCC, los países acordaron conservar y mejorar los sumideros y reservorios de carbono (FAO, 2012).
En este contexto, los bosques naturales juegan un rol importante en la regulación climática, pues son capaces de fijar CO2, el principal gas de efecto invernadero, mediante la fotosíntesis, regulando el intercambio de carbono entre la atmósfera y la biomasa. Lamentablemente, este tipo de servicio ambiental no ha sido suficientemente evaluado en términos de retención de carbono y su valor económico para la sociedad (Valera & Carvajal, 2014).
El carbono almacenado en los bosques colombianos estuvo por encima de los 7800 millones de toneladas para el año 2007. Por lo tanto, las mayores reservas se encuentran en los bosques tropicales muy húmedos (135.29 t.ha-1), seguido de los bosques tropicales húmedos (131.87 t.ha-1). A nivel regional, el pacífico ocupa el segundo lugar, después de la Amazonía, con los promedios más altos de carbono (131 t.ha-1), aun cuando su extensión total es de las menores (7 millones de hectáreas) (Ríos & Gómez, 2018).
Esta tendencia se repite en el área de estudio, el departamento del Chocó, cuyo contenido promedio de carbono para bosques naturales se estima por encima de las 135 t.ha-1, ocupando el tercer lugar a nivel departamental (Rios & Gómez, 2018).
La importancia de los bosques como fijadores de carbono (C) puede constatarse a partir de la estimación de la biomasa forestal. Por lo general, el C almacenado en la vegetación se obtiene del producto entre la biomasa vegetal y un factor de conversión a C (fracción de C). Conocer este coeficiente es de sumo interés para lograr estimaciones precisas (Sione et al., 2019).
En este contexto, determinar la fracción de carbono (Fc) de un bosque representa la base para conocer los stock de C presentes en sus componentes, que tienen gran capacidad para captar y almacenar C y constituyen una de las maneras más efectivas y económicas para remover el C de la atmósfera a través del manejo sostenible de los bosques. En estos ecosistemas hay árboles y otras plantas que, a medida que crecen, remueven importantes cantidades de CO2 de la atmósfera y almacenan el C en la biomasa de hojas, flores, frutos, ramas, tallos y raíces (Sánchez, 2016).
Por consiguiente, el panel intergubernamental de cambio climático IPCC (2006) ha proporcionado diferentes valores de la fracción de C para bosques. Estos, según el tipo de especies arbóreas y basados en análisis químicos de muestras, son de 0.47 para especies tropicales y subtropicales y de 0.48 y 0.51 para especies de hoja ancha y coníferas en bosques templados respectivamente.
De igual manera, Sione et al. (2019) determinaron la Fc de Algarrobo Prosopis affinis en tres componentes aéreos de la biomasa como fustes (F, 0.48), ramas grandes (RG, 0.48) y ramas pequeñas, hojas, flores y frutos (RPHFF, 0.45) en un bosque nativo de Argentina, con una fracción de C promedio entre componentes de 0.47.
Dryw y Hara (2018) determinaron la fracción de carbono por combustión en el reservorio del tronco de nueve especies coníferas ecológicamente importantes en California, donde encontraron valores de Fc para Abeto Douglas costero (Pseudotsuga menziesii) de 0.58, de 0.54 para secuoya gigante (Torr florin), de 0.56 para cedro del incienso (Calocedrus decurrens), de 0.51 para pino Jeffrey (Pinus jeffreyi Balf.), de 0.52 para pino ponderosa (P. ponderosa), de 0.52 para abeto rojo (Abies magnifica), de 0.57 para pino azucarero (P. lambertiana), de 0.52 para abeto blanco (A. concolor) y de 0.53 para Franco var (menziesii), valores significativamente importantes para especies de Norteamérica.
