











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
El aumento constante de la población ha puesto en riesgo la seguridad alimentaria, por lo que se hace necesario el incremento de la productividad de los cultivos (Lattre-Gasquet et al. 2018). La papa es uno de los cultivos más importantes en Colombia, su producción se ha incrementado en un 8% durante los últimos 18 años y su productividad ha aumentado de 17,96t ha-1, en 2006, a 21,4t ha-1, en 2017; sin embargo, aún no se cumple con la producción necesaria para satisfacer la demanda nacional (Fedepapa, 2019).
Una de las estrategias para aumentar la productividad de los cultivos es reducir las pérdidas causadas por la presencia de plagas y de enfermedades, estimadas entre un 25 a 40%, de la producción total (Sobhy et al. 2015). El cultivo de papa es el producto de origen agrícola que, en Colombia, demanda la mayor cantidad de fungicidas e insecticidas (Gobernación de Antioquia, 2019).
Una alternativa ambiental y de bajo impacto utilizada en el control de plagas y de patógenos son los elicitores, los cuales, son compuestos que pueden ser aplicados a los cultivos, con el fin de fortalecer las plantas y aumentar su vigor, resistencia y rendimiento (Sobhy et al. 2015). La resistencia sistémica inducida es una forma no específica de resistencia a las enfermedades en las plantas, que actúa contra una amplia gama de patógenos y es activada por una amplia gama de elicitores no específicos (Bruce, 2014). Estas moléculas pueden ser, tanto de naturaleza microbiana de patrones moleculares asociados a patógenos como de las automoléculas o patrones moleculares asociados a daños (Aubel Van et al. 2016). La mayoría de los estudios, se han centrado en las vías de señalización subyacentes y la posible influencia en interacción planta-patógeno, mientras que poco conocimiento se tiene sobre el efecto de la aplicación de elicitores en condiciones de campo, en las variables de crecimiento y de producción del cultivo y en los costos de asignación que puede generar la resistencia sistémica adquirida (RSA) (Agrawal, 1999; Heil, 1999). Dentro de los llamados elicitores, se encuentran, principalmente, el ácido salicílico (AS) y el ácido jasmónico (AJ), que permiten una respuesta coordinada de toda la planta, contra posibles invasores (Pieterse et al. 2012). Al respecto, Navarre et al. (2003) encontraron que al aplicar AS no hubo un efecto claro en las plantas de papa, por lo que sugieren aclarar si los altos niveles basales de AS constituyen una defensa efectiva o, por el contrario, disminuyen la capacidad de respuesta al AS.
Por lo anterior, el objetivo de la presente investigación fue evaluar el efecto de un grupo de sustancias elicitoras sobre el crecimiento, el desarrollo y la producción del cultivo de papa variedad Diacol Capiro.
MATERIALES Y MÉTODOS
Este estudio, se llevó a cabo en la vereda el Toval, del municipio de Tutazá, departamento de Boyacá, Colombia, situada a 6°04´43,73´´ N y a 72°53´04,67´´ O, con una altitud de 3.243m s.n.m. y una temperatura promedio de 14°C. El cultivo seleccionado corresponde a la variedad Diacol capiro.
Se utilizó un diseño experimental en parcelas divididas con tres niveles, en donde cada uno de estos correspondió a la época de aplicación del elicitor, de acuerdo con el estado fenológico del cultivo, según escala fenológica BBCH, así: época uno (E1; formación de tallos secundarios), época dos (E2; floración) y época tres (E3; aplicación tanto en formación de tallos secundarios como en floración); lo anterior, acorde con Valbuena et al. (2009). Las subparcelas tuvieron cinco niveles, que fueron el elicitor empleado: glucosamina 1g L-1, Ácido γ-amino butírico (GABA) 1g L-1, Fosfito de Potasio 1,5ml L-1 y ácido salicílico (AS) 1g L-1 y el testigo sin aplicación.
Cada parcela tuvo un tamaño de 3m x 80m y cada subparcela 3m x 4m, lo que generó un total de 60 unidades experimentales, producto de multiplicar las tres épocas de aplicación por los cinco elicitores aplicados y cuatro repeticiones.
