










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkCaldasia
Print version ISSN 0366-5232On-line version ISSN 2357-3759
Caldasia vol.30 no.2 Bogotá July/Dec. 2008
MIGUEL ANTONIO MURCIA-RODRÍGUEZ
MARTHA PATRICIA OCHOA-REYES
Programa de doctorado en Biología. Biodiversidad y Conservación. Instituto de Ciencias Naturales, Universidad Nacional de Colombia. miguel.murcia.r@gmail.com
Departamento de Biología y Química, Universidad de Pamplona, Norte de Santander, Colombia. martha.ochoa@unipamplona.edu.co
RESUMEN
Se determinó la respiración del suelo (RS) en una comunidad sucesional de pastizal de Holcus lanatus y Anthoxanthum odoratum en la franja altoandina de la cuenca del río Pamplonita, Norte de Santander, Colombia. Las medias mensuales de la RS fueron unimodales de día y de noche; la anual diurna fue de 118,44 mg CO2 m-2 h-1; y la nocturna, de 140,57 mg CO2 m-2 h-1. La diferencia entre estos dos valores se atribuyó a un incremento del brillo solar y a una disminución del recorrido del viento entre septiembre y diciembre de 2006 que condujo a un aumento de la tasa de evaporación. La RS nocturna se correlacionó con la humedad relativa y la temperatura máxima del aire y la diurna con el recorrido del viento. La RS acumulada (diurna y nocturna) mostró una tendencia lineal con respecto al tiempo. La producción anual diurna de carbono, con base en la RS acumulada anual, se estimó entre 32,67 y 40,73 y la nocturna entre 35,65 y 43,02 g de C m-2.
Palabras clave. Respiración del suelo, franja altoandina, sucesión ecológica, pastizal, Holcus lanatus.
ABSTRACT
Soil respiration (SR) was determinated in a successional weed community with Holcus lanatus and Anthoxanthum odoratum within the high Andean region in the Pamplonita river basin (Norte de Santander, Colombia). The SR monthly averages were unimodal both in the day and night; the diurnal average was 118.44 mg CO2 m-2 h-1; and the nocturnal one, 140.57 mg CO2 m-2 h-1. The difference between these two values was attributed to an increase in sunshine and a decrease in wind between September and December 2006, which, in turn, caused an increase in evaporation rate. Nocturnal SR was correlated with night maximum temperature and relative humidity and diurnal SR was correlated with diurnal winds. The day and night accumulated SRs showed a linear trend with respect to time. The diurnal annual carbon production based on the accumulated SR was estimated between 32.67 and 40.73 and the nocturnal one, between 35.65 and 43.02 g de C m-2.
Key words. Soil respiration, high Andean region, ecological succession, weed community, Holcus lanatus.
INTRODUCCIÓNLa respiración del suelo se define como la producción total de CO2, por unidad de área y de tiempo, en suelos intactos debido a la respiración de organismos edáficos, raíces, hifas micorrícicas, y, en menor extensión, por la oxidación química de los compuestos de carbono (Raich & Schlesinger 1992, Lloyd & Taylor 1994, Yi et al. 2007). Esta actividad metabólica se mantiene debido al equilibrio dinámico que existe entre las entradas de materia orgánica, procedente de la biomasa y del sistema radicular de la vegetación (Witkamp 1969, Raich & Schlesinger 1992) y las salidas por descomposición de la hojarasca y su eventual acumulación en el suelo (Schulze 1967). La respiración edáfica juega un papel crítico en la determinación de un amplio rango de fenómenos ecológicos que van desde el funcionamiento individual de las plantas hasta la concentración global del CO2 atmosférico (Liu et al. 2006). La respiración del suelo está regulada por una serie de factores bióticos y abióticos tales como la temperatura, el contenido hídrico, el inventario de nutrientes, la estructura de la vegetación, la actividad fotosintética o el desarrollo fenológico de la planta (Singh & Gupta 1977, Subke et al. 2006) así como por la biomasa de raíces finas y microbiana (Adachi et al. 2006). La respiración anual del suelo está relacionada directamente con la productividad primaria neta en turberas del subártico (Moore 1986) y con la producción de hojarasca en ecosistemas forestales (Schlesinger 1977, Raich & Nadelhoffer 1989, Davidson et al. 2002). En bosques tropicales, el factor que mayor influencia tiene sobre la variación temporal de la tasas de respiración del suelo no es tanto la temperatura edáfica sino el contenido de agua o la precipitación debido a que la temperatura del suelo es relativamente constante (Kursar 1989, Davidson et al. 2000). A escala global, las tasas anuales de la respiración edáfica están correlacionadas positivamente con las medias anuales de la temperatura del aire, la precipitación y la productividad primaria neta en diferentes tipos de vegetación (Raich & Schlesinger 1992).