En el mismo contexto, Duque (2020) generó directrices para la selección de ecuaciones, parámetros y datos para calcular las remociones de GEI de actividades forestales, en aras de realizar estimaciones pertinentes, coherentes, exactas y transparentes. En este contexto, se calcularon la densidad básica, el factor de expansión de biomasa y la fracción de carbono (Fc) para 12 especies forestales en Colombia. Se observaron valores de Fc para individuos de Acacia mangium Willd de 0.54, para Cupressus lusitanica Mill de 0.54, para Eucalyptus camaldulensis Dehnhardt de 0.47, para Eucalyptus globulus Labill. de 0.45, para Eucalyptus grandis W. Hill ex Maiden de 0.46, para Eucalyptus urophyla ST Blake de 0.46, para Eucalyptus urograndis de 0.46, para Gmelina arborea Roxb. ex Sm de 0.42, para Hevea brasiliensis Muell. Arg. de 0.43, para Pachira quinata (Jacq.) W.S. Alverson de 0.39, para Pinus patula Schltdl. & Cham de 0.55 y para Tectona grandis L. f. de 0.54.
Teniendo en cuenta la importancia del objeto de estudio, los autores no tienen conocimiento de trabajos que aporten a la identificación de la fracción de carbono en los bosques del departamento del Chocó. Es por ello que la presente investigación determinó la Fc en los ecosistemas naturales de los municipios de Quibdó, Unión Panamericana y Unguía. Los resultados de esta investigación podrían contribuir a precisar el potencial de estas zonas en cuanto a fijación y almacenamiento de carbono. Atendiendo esta consideración, se pretende dar a conocer cuál es la fracción de carbono de las especies de mayor peso ecológico en los bosques tropicales de Pacurita, Salero y Puerto en Chocó, Colombia.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se desarrolló en los corregimientos de Pacurita, Salero y Puerto de los municipios de Quibdó, Unión Panamericana y Unguía respectivamente, en departamento del Chocó, Colombia.
El municipio de Quibdó tiene una extensión de 180 Km2, con una altura geográfica de 53 m, presentando temperaturas superiores a los 28 °C y una humedad relativa de 90 %. Este municipio alcanza una precipitación anual que oscila entre 6000 y 8000 mm. El corregimiento de Pacurita, por su parte, corresponde a las zonas de vida bosque pluvial tropical (bp-T) y bosque muy húmedo premontano (bmh-PM). Bajo esas condiciones climáticas se establecieron tres parcelas permanentes de monitoreo en las siguientes coordenadas geográficas: PPM 1- NE 5°41.960N - 76°35,800W; PPM 2- NE: 5°40.253N - 76°34.951W; PPM 3- NE: 5°42.280N - 76°26.241W (Alcaldía de Quibdó-Chocó, 2005).
El municipio de Unión Panamericana tiene una extensión de 3986.13 ha, con una altura geográfica de 150 m de altitud. Se caracteriza por tener una humedad relativa por encima del 85 % y concurrentes en temperaturas 20.07-34. 9 °C, alcanzando precipitaciones que normalmente superan los 5000 mm. La mayor parte del territorio está situado en la región de las calmas ecuatoriales. Según el sistema de Holdrige, corresponde a las zonas de vida bosque pluvial tropical (bp-T) y bosques muy húmedo tropical (bmh-T). En el corregimiento de Salero se establecieron dos parcelas permanentes de monitoreo, georreferenciadas con las siguientes coordenadas: PPM 1- NE: 5°18.925N - 76°36.184W; PPM 2- NE: 5°19.700N - 76°37.181W (Alcaldía de Unión Panamericana-Chocó, 2015).
El municipio de Unguía, ubicado en la subregión del Darién Chocoano, con una extensión de 1307 km2, se caracteriza por su gran riqueza hídrica y biodiversidad, cuyo principal eje es el río Atrato, ligado a un clima húmedo y lluvioso, con una precipitación media anual de 2089 mm y una temperatura promedio de 28 °C. Tiene áreas circundantes en estuarios con una altura geográfica de 15 m. La vegetación donde se llevó a cabo el estudio pertenecía a un ecosistema homogéneo de P. copaifera. Se estableció una parcela permanente de monitoreo georreferenciada de la siguiente manera: PPM 1- NE: 8°01.316N - 77°04.330W (Alcaldía de Unguía-Chocó, 2005).