La toma de datos, se realizó a los 60, 80 y 100 días después de siembra (dds) a las variables: altura de la planta (cm), desde la parte basal hasta el foliolo más alto; número de foliolos por planta, contando la totalidad de foliolos expresado en unidades; clorofilas totales, por medio de un clorofilómetro SPAD-502 (Konica Minolta, Inc., Tokyo, Japón), para lo cual, se realizaron 4 mediciones por planta, en la tercera hoja bajando del tercio alto de la planta, de esta forma, al final de cada medición, se generó un promedio de clorofilas totales por unidad experimental, expresado en unidades SPAD. La masa seca, se determinó con una balanza VIBRA AJ220E (Shinko Denshi Co, Tokyo, Japon), después de cosechar los tubérculos y secarlos en mufla, a 75°C, hasta obtener peso constante. La producción total, se determinó pesando todos los tubérculos por unidad experimental y los resultados se llevaron a unidades de t ha-1. El número de tubérculos de calidad de primera, se estableció de acuerdo con los estándares establecidos por el mercado, riche (<4cm de diámetro), primera (entre 4 y 9cm de diámetro), cero (>12cm de diámetro).
La eficiencia fotosintética, se midió en campo, con el fluorómetro Junior-Pam (Heinz Walz GmbH, Effeltrich, Alemania), en 4 plantas por unidad experimental, para lo cual, se cubrió la tercera hoja bajando del tercio alto, con 2 bolsas de aluminio y una bolsa de papel, durante 60min, con el fin de generar la reducción de la actividad del PSII a valores cercanos a 0. Pasada una hora, se procedió a realizar las mediciones con el sensor del equipo, conservando la condición de oscuridad de la hoja.
Con los datos obtenidos, se realizaron pruebas de normalidad y de homogeneidad de varianzas; posteriormente, se llevó a cabo el análisis de varianza (Anova), para determinar diferencias estadísticas, con un grado de confiabilidad del 95% y se determinaron pruebas de comparación de promedios Tukey (P≤0,05). Los análisis, se realizaron con el software R versión 3.1.2.2 (R Team Core, 2015).
RESULTADOS Y DISCUSIÓN
Altura de la planta. Al comparar las épocas de aplicación y los tratamientos no se presentaron diferencias significativas, ni tampoco en la interacción a los 60 dds; no obstante, se presentaron diferencias a los 80 y 100 dds, para la interacción época por tratamiento, con valores promedio de 42 y 68cm, respectivamente. Es posible que esto ocurra, porque las plantas respondieron a las variaciones en la disponibilidad de fotoasimilados, a partir de la asignación y repartición de éstos entre los distintos órganos, ya que optimizan la captura de los mismos y maximizan, consecuentemente, la tasa de crecimiento (White et al. 2015).
En la interacción tratamiento por época, a los 80 dds, se presentan diferencias significativas (Figura 1A), en donde la aplicación de fosfito de potasio en la E1 presentó los mayores valores promedio de altura (47cm). Al respecto, Rossall et al. (2016) afirman que los fosfitos promueven el crecimiento de las plantas, incluso, en ausencia de patógenos vegetales, lo que concuerda con los resultados obtenidos en este trabajo; posiblemente, esto ocurre, porque el fosfito aplicado vía foliar es absorbido más rápidamente que los fosfatos, lo que hace que algunas funciones, como la síntesis de fitoalexinas y aminoácidos, sean más rápidas (Gómez-Merino & Trejo-Téllez, 2015). Además, el fosfito de potasio es capaz de producir un rápido estímulo de importantes procesos metabólicos en las plantas, implicados en la superación del estrés ambiental, patológico y nutricional (Cohen, 2002), debido a que el compuesto, al entrar en la planta, separa el anión del catión, dejando que el potasio contribuya con la mayor translocación de fotoasimilados a los órganos vertederos (hojas y tallos nuevos). Lo anterior coincide con los resultados hallados por Ortiz & Zapata (2011), quienes encontraron plantas de plátano más altas, al aplicar fosfito de potasio y afirman que este producto activa el sistema de defensa de las plantas y, al mismo tiempo, metaboliza el fosfito a fosfato, para utilizarlo como nutriente, desempeñando un papel importante en el crecimiento y el desarrollo del cultivo.