En años recientes, se ha prestado mucha atención a la respiración edáfica, ya que este proceso ecológico se reconoce como la principal fuente de flujo de carbono procedente de la superficie del suelo y uno de los componentes cruciales dentro del ciclo del carbono en ecosistemas terrestres (Raich & Schlesinger 1992, Raich & Potter 1995). Se ha reportado que la respiración del suelo podría representar del 40 al 90% de la respiración de los ecosistemas forestales (Schlesinger & Andrews 2000). En una escala global, la respiración del suelo produce 80,4 Pg CO2-C anualmente (1Pg=1015g), el cual es aproximadamente 10 veces más alto que la combustión de compuestos fósiles y la deforestación combinadas (Raich et al. 2002). Así, pequeños cambios en la respiración del suelo pueden influir enormemente en la concentración del carbono atmosférico y en el balance calórico (Veenendaal et al. 2004, Kane et al. 2005) La respiración del suelo se ha convertido en el evento central de los cambios ecológicos globales debido a su papel controversial en los procesos de calentamiento global (Giardina & Ryan 2000) ya que contribuye a determinar si un ecosistema dado se comporta como fuente de carbono o sumidero de CO2 (Jassal et al. 2007).
En Colombia, la franja alto andina (3000-3200 m) o Bosque altoandino (Rangel 2000) del nororiente del país ha sido transformado dramáticamente por el hombre para ser utilizado como zonas de pastoreo (pastizales) y de siembra de cultivos agrícolas como papa, trigo y especies forestales como Pinus patula. Sin embargo, en la última década, la comunidad rural aledaña a las zonas perturbadas y las entidades estatales como CORPONOR se han preocupado por la recuperación y conservación de los nacimientos de las quebradas que dan origen al río Pamplonita. Además, a través del convenio Universidad de Pamplona-COLCIENCIAS se han podido desarrollar estudios de restauración ecológica de los ecosistemas estratégicos de la cuenca alta del río Pamplonita. Este estudio trata de explicar el proceso dinámico que acontece en los primeros estadios sucesionales del bosque altoandino tendientes a restablecer los atributos estructurales y funcionales de este orobioma. Nuestro principal objetivo fue establecer el comportamiento de la respiración edáfica y su relación con los parámetros meteorológicos para entender cuáles son los factores que regulan los flujos del CO2 suelo-atmósfera en un ciclo anual en una comunidad de Pastizal de 10 años sin intervención agrícola.
MATERIALES Y MÉTODOS
Área de estudio
El presente estudio se realizó en la Reserva Natural El Volcán, Municipio de Pamplona (Norte de Santander, Colombia). Esta reserva se localiza en la cuenca alta del río Pamplonita, entre 2970 y 3450 m de altitud, tiene una extensión aproximada de 590 hectáreas, con coordenadas geográficas (N 7º 20’ 57.4’’, W 72º 42’ 12.2’’). Esta cuenca presenta zonas de vida altoandina y de páramo con diferentes grados de perturbación ocasionados por la rotación de la actividad agrícola y ganadera. La franja altoandina se caracteriza por presentar laderas con pendientes pronunciadas que configuran cañones angostos donde persisten relictos de bosque primario. En sectores con pendientes moderadas se desarrollan comunidades herbáceas, de origen antrópico, dominadas por Holcus lanatus, Anthoxanthum odoratum, Hypochaeris radicata y Rumex obtusifolius. Estas comunidades corresponden a etapas priserales de bosque altoandino denominadas en este estudio como pastizales. La zona de pastizal o potrero, donde se realizaron las mediciones de la respiración edáfica, comprende una extensión aproximada de (56,5) ha, la cual se encuentra en estado de barbecho desde el año 1997. Actualmente, el área de estudio exhibe una serie de núcleos de expansión o espacios colonizados por elementos arbustivos como Lepechinia conferta, Monochaetum myrtoideum, Rubus macrocarpa, Baccharis prunifolia, Hypericum lancioides, H. Phellos y Hesperomeles glabrata (Sánchez et al. 2007).