Establecimiento de unidades de muestreo
Las áreas se identificaron revisando la cartografía disponible en el Centro de Sistemas de Información Geográfica de la Universidad Tecnológica del Chocó. Los sitios de muestreo se seleccionaron al azar a través del programa ArcView. Posteriormente, se buscaron dichas coordenadas en campo mediante recorridos y sondeos, y se establecieron seis PPM de 100 x 100 m (1 ha), distribuidas de la siguiente manera: en Pacurita, tres PPM inmersas en el bosque primario; en Salero, dos en ecosistemas de bosque secundario poco intervenido; y en Unguía, una en el ecosistema catival. Cada PPM de 1 ha se dividió en cinco bloques de 100 x 20 m, que a su vez se subdividieron en sub-bloques de 20 x 20 m, para un total de 25 sub-bloques de 400 m2 cada uno. Estos bloques fueron identificados con combinaciones consecutivas de números o letras: de 1 a 5 en orientación W-E, y con letras de la A hasta la E en sentido S-N, iniciando por el sub-bloque 1A en zig-zag hasta llegar al bloque 1E, para luego pasar a los bloques 2, 3 y 4 en zig-zag hasta llegar al bloque 5E. El muestreo inició en la esquina SW (sub-bloque 1A0), según las recomendaciones del Ministerio de Medio Ambiente (2002).
Caracterización de la composición florística
Se registraron los nombres vulgares y científicos, y se midió el diámetro a la altura del pecho (DAP ≥ 10 cm), registrando la altura total (ht) de todos los individuos de las PPM.
En cada PPM se estimaron la abundancia, la frecuencia, la dominancia (basado en el área basal) y el índice de valor de importancia empleando las siguientes ecuaciones:

Donde: Ab = área basal (m2); d² = diámetro del tronco medido a 1,30 m del suelo al cuadrado (cm).

Donde: V = volumen (m3); V3 = área basal (m2); altura comercial; factor forma (0.7854).



Donde: Ar = abundancia relativa (%), que es la relación porcentual del número de árboles de una especie frente al número total de árboles muestreados; Nsp = número de árboles por especie; Nt: número total de árboles del área muestreada.


Donde: Fr = frecuencia relativa (%), que es la relación porcentual de la frecuencia absoluta de una especie entre la sumatoria total de las frecuencias absolutas de todas las especies; Fasp = frecuencia absoluta de una especie (%); ∑Fa = sumatoria de frecuencias absolutas de las especies (%).


Donde: Dr% = dominancia relativa (%), que es la relación porcentual entre el área basal de una especie y la sumatoria total de las áreas basales de las especies; Ab = área basal de cada especie (m2): ∑Ab = área basal total en el área de muestreo (m2).

Donde: IVI = índice de valor de importancia; Fr = frecuencia relativa (%); Ar = abundancia relativa (%); Dr = dominancia relativa (%).
Con base en el IVI, se seleccionaron las especies de mayor peso ecológico de los tres bosques.
Determinación de la fracción de carbono
Se seleccionaron 39 especies distribuidas de la siguiente manera: 23 en Pacurita, 15 en Salero y 1 en Unguía.
Recolección de muestras
Se obtuvo una muestra de 500 gr de las ramas de un individuo de cada especie, las cuales fueron forradas con papel periódico para enviarlas al laboratorio.
Análisis de laboratorio
Se obtuvo la Fc presente en la biomasa aérea utilizando el método de calorimetría sugerido por la tesis de Segura (1997), que consistió en tres fases:
La fase 1 consistió en preparar todas las muestras para la obtención del porcentaje de materia seca (%M.S.), para lo cual se trituraron las muestras mediante un molino con una malla de 0.1 mm. Estas se colocaron en placas de Petri, donde se pesó 1 g de la muestra homogenizada. Las muestras se situaron en un horno al vacío hasta alcanzar 70 ºC, a una presión de 100 mm Hg, por un lapso de 24 horas (PSH). Pasado ese tiempo, se procedió calcular el porcentaje de materia seca a través de la siguiente fórmula:

Donde: %MS= porcentaje de materia seca; PSH= peso seco al horno (g); PS= peso seco (g).