Figura 1 Altura de la planta de papa. a. 80 dds; b. 100 dds. AS: Ácido salicílico; GABA: Ácido γ-aminobutírico. Letras diferentes indican diferencias significativas entre tratamientos, según la prueba de Tukey (P≤0,05). Barras indican el error estándar (n=4).
Las plantas tratadas con glucosamina presentaron los valores más bajos de altura en la E1 y E3, posiblemente, al efecto que esta tiene en la elongación celular, ya que una de las características de este polímero es que influye en las propiedades funcionales de los sistemas en donde se adicione, pues interviene, de manera significativa, en el crecimiento de las plantas (Falcón et al. 2008).
A los 100 dds, se presentaron diferencias significativas para la interacción época por tratamiento (Figura 1B), siendo el tratamiento de GABA el que mostró las plantas con mayor altura, con un promedio de 76,5cm, en la E1 y las más bajas, en los tratamientos AS y glucosamina en la E3, mientras que para GABA, en la E2, las alturas no superaron los 62cm. Para la E2, al aplicar GABA a las plantas, el proceso involucrado en el crecimiento fue menor, ya que la competencia metabólica podría resultar en la variación de los costos de asignación. Caso contrario a lo reportado por Rose et al. (2003), quienes afirman que con la adición de 4% de quitina al medio, el crecimiento de las plantas mejoró de forma significativa y no solo actuó como inductor de respuestas de defensa de las plantas.
Número de foliolos. El número de foliolos no presentó diferencias significativas a los 60 dds y tuvo un promedio de 26, mientras que a los 80 y 100 dds sí se presentaron diferencias entre tratamientos (Figura 2A y 2B), épocas (Figura 2C) y la interacción época por tratamiento (Figura 2D), con promedios de 75 y 183, respectivamente. La E1 mostró los valores más altos, con 187 foliolos en promedio; no obstante, la aplicación en E3 tuvo el menor valor, con un promedio de 180 foliolos. Al respecto, la eficiencia en la producción de follaje puede estar afectada por factores internos de crecimiento relacionados con el genotipo y factores externos relacionados con el ambiente y las prácticas de manejo utilizadas, durante el ciclo de cultivo (White et al. 2015). Posiblemente, el gasto energético de las plantas fue menor en la producción de foliolos por la doble aplicación de elicitores, durante esta época, debido a que los elicitores inducen derivados del ácido salicílico (AS), los cuales, se acumulan en las plantas y pueden activar la expresión de genes específicos de patogénesis relacionada (PR), que no son inducidos por AS (Chen et al. 2009).

Figura 2 Número de foliolos por planta de papa. a. 80 dds; b. 100 dds; c. Clasificados por elicitores; d. Clasificados por épocas fenológicas de aplicación; BBCH E1: Formación de tallos secundarios, BBCH E2: Floración y BBCH E1 + BBCH E2: en formación de tallos secundarios y en floración. AS: Ácido salicílico; GABA: Ácido γ-aminobutírico. Letras diferentes indican diferencias significativas, según la prueba de Tukey (P≤0,05). Barras indican el error estándar (n=4).
Després et al. (2007) determinaron que los esfuerzos metabólicos en las plantas dependen de la etapa de crecimiento y la disponibilidad de recursos, que pueden causar costos de asignación, lo que se ratifica en este estudio, en donde la E3 tiene el menor número de foliolos, ya que la aplicación es tardía y se da en una etapa de mayor requerimiento de fotoasimilados. Para el caso de interacción entre tratamientos y épocas, a los 80 dds, se presentaron diferencias significativas; asimismo, el tratamiento de Glucosamina presentó una disminución notable del número de foliolos para la época 1 y 3.
A los 100 dds, se presentaron diferencias significativas en la interacción entre tratamientos y épocas, en donde las plantas con mayor número de folíolos fueron las del tratamiento testigo en las tres épocas, con un promedio de 193 y las plantas con menor número de foliolos fueron las que recibieron la aplicación de glucosamina, con 175 y 170 foliolos, en la E2 y E3, respectivamente. Es posible que la glucosamina en las plantas cree barreras físicas para la protección de la planta (Riveros, 2010), debido a que la glucosamina proviene de la ruta del ácido shikímico, que es el precursor de la fenilalanina y los fenil propanoides, los cuales, sintetizan la lignificación de tejidos, flavonoides y estilbenos, tanino suberinas y cutinas, además de la producción de fitoalexinas (Barber et al. 1999); Albersheim & Valent (1978) demostraron que oligómeros de glucanos de las paredes celulares son inductores activos de fitoalexinas, por ende, disminuye la producción de foliolos, ya que el gasto energético va dirigido a estos procesos metabólicos.