Los registros meteorológicos diarios fueron obtenidos a partir de la estación climatológica del Instituto Superior de Educación Rural (ISER-IDEAM) ubicada a 2340 m, perteneciente a la corriente del río Pamplonita.
Respiración del suelo
La estimación de respiración del suelo (RS) se llevó a cabo de acuerdo con la metodología de la “caja invertida” propuesta por Walter & Haber (1957), descrita por Medina & Zelwer (1972), y empleada por Cavelier & Peñuela (1990) para bosques de niebla y caducifolios en la serranía de Macuira, Colombia. Este método consistió en estimar el comportamiento del dióxido de carbono en el suelo utilizando frascos de compota en los cuales se depositó un volumen de 10 ml de solución de KOH 0,5 N. Estos frascos se cubrieron con cilindros de acero inoxidable 30 cm de longitud y 15 cm de diámetro; existiendo una relación de 15:1 entre una emisión aérea (suelo) y la absorción aérea (solución). Los frascos se colocaron sobre un trípode metálico aproximadamente a 2 – 3 cm por encima del suelo y los cilindros se sellaron con suelo alrededor del margen más bajo. Se realizaron cinco (5) réplicas, distribuidas al azar. Los cilindros invertidos se dejaron expuestos durante el día (desde las 6 horas hasta las 18 horas) y la noche (desde las 18 horas hasta las 6 horas) para determinar el comportamiento del CO2. Después de la exposición, el CO2 fijado por el KOH se determinó por titulación con solución de H2SO4 0,1 N, usando un titulador automático 702 SM Tritino. Los muestreos se realizaron mensualmente, durante la estación seca y húmeda, por un período de 12 meses. El valor estimado por triplicado fue multiplicado por un factor de 4/3 ya que la producción de CO2 representa el 75% de la evolución actual del CO2 medido por análisis infrarrojo (Haber 1958, Medina et al. 1980).
Análisis estadístico
Los resultados se analizaron con base en un modelo lineal general univariante (conocido como GLM-U en la literatura anglosajona) para determinar el efecto de la interacción temporal (mes-período) sobre la respiración del suelo (Zuur et al. 2007). La relación entre la respiración edáfica media mensual y los parámetros meteorológicos se llevaron a cabo a través de un análisis de correlación paramétrico (Pearson) y no paramétrico (Kendall y Spearman). Para establecer el efecto de las variables ambientales sobre la dinámica de la respiración diurna y nocturna se realizó un análisis de regresión lineal múltiple. La selección de las variables independientes se determinó mediante el método de pasos sucesivos (Hair et al. 1998, Vittinghoff et al. 2005). Para incrementar la potencia del modelo diurno y nocturno, la variable predictiva se ajustó a un modelo de mínimos cuadrados ponderados MCP (Ramírez & Ramírez 2006). La tasa anual de la respiración del suelo se estableció, con un intervalo de confianza del 95%, a través de un análisis de regresión lineal donde la variable dependiente (y) fue la respiración del suelo acumulada (mg CO2 m-2 h-1) y la variable independiente (x) fue el tiempo medido en días. La variable acumulada (y) se creó con el fin de establecer sí la tasa de cambio de la respiración del suelo exhibe un crecimiento constante a lo largo del año (Hernández & Murcia 1995). La producción neta de Carbono se estimó multiplicando la tasa anual de la respiración del suelo por el área del Pastizal. Se utilizó el programa estadístico SPSS versión 16.0.