Se estandarizó la bomba calorimétrica de metal pesado W = 2404,96. Simultáneamente se insertó una muestra de 1 g en forma de pastilla (PM), y se colocaron 10 cm de hilo fino de platino, admitiendo oxígeno puro a presión (20 Atm). La muestra se sumergió con la bomba en dos (2) litros exactos de agua destilada, y se inició el proceso hasta conocer el poder calorífico.
El objetivo principal de la fase 2 fue conocer las conversiones y el cálculo de la fracción de carbono mediante la energía liberada por la muestra, donde se tomó como referencia la temperatura inicial (antes de la combustión). Aquí, cuando la temperatura llega a ser constante se quema la muestra por el calentamiento momentáneo del hilo fino de platino. Más tarde se anotó la temperatura final (después de la combustión) antes de que iniciara el descenso, se midió la cantidad de alambre de platino que se quemó, transformándola a su equivalente en calorías (cada centímetro de alambre consumido de la bomba equivale a 2.3 cal g-1) para determinar las calorías liberadas. Esto se hizo mediante la siguiente formula:

Donde: E= energía liberada (cal.g-1); TF= temperatura final (°C); TI= temperatura inicial (°C); W= constante de la bomba calorimétrica; 10= longitud de alambre de platino (cm); E1= longitud de alambre consumido; PM= peso de la muestra (g).
En la fase 3 se calcularon las calorías y la Fc utilizando la metodología de Hipkins (1984). Las calorías producidas se pasaron a Julios por gramo (J.g-1), multiplicando por la constante 41 868 J. Acto seguido, los Julios por gramo se transformaron a moles de carbono, dividiendo por la cantidad de energía requerida por una planta de 4.8 x 105 J.mol-1 para fijar las moléculas orgánicas un mol de carbono. Por último, los Julios se convirtieron a gramos de C, multiplicando el peso molecular del C (12 gr) para calcular la fracción de carbono fijado con la siguiente formula:

Donde: Fc= fracción de carbono; GC= gramos de carbono (g); PM peso de la muestra (g); MS (%)= porcentaje de materia seca.
RESULTADOS
Composición florística
En las tres áreas estudiadas se identificaron un total de 2841 individuos en Pacurita (1501), Salero (1082) y Unguía (258). Estos individuos estaban distribuidos en 260 especies, pertenecientes a 207 géneros y 99 familias. Se observó una mayor riqueza florística en Pacurita y Salero. Esta diversidad resalta el estado de conservación del bosque primario, pues estas zonas registraron los valores más altos en cuanto a familias, géneros y especies, a diferencia de Unguía, donde se obtuvieron los valores más bajos. Cabe mencionar que esta última es un ecosistema dominado por la especie P. copaifera. Se logró evidenciar la abundancia de las familias Aracaceae en Salero y Pacurita, Fabaceae en Unguía y Pacurita, y Lecythidaceae en Unguía y Salero (Figura 1).

En las localidades de Pacurita y Salero, las especies presentaron IVI de 5.3 a 28.6. Sin embargo, en el corregimiento de Puerto, la especie seleccionada (P. copaifera) presentó un IVI de 202.6. En el corregimiento de Pacurita, las especies más representativas fueron P. pendula (28.6), Ambelania Sp (23.4) y Boehmeria nívea (19.7). En el corregimiento de Salero, las especies más representativas fueron C. argenteu, E. integrifolian y Otoba gardoniifolia, con IVI de 26.0, 22.1 y 13.4 respectivamente. Inga edulis y Wettinia quinaria fueron las únicas especies presentes en dos sitios diferentes (Pacurita y Salero) (Tabla 1).

Fracción de carbono
Se obtuvo una Fc promedio de 0.46, con un rango de 0.43-0.48.
En el corregimiento de Pacurita, las 23 especies con mayor peso ecológico presentes en las tres PPM presentaron una Fc promedio de 0.46, con un rango de 0.44-0.47. Esto, a fuerza de algunas especies que presentaron los valores más altos, como la E. integrifolia y C. webervaueri, mientras que B. nívea presentó el valor más bajo (Figura 2).