En un estudio de la interacción en tomate y la pudrición de corona de la raíz por patógeno F. oxysporum f. sp., Benhamou & Theriault (1992) demostraron que la aplicación de glucosamina foliar, para tratamiento de una infección de las raíces, aumentó, en gran medida, la formación de barreras físicas en los tejidos de la raíz; de hecho, el quitosano se ha utilizado para proteger una gama de huéspedes contra patógenos importantes, incluidos, los tubérculos de papa contra Erwinia carotovora y Rhizoctonia solani (Linden et al. 2000). La baja en el número de foliolos también se puede atribuir a que la aplicación de glucosamina genera un aumento de calcio citosólico libre y formación de H2O2, seguida de la expresión de genes, que codifican enzimas clave de la ruta de los fenilpropanoides y de PR (Aziz et al. 2007).
Clorofilas totales. No se presentaron diferencias significativas para las épocas de aplicación en ninguna de las evaluaciones, ya que los valores oscilaron entre 53 y 58 unidades SPAD, lo que concuerda con los resultados obtenidos por Africano & Pinzón (2014), quienes mencionan que es probable que no se afecten los fotosistemas, en especial, porque la relación fuente vertedero se ha mantenido y no ha comenzado a traslocar fotoasimilados.
Para la interacción tratamiento por época a los 60 y 80 dds, no se presentaron diferencias significativas, con valores promedio de 53 y 52 unidades SPAD, respectivamente, lo que indica que, posiblemente, no se han provocado daños en membranas de cloroplastos o degradación de las clorofilas, por la etapa fenológica en la que se encuentran o que no ha comenzado la traslocación a los tubérculos y la clorofila, aún se esta distribuyendo equitativamente entre las nuevas hojas, acorde con lo encontrado por Argentel et al. (2009), quienes mencionan que el contenido de clorofilas totales disminuyó a medida que la planta entra en fase reproductiva.
A los 100 dds (Figura 3) hubo diferencias significativas, en donde el menor número de clorofilas totales se presentó en las plantas que recibieron la aplicación de fosfito de potasio en la E2, con un promedio de 47 unidades SPAD, lo que, posiblemente, puede indicar que, debido a la competencia ejercida entre fosfitos y fosfatos por canales de entrada intracelulares, se produce una deficiencia de fosfato, con poco adenosin trifosfato (ATP), para desarrollar síntesis de clorofilas a partir del ácido glutámico, similar a lo reportado por Wu et al. (2011), quienes mencionan que, probablemente, la liberación de ATP fue bloqueada por los canales de aniones de la membrana, pero no por un canal específico de cloro.

Figura 3 Clorofilas totales en plantas de papa a los 100 dds. BBCH E1: etapa 1, formación de tallos secundarios; BBCH E2: etapa 2, floración. Letras diferentes indican diferencias significativas entre tratamientos, según la prueba de Tukey (P≤0,05). Barras indican el error estándar (n=4).
Eficiencia fotosintética (EF). No se presentaron diferencias significativas a los 60 dds, mientras que a los 80 dds, sí se observaron entre épocas. A los 100 dds hubo diferencias entre tratamientos y en la interacción tratamiento por época en la eficiencia fotosintética de las plantas de papa; sin embargo, la EF mostró una tendencia a disminuir hacia los 100 dds, con valores que oscilaron entre 0,78 y 0,75. En la época 2, la EF, a los 80 dds, tuvo un valor promedio para todas las aplicaciones, de 0,81, superior a los 0,77, registrados en la E3 (Figura 4A). Cabe mencionar que la fotosíntesis consta de dos fases principales, la fase fotoquímica y el ciclo de Calvin, por lo que cualquier factor que altere el normal funcionamiento de alguna de estas fases es causal del descenso de la EF; adicionalmente, los elicitores son reconocidos como reguladores endógenos en las plantas, al estar involucrados en muchos procesos fisiológicos de las plantas (Mishra et al. 2012).