RESULTADOS
Clima
La marcha anual de la precipitación y de la evaporación mostraron un comportamiento bimodal altamente contrastante (r=-0,624, p=0,03, n=12, la relación entre estas dos variables es lineal e inversamente proporcional, si aumenta la precipitación se espera que la evaporación disminuya). Los valores más altos de la precipitación se presentaron en marzo y octubre 156,7- 165,2 mm; mientras la evaporación más alta ocurrió en agosto y febrero 118,6-111,1 mm (Fig. 1). La humedad relativa del aire (HR%) se correlacionó inversamente con la temperatura máxima del aire (Tmáx) y con el recorrido del viento (r=-0,668, p=0,018; r=-0,614, p=0,034). El brillo solar mostró el mayor grado de asociación con la evaporación (r=0,859, p<0,001). El comportamiento general de los parámetros meteorológicos configuraron un año de marcada estacionalidad en lo referente a la precipitación y a la (HR%), lo cual condujo a un desbalance hídrico (-128,1 mm) entre la entrada por lluvia y la pérdida por evaporación.

Variación temporal de la respiración del suelo
Las medias mensuales diurnas y nocturnas de la respiración del suelo (RS) fueron unimodales (Fig. 2) y significativamente disímiles a lo largo del año (Tabla 1 y Tabla 2): las mayores en junio, 320,91 y 253,32 mg CO2 m-2 h-1, respectivamente, y las menores en mayo, 56,03 y 54,51, pese a la precipitación relativamente alta en este mes (120,9 mm). La RS diurna fue más contrastante que la nocturna, ya que junio, agosto y enero mostraron la mayor actividad metabólica. Mientras que la respiración nocturna tendió a ser homogénea, la menor actividad fue en mayo, marzo, abril y febrero; la mediana en enero, julio y noviembre; y la máxima en octubre, diciembre, septiembre, agosto y junio.


Tabla 2. Distribución por periodo (día-noche) de la respiración mensual del suelo en subconjuntos con promedios homogéneos. Tukey HSD (N=5).

El análisis de la RS mensual mostró una distribución metabólica más definida (Tabla 3): junio y agosto, con 287,12 y 218,87 mg CO2 m-2 h-1, constituyen el subconjunto de máxima actividad (p>0,05). La tasa promedio anual de la RS nocturna (140,57) superó ampliamente la tasa media de la RS diurna (118,44) en 22,13 mg CO2 m-2 h-1 (p<0,01, n=60). Esta diferencia se incrementó entre septiembre y diciembre, cuando la RS nocturna se mantuvo cerca de 167,7 mg CO2 m-2 h-1 y la RS diurna decreció alrededor de 72,1 mg CO2 m-2 h-1 (Fig. 2), debido posiblemente a un descenso en el recorrido promedio del viento (Fig. 3), factor meteorológico asociado directamente con el incremento de la RS diurna (Tabla 4).



Efecto del clima sobre la RS
La humedad relativa del aire (HR%) se correlacionó inversamente con la RS nocturna (r=-0,642, p< 0,05), es decir, que por cada unidad de aumento que exhibe la HR, la RS disminuye en 10,04 mg de CO2 m-2 h-1 (Fig. 4). Mientras que, la temperatura máxima del aire (Tmáx) se relacionó directamente con la RS nocturna (r=0,61, p<0,05), lo cual equivale a que por cada grado centígrado que se incremente la Tmáx, la RS aumenta en 55,69 mg de CO2 m-2 h-1 (Fig. 5). Estos resultados nos permiten identificar que los cambios en el comportamiento de la temperatura máxima del aire pueden afectar en mayor proporción (5,5:1) el incremento de las tasas de respiración nocturna del suelo más que la humedad relativa del aire. El recorrido del viento en Km (Viento) se correlacionó significativamente con la RS diurna (rho=0,594, p< 0,05), lo cual puede indicar que la velocidad del viento afecta la aireación del sustrato superficial del suelo incrementando el flujo de CO2.


En el ámbito circadiano, la HR mostró un efecto inverso más determinante sobre la dinámica respiratoria (r=-0,658, p<0,05): en agosto, la HR media más baja (72,81) coincidió con uno de los picos de mayor RS (218,87) mg de CO2 m-2 h-1; y en febrero, la HR más alta (84,43) concordó con una de las menores tasas de respiración 79,95 mg de CO2 m-2 h-1.