De igual manera, en el corregimiento de Salero, las especies presentaron una Fc promedio de 0.46. Es conveniente acotar que en esta área se encontraron las especies con mayor Fc (0.48 para M. gleasoniana) y menor Fc (0.43 para W. quinaria) de todas las zonas estudiadas (Figura 2).
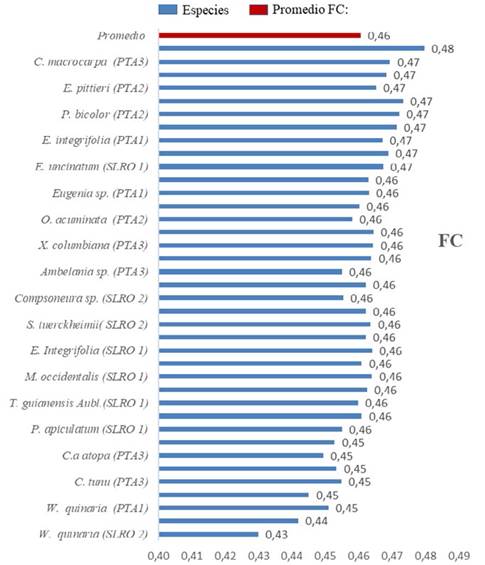
Figura 2 Fracción de carbono de las especies de mayor peso ecológico de los bosques de Pacurita y Salero en Chocó, Colombia. El color rojo, representa el promedio de Fc para 38 especies en los diferentes ecosistemas.
En la localidad de Puerto. La especie P. copaifera presentó una Fc igual al promedio de los bosques de los corregimientos de Pacurita y Salero (0.46).
DISCUSIÓN
Composición florística
Las tres zonas de estudio reportan en promedio 460 individuos/ha, distribuidos en 260 especies, 207 géneros y 99 familias, señalando una variación a nivel florístico y de diversidad en los bosques tropicales húmedos de los municipios de Quibdó, Unión Panamericana y Unguía, datos que son superiores a los hallados por Mena-Mosquera et al. (2019), quienes realizaron un estudio de composición florística en el bosque pluvial tropical de la subcuenca del río Munguidó, Quibdó, Chocó. En el mismo contexto, se reportan datos inferiores en términos de género, especies y familias en los estudios de diversidad en bosques húmedos tropicales realizados por Rojas et al. (2008), Segura-Madrigal et al. (2020) y Escobar (2013).
Asimismo, este estudio indica valores más representativos que los reportados por Mosquera-Ramos et al. (2007), quienes determinaron la composición florística del bosque húmedo tropical en los corregimientos de Pie de Pató, municipio del Alto Baudó, Chocó, Colombia. Estas diferencias pueden atribuirse a condiciones edáficas y climatológicas diferentes. Al comparar estas evidencias, se puede comprender que los ecosistemas de este estudio tienen una gran riqueza florística. A nivel de familias, la composición florística de los bosques estudiados es superior a la reportada por Henry y Bendlis (2017), quienes realizaron un análisis de la estructura y composición florística del bosque húmedo tropical de la comunidad de San Jerónimo.
Las familias botánicas mejor representadas en términos de abundancia de individuos en las treszonas de estudio fueron Fabaceae, Aracaceae y Lecythidaceae, demostrando una alta probabilidad de hiperdominancia de algunas de las familias que coinciden en la misma zona de influencia en bosques húmedos tropicales, de acuerdo con los resultados obtenidos por Mosquera y Hurtado (2014a) para la familia Arecaceae. Bennett y Jiménez (2018), en su descripción florística de un fragmento de bosque húmedo tropical en el área de Margarita (corregimiento de Cristóbal, provincia de Colón, Panamá), reportan un índice de valor de importancia por familia IVF que coloca a Fabaceae como una de las más representativas. Esto, debido a que los ecosistemas de esta investigación muestran índices muy bajos de intervención antrópica. Asimismo, se encontró una alta similitud en la abundancia de las familias Arecaeae y Fabaceae en estudios de muestreo de diversidad arbórea (Zarco et al., 2010; Escobar, 2013a).