Figura 4 Eficiencia fotosintética de plantas de papa. a. Clasificadas por días de medición; b. Clasificadas por épocas de aplicación de diferentes elicitores; BBCH E1: etapa 1, formación de tallos secundarios; BBCH E2: etapa 2, floración. Letras diferentes indican diferencias significativas entre el tiempo de medición, según la prueba de Tukey (P≤0,05). Barras indican el error estándar (n=4).
El tratamiento con aplicación de ácido salicílico mostró una tendencia ascendente en la EF de las plantas de papa en las dos primeras mediciones y presentó a los 100 dds valores ligeramente menores, a los exhibidos por las plantas del tratamiento testigo (Figura 4B). De igual forma, se aprecia que la aplicación de fosfito y glucosamina en las dos etapas, influye de forma negativa en la EF, ya que disminuye en un 7 y 5%, respectivamente, en relación con la aplicación de estos elicitores en una sola etapa.
Las plantas que recibieron AS, tanto en la E1 como en la E2, presentaron valores similares al testigo; sin embargo, cuando se aplicó AS en las dos etapas, la EF se redujo en 2,6%, debido a la doble aplicación de AS, la cual, probablemente, pudo haber generado estrés en las plantas de papa; lo anterior es similar a lo encontrado en semillas de arroz, por Le Thanh et al. (2017), quienes al evaluar la aplicación de AS obtuvieron un 28% más de producción de especies reactivas de oxígeno, frente al testigo, lo que es un indicativo de la situación de estrés en las plantas que origina el AS.
Biomasa seca. La medición de la masa seca mostró diferencias significativas en la interacción entre épocas y tratamientos. La masa seca del tratamiento testigo fue superior en un 20,8, 16,2, 24,1 y 11,7% a los tratamientos, cuyas plantas recibieron las aplicaciones de AS, GABA, glucosamina y fosfito, respectivamente, en donde la presencia de órganos de la planta, con una demanda neta por asimilados, es probable que altere, de gran manera, los patrones de producción y de distribución de la biomasa seca en los diferentes órganos de la planta (Dietrich et al. 2005).
El tratamiento con glucosamina presentó los valores más bajos para la producción de masa seca (Figura 5A). Al respecto, es probable que las plantas que fueron inducidas con glucosamina, posiblemente, gastaron mayor energía en la defensa que en la producción de biomasa seca, ya que este polímero ha sido extensamente evaluado, para determinar la capacidad de inducir respuestas de defensa natural en la planta, presentando cambios fisiológicos y bioquímicos que ocurren dentro de la planta, debido a la inducción por quitosano (Rodríguez-Pedroso et al. 2009).

Figura 5 a. Biomasa de plantas de papa; b. Producción total de plantas de papa sometidas a la aplicación de diferentes elicitores. Letras diferentes indican diferencias significativas entre elicitores aplicados, según la prueba de Tukey (P≤0,05). Barras indican el error estándar (n=12).
Del mismo modo, el AS también afectó, de forma significativa, la acumulación de masa seca de las plantas. Al respecto, es probable que la aplicación de AS actúe como un regulador sobre el balance óxido/reducción de las células vegetales; además, induce respuestas adaptativas, fisiológicas y morfológicas en las plantas, que incluyen el incremento en la producción de lípidos (Le Thanh et al. 2017), el aumento de la actividad enzimática de la peroxidasa y la polifenol oxidasa, así como el aumento en la producción de fitoalexinas, orizalexinas y de la actividad antioxidante (Li et al. 2012), lo cual, implica un desgaste de la planta y explica la menor generación de biomasa seca.
Producción total. Se presentaron diferencias significativas entre tratamientos. El testigo mostró la producción total más alta (Figura 5B), lo que implica que la aplicación de elicitores afecta la producción del cultivo. Las plantas de papa que recibieron la aplicación de AS y glucosamina fueron las de más baja producción (28,6 y 15,8% menos que el testigo, respetivamente), lo cual, puede ser atribuido a que estos dos elicitores promueven una mayor asignación de recursos a la defensa de la planta que a la generación de apios de arracacha.