Tasas de cambio de la RS
La respiración del suelo acumulada (RSA) o tasa de respiración anual (diurna y nocturna) está relacionada directamente con el tiempo, a través de un modelo de regresión lineal (Fig. 6):
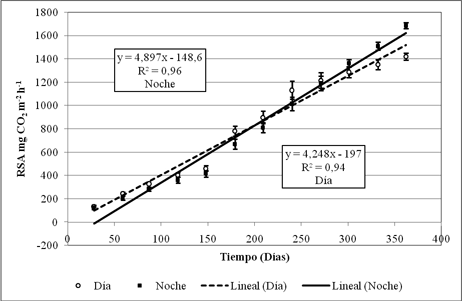
Ecuación (1): RSAd = 4,248D – 19,7
(R2=0,94, p<0,0001, n=60)
Ecuación (2): RSAn = 4,897D – 148,6
(R2=0,96, p< 0,0001, n=60)
Donde RSA, es la respiración del suelo acumulada promedio (d=diurna, n=nocturna) expresada en mg de CO2 m-2 h-1, y D el tiempo (días). El valor del intercepto para el período diurno no es significativamente diferente de cero (t=-0,629, p=0,532); pero si para el periodo nocturno (t=-5,136, p<0,001).
Según las ecuaciones de regresión (1 y 2), la tasa anual de respiración del suelo, para el periodo diurno, se calculó entre 32,76 y 40,73 g de C m-2 año-1, y para el nocturno entre 35,65 y 43,02 g de C m-2 año-1. Estos bajos estimativos son relativamente similares a los obtenidos por varios autores en ecosistemas de tundra, pantanos y pasturas de Brachypodium sp (Tabla 5).

La intersección entre las ecuaciones (1 y 2) acontece a mediados de julio (194 días) cuando la RSA alcanza los 800 mg de CO2 m-2 h-1. Este evento concuerda con el momento en que la HR y la Tmáx del aire se incrementan, y con el instante en que el recorrido del viento desciende abruptamente (Fig. 2 y Fig. 3). Los dos primeros factores ambientales regulan la RS nocturna, mientras el tercero disminuye la RS diurna. Sin embargo, la tendencia de la Tmáx podría explicar la mayor razón de cambio de la RSA nocturna (4,9) con respecto a la diurna (4,3).
La producción neta de C en la comunidad sucesional de pastizal, de la franja altoandina, podría calcularse entre 20,76 y 22,22 Toneladas de C año-1 para el periodo diurno y nocturno.
Modelación de la RS
La RS diurna y nocturna se ajustó a un modelo de mínimos cuadrados ponderados mediado por la precipitación (Fig. 2):
Ecuación (3): RSD = 962,791 – 10,873(HR)
Ecuación (4): RSN = 920,434 – 10,069(HR)
Donde RSD, es la respiración diurna del suelo estimada y RSN la nocturna expresada en mg de CO2 m-2 h-1, y (HR), la humedad relativa del aire promedio mensual. La significancia de los parámetros y los estadísticos de prueba para la comprobación de la idoneidad de las ecuaciones (3 y 4), a través del análisis de residuos, indican que todos los supuestos se cumplen satisfactoriamente (Tabla 6), y por lo tanto, la variación explicada por el modelo se debe a los cambios en la HR. Sin embargo, la HR solo determina el 35 y el 46 (%) de la variación total de la RS diurna y RS nocturna respectivamente. El porcentaje restante (65 y 54) podría ser explicado por otras variables de tipo endógeno como el desarrollo y la morfo-fisiología de las especies vegetales, así como por las propiedades fisicoquímicas del suelo.

A pesar de la complejidad del fenómeno ecológico de la respiración del suelo, la acción de la HR logra estimar proporcionalmente las oscilaciones abruptas mensuales del comportamiento diurno y nocturno en la comunidad de pastizal altoandino.
DISCUSIÓN
Los resultados muestran una clara estacionalidad de la respiración del suelo en el periodo nocturno en concordancia inversa y directa con el patrón de la humedad relativa y el brillo solar. El incremento en las horas de brillo solar observado entre agosto y diciembre se relaciona con los mayores valores de respiración del suelo nocturno; este comportamiento concuerda con lo descrito por (Walter 1960, Lieth & Oullette 1962, Medina 1966, Cavalier 1990) quienes observaron que, después de días soleados, los valores nocturnos de la respiración edáfica eran generalmente mayores que los diurnos. Los días con alta radiación conducirían a un aumento de los valores de la respiración radicular necesarios para el adecuado transporte de asimilados (Walter 1960). Sin embargo, esta diferencia es producto de la alta transpiración foliar, la cual arrastraría CO2 hacia las hojas, procedente de la respiración radicular, debido a que hay mucha evaporación durante el día (Haber 1958). Los valores de la respiración edáfica nocturna eran más altos en días soleados y que en los días nublados las diferencias eran mínimas (Hernández et al. 1995). Así, durante los meses de marzo a mayo se observaron los valores más bajos de la respiración del suelo en relación con los valores más altos de la precipitación y los más bajos de la evaporación (Fig. 1). El incremento en el contenido de la humedad del suelo podría permitir un aumento en la resistencia del suelo a la difusión de gases inhibiendo el desplazamiento del CO2 del suelo hacia la atmósfera (Cavalier & Goldstein 1989, Cavalier 1990).