Por otro lado, los resultados de esta investigación están por encima de los reportados por Tirado (2016) términos de familias, géneros y especies. Tirado (2016) instaló una parcela permanente de 1 ha en el bosque muy húmedo tropical del Sector de Angostura, Río Santiago, donde determinó la composición florística y la estructura de individuos con DAP ≥ 10 cm. En cuanto a las familias, ambas investigaciones reportan Fabaceae y Lecythidaceae en las diferentes zonas estudiadas. En términos del índice de valor de importancia, los estudios coinciden en las especies W. quinaria y P. veneralense para el corregimiento de Pacurita y B. alicastrum para Salero. Esto se debe a que los ecosistemas y las áreas muestreadas presentan varias similitudes; al realizar un muestreo en un bosque húmedo tropical en el departamento del Chocó, existe una alta probabilidad de registrar un número importante de individuos de estas especies.
Las cuatro especies de mayor peso ecológico en los bosques estudiados (B. utile, C. argenteum, P. veneralense y S. pittieri) coinciden con las reportadas por Quinto y Álvarez (2010) y Mosquera et al. (2012) para el departamento del Chocó.
Álvarez et al. (2021), en su estudio sobre la composición florística, la estructura y la diversidad arbórea de un bosque amazónico en Perú, reportan cinco especies forestales, i.e., P. pendula, T. guianensis, B. alicastrum, I. edulis, C argenteu, las cuales se encuentran dentro de las más representativas de esta investigación, que destaca los índices de valor de importancia de los individuos que presentaron los rangos más altos de Fc en los corregimientos de Parcurita y Salero.
Asimismo, Esteban et al. (2016), en su estudio de estructura y diversidad de tres asociaciones vegetales en el río San Juan, Chocó, Colombia, reportaron un alto índice de valor de importancia para las especies dominantes en tres fisiografías del Río San Juan en individuos con DAP> 10 cm. Entre las especies más comunes están O. gardoniifolia, T. guianensis, O. acuminata, W. quinaria, C. atopa, E. pittieri y P. bicolor. Esta riqueza de especies concuerda con los resultados de esta investigación para los corregimientos de Pacurita y Salero en árboles de terraza alta.
Fracción de carbono
Los resultados indican una Fc promedio de 0.46 para las muestras de ramas de las especies presentes en bosques húmedos, con un rango de 0.43-0.48. Estos datos son superiores a los reportados por Montero y Kanninen (2002) para un bosque muy húmedo tropical de la zona sur de Costa Rica, donde las Fc para fustes, ramas y hojas fueron de 0.48, 0.43 y 0.42 respectivamente. En este contexto, los bosques de los municipios de Quibdó, Unión Panamericana y Unguía tienen un alto contenido de carbono en la biomasa aérea, datos que coinciden con la investigación de Fonseca et al. (2021), quienes reportan valores de Fc en ramas de 0.39 para Gmelina arborea en plantaciones forestales.
La Fc varía dependiendo de las especies y las condiciones climatológicas de los ecosistemas. Es por ello que conocer la Fc de especímenes forestales contribuye a la mitigación del cambio climático, generando bases para precisar la capacidad de absorción, almacenamiento y fijación de carbono de estos ecosistemas. De igual manera, esto permite generar información sobre las condiciones climatológicas particulares de cada región, en aras de determinar el carbono almacenado en bosques tropicales húmedos. Este estudio aplica una fracción moderna que se encuentra por debajo del rango establecido por el ( IPCC 1996, IPCC 2006), quienes recomiendan un valor de 0.5 para los bosques naturales. Al utilizar la fracción recomendada para los bosques húmedos tropicales, se estaría sobrestimando la cantidad de carbono que estos ecosistemas pueden almacenar, por lo cual esta investigación sugiere la aplicación de una Fc de 0.46, de acuerdo con las condiciones particulares en la región.