De forma similar, Walters et al. (2005) afirman que los elicitores pueden facilitar la desviación de metabolitos y de energía del crecimiento y otros procesos hacia la defensa de la planta; también se menciona, que sistemas, como el de la producción de semillas, se puede ver afectado, debido a la variación en los costos de asignación, que provocan los elicitores (Thakur & Sohal, 2013; Dietrich et al. 2005). Otros efectos negativos que pueden causar los elicitores son el efecto autotóxico, ya que algunos de estos rasgos de resistencia son tóxicos para la planta y su expresión constitutiva puede imponer una carga metabólica adicional (Ghimire et al. 2019).
Las épocas de aplicación de los elicitores no revelaron diferencias significativas en la producción total de tubérculos de papa; no obstante, cuando se aplicaron los elicitores en la E1 (63,8t ha-1), los valores fueron 9,17 y 11,7%, mayores que la producción de las plantas tratadas en la E2 (57,9t ha-1) y en la E3 (56,33t ha-1), respectivamente.
Número de tubérculos de calidad primera (entre 4 y 9cm de diámetro). Los tubérculos de calidad primera mostraron diferencias en la interacción entre tratamientos y épocas (Figura 6), en donde la aplicación de fosfito de potasio generó la mayor producción en las tres épocas de aplicación, seguida por el tratamiento que recibió la aplicación de glucosamidas, que fue un 11,3, 3,48 y 4,22% inferior, para la E1, E2 y E3, respectivamente, lo que permite afirmar que la producción de la calidad primera se ve beneficiada por la aplicación del ión potasio, similar a Lobato et al. (2008), quienes mencionan que las plantas presentan un mayor verdor en la madurez del cultivo y, finalmente, la obtención de un mayor rendimiento de tubérculos (mayor tamaño y número de tubérculos).
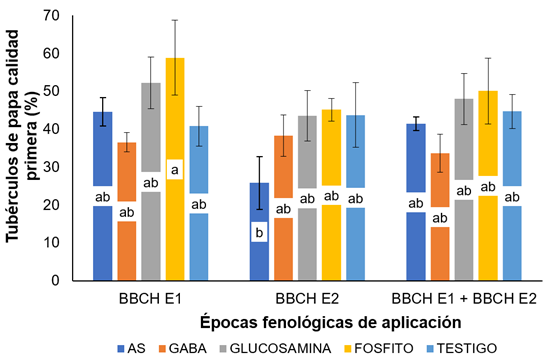
Figura 6 Porcentaje de tubérculos de papa de calidad primera (entre 4 y 9cm). BBCH E1: etapa 1, formación de tallos secundarios; BBCH E2: etapa 2, floración. Letras diferentes indican diferencias significativas entre tratamientos, según la prueba de Tukey (P≤0,05). Barras indican el error estándar (n=4).
Cabe resaltar, que, al aplicar los tratamientos en la E2, el porcentaje de tubérculos de papa, en promedio, disminuyó frente a las demás épocas de aplicación, lo que permite inferir, que esta etapa de desarrollo del cultivo es más sensible a los elicitores aplicados. Estos resultados son similares a los encontrados en un estudio diseñado específicamente para cuantificar los costos de asignación de SAR, en el cual, plantas de trigo tratadas con BTH, bajo condiciones libres de patógenos, mostraron una reducción significativa en la biomasa y la producción de semilla (Heil et al. 2000).
Se puede concluir que los elicitores afectan significativamente el crecimiento y el desarrollo del cultivo de papa variedad Diacol Capiro, en especial, cuando son adicionados a la planta, en dos aplicaciones en la E3, mientras que, si se realiza una sola aplicación de cualquier elicitor en la época 1, el crecimiento y el desarrollo del cultivo no se ve afectado. En cuanto a los parámetros fisiológicos del cultivo, el elicitor que menor repercusión tiene sobre el crecimiento y el desarrollo es el ácido salicílico, ya que tiene un comportamiento muy parecido al del testigo. La etapa de floración es la más sensible a los elicitores, debido a que estos redujeron el porcentaje de tubérculos de papa. En general, la aplicación de elicitores disminuyó la producción de papa.