La Tmáx tendió a incrementarse linealmente con el tiempo a razón media de 0,004 °C por día (R2 = 0,469, p<0,05, n= 12), acoplada con los aumentos de las tasas de respiración edáficas, lo cual significa que un leve aumento en la tasa de cambio de la Tmáx podría incrementar significativamente la razón de cambio anual de la RS en las diferentes comunidades de pastizales que persisten en la reserva El Volcán. Según Houghton et al. (1990) y Raich (1992), la relación entre la respiración del suelo y la temperatura es esencial para investigar como los cambios de la temperatura global pueden influir sobre las tasas de respiración del suelo, y sugieren que una correlación significativa podría indicar un incremento en la acumulación de gases invernadero.
El descenso de la precipitación y del recorrido del viento propicia un aumento en las horas de brillo solar, el cual ejerce un efecto sobre las condiciones de la HR, la Tmáx y, por ende, ocasionan un aumento en las tasas de evaporación. Este incremento evaporativo posiblemente explique el comportamiento atípico de la respiración nocturna entre los meses de Septiembre a diciembre, época en la cual los niveles de precipitación disminuyeron abruptamente generando un incremento en los valores promedios de la (Tmáx) favoreciendo un flujo mayor de CO2 hacia la atmósfera ya que la baja cobertura vegetal de esta comunidad no tiene una buena capacidad de retención térmica durante las noches despejadas propias de este periodo seco (Murcia 2001, Murcia & Mora 2003). Esta cobertura vegetal se caracteriza por tener casi exclusivamente elementos herbáceos del grupo de las gramíneas, tipo macolla, conformados por Holcus lanatus y Anthoxanthum odoratum, con valores que oscilan entre 20 y 60 % respectivamente.
La tasa anual de la RS mostró una tendencia hacia el incremento de la producción nocturna del CO2 hacia el final del periodo de estudio. Este comportamiento refleja el cambio de las condiciones ambientales y el efecto que tiene el incremento de la radiación global sobre el calentamiento de la superficie del suelo y posiblemente sobre las tasas de transpiración momentáneas de la comunidad herbácea. Además, las bajas temperaturas que se presentan en las épocas más secas del año pueden inhibir las tasas de respiración del suelo en estas comunidades sucesionales del bosque altoandino. De igual forma, los bajos niveles de precipitación multianual (menores de 892,8 mm) afectan la actividad de los microorganismos del suelo y las tasas de respiración radicular de las especies de gramíneas.
La acentuada fluctuación diaria o ciclo circadiano de los parámetros meteorológicos, propia de los ecosistemas de alta montaña tropical, configuran tasas contrastantes en el día y en la noche (Mora & Sturm 1995). Estos cambios intempestivos del clima y del fotoperiodo tienen como consecuencia el lento crecimiento de las especies de plantas debido al fuerte descenso del potencial hídrico del suelo que induce una sequía fisiológica (Mora 2002, 2004). Es posible que esta sequía se incremente y se prolongue durante los meses en los cuales las condiciones ambientales son más extremas (Murcia 2001, Murcia & Mora 2003). Por tal razón, la tasa anual de respiración edáfica de este pastizal sucesional es similar a los encontrados en comunidades de líquenes, juncos y arbustos de la tundra en Alaska (Poole & Millar 1982, Giblin et al. 1991), pantanos ombrotróficos en Suecia (Svenennson 1980), áreas dominadas por Sphagnum en Inglaterra (Clymo & Reddaway 1971) y pastizales de Brachypodium en Francia (Billes et al 1975).