Esta investigación obtuvo resultados similares a los obtenidos por Valera y Carvajal (2014), quienes determinaron el contenido de carbono o fracción de seis especies con rangos de Fc de 0.42 a 0.46 en bosques de los parques nacionales de Caracas, Venezuela. Esto difiere de la investigación realizada por Navarro (2011), que determinó la Fc por medio del peso específico para cuatro especies forestales en estadios sucesionales de bosques tropicales del corredor biológico Osa de Costa Rica. En dicho estudio, Apeiba tibourbou y Tetrathylacium macrophyllum se encuentran en los rangos de esta investigación, con valore de Fc de 0.40 y 0.48 respectivamente. Sin embargo, los otros individuos arrojaron resultados superiores a los de esta investigación, a saber: Guatteria amplifolia con 0.49 y Hyeronima alchorneoides con 0.51. Esto demuestra que las fracciones de carbono obtenidas con la metodología de poder calórico en este trabajo no se alejan de la media de 0.50 (o 50 %) de las cifras para biomasa, peso seco y FC propuestas por el (IPCC 1996, 2006).
Por su parte, Ticona (2012) determinó que la Fc en cada componente de la vegetación arbustiva Añahuaya Adesmia miraflorensis Remyfue de 0.49 en las hojas, 0.52 en el tallo y 0.44 en la raíz, con un promedio de 0.48 en la suma de los reservorios, datos que son congruentes en los aportes de este estudio.
En su estudio, Ragland et al. (1991) reportaron la Fc de cinco individuos de Guayabo (0.48); Guamo, Lirio, Churimo y Zanca araña (0.47); y Palma meme (0.43). Estos resultados son similares a los obtenidos en este trabajo, y cabe aclarar que las variaciones observadas se deben a las condiciones ambientales, el crecimiento de los árboles y, en especial, la composición química que presenta cada una de las especies. En ese sentido, las causas de la variación de la Fc se deben al contenido de lignina y los extractos de cada una de las maderas. Por ejemplo, los lípidos, la lignina y las proteínas muestran concentraciones elevadas de carbono, mientras que los ácidos orgánicos y minerales contienen poco o nada de carbono.
Otros estudios, tales como los desarrollados por Catpo (2004) y Lam (2011), reportan índices de Fc entre 0.42 y 0.48, intervalo que abarca los valores hallados en el presente estudio. Sin embargo, Lammlom y Savidge (2003) encontraron variaciones de Fc en especies forestales en un rango más amplio, que va de 0.47 a 0.59, asumiendo que cada tipo de madera tiende a ser química y anatómicamente única, por lo que es razonable que cada especie pueda tener una Fc característica. Esto puede estar influenciado más por la edad del árbol que por el diámetro del mismo.
De acuerdo con el método de calorimetría aplicado en esta investigación, los resultados de la fracción de carbono promedio están muy cerca a los resultados de Uceda (2015), quien aplicó el mismo método para 20 especies forestales de la Amazonía peruana, hallando un poder calorífico de 0.47 por la densidad promedio de todos los individuos, lo cual indica un alto nivel de similitud con las especies de los corregimientos de Salero y Pacurita.
Por otro lado, la investigación de Curo (2019), quien aplicó esta misma metodología de poder calorífico para determinar fracción aérea de bosques de relictos de Polylepis mediante la materia seca y la biomasa aérea seca de la especie, encontró Fc por individuo entre 0.42 y 0.44, con un valor promedio para la especies de 0.42, dato que está por debajo de los resultados de esta investigación.
Conclusiones
Los ecosistemas estudiados presentaron comportamientos diferentes en cuanto su estructura florística en términos de familias, géneros y especies, a pesar que se encuentran en condiciones climatológicas similares. Por otro lado, estos ecosistemas cuentan con gran diversidad, representada por un coeficiente de mezcla de 11/1, donde se registró una sola especie por cada 11 árboles inventariados. Las especies con mayor peso ecológico en las tres zonas de estudio fueron P. pendula, B. alicastrum, C. argenteu, E. integrifolian y P. copaifera. Otro rasgo importante es que las 39 especies con mayor IVI alcanzaron un promedio de fracción de carbono de 0.46 gr.cm3, dato que se constituye en una herramienta de utilidad para estimaciones más precisas y confiables de almacenamiento y fijación del carbono en bosques húmedos tropicales, como también para la determinación de emisiones evitadas de CO2 causa de la deforestación y degradación forestal.