En general, el comportamiento de la respiración edáfica de esta comunidad priseral de pastizal se ve reducida por la pérdida de la cobertura vegetal y por el deterioro de la capa orgánica del suelo del bosque nativo, el cual fue transformado para el desarrollo de las actividades agrícolas y ganaderas. A pesar de los diez años sin intervención humana (1997-2007) esta comunidad no la logrado alcanzar un estrato subarbustivo que pueda atenuar los cambios intempestivos del clima sobre la superficie del suelo mediante un aporte significativo de hojarasca al suelo (Murcia & Ochoa en preparación). Según hemos podido establecer, el aporte de hojarasca en diferentes comunidades sucesionales del bosque altoandino conduce a una disminución de las tasas de respiración diurnas e incrementa las tasas de respiración nocturnas a lo largo del gradiente sucesional: matorral, rastrojo, bosque secundario y bosque maduro.
Las actividades de pastoreo, la siembra intensiva de papa y trigo, y el leñateo, generados por más de 300 años, provocaron notables cambios estructurales y funcionales en el suelo que han impedido el desarrollo de las etapas priserales del bosque altoandino en la Reserva El Volcán.
CONCLUSIONES.
La humedad relativa y la temperatura máxima del aire en el periodo nocturno y el recorrido del viento en el diurno tuvieron mayor efecto sobre el flujo de CO2 en la superficie del suelo de la comunidad de pastizal debido particularmente a la cobertura vegetal baja. Las tasas de respiración del suelo estimadas en este estudio son relativamente pequeñas pero continuas a lo largo del año y semejantes con las de otros ecosistemas, en los cuales, las bajas temperaturas son el principal factor de regulación del crecimiento vegetativo y, por ende, de la respiración radicular y microbiana.
AGRADECIMIENTOS.
Queremos expresar nuestro agradecimiento a COLCIENCIAS por el apoyo financiero que brindó al proyecto de investigación “Restauración ecológica de la selva altoandina en la cuenca del río Pamplonita”. A los directivos de la Universidad de Pamplona, especial reconocimiento al Sr. Dr. Peñaranda rector de la Universidad por el apoyo financiero y logístico. Al IDEAM por las base de datos de la estación meteorológica del ISER, municipio de Pamplona. Al biólogo Luís Roberto Sánchez por la determinación del material vegetal y por su trabajo en campo. Al biólogo Fidel Ernesto Poveda por la asesoría en la interpretación y el análisis tanto estadístico como conceptual de la información y la estructuración crítica del manuscrito. Al Dr. Víctor Barradas por sus comentarios en algunos apartes climáticos y edáficos del texto. Un especial agradecimiento Al Prof. Dr. Orlando Rangel por la revisión, los comentarios y las correcciones de este manuscrito.
LITERATURA CITADA
1. ADACHI, M., Y.S. BEKKU, W. RASHIDAH, T. OKUDA & H. KOIZUMI. 2006. Differences in soil respiration between tropical ecosystems. Applied Soil Ecology 34, 258-265. [ Links ]
2. BILLES, G., J. CORTEZ, P. LOSSAINT, F. LAFONT & J. PRIOTON. 1975. Etude comparative de l’activité biologique des sols sous peuplements arbustifs et herbacés de la garrigue méditerranéenne. (I). Minéralisation du carbone et de l’azote. Rev. Ecol. Biol. Sol. 12, 115-139. [ Links ]
3. BILLINGS, W.D., K.M. PETERSON & G.R. SHAVER. 1978. Growth, turnover, and respiration rates of roots and tillers in tundra graminoids. In: Vegetation and production ecology of Alaskan artic tundra (ed. L. L. Tieszen). Nueva York: Springer-Verlag, 415-434. [ Links ]
4. CHANG, S.C., K.H. TSENG, Y.J. HSIA & C.P. WANG. 2008. Soil respiration in a subtropical montane cloud forest in Taiwan. Agricultural and Forest Meteorology. 148, 788-798. [ Links ]
5. CAVELIER, J. & G. GOLDSTEIN. 1989. Mist and fog interception in tropical elfin cloud forests. J. Trop. Ecol. 5: 309-322. [ Links ]
6. CAVELIER, J. & M.C. PEÑUELA. 1990. Soil respiration in the cloud forest and dry deciduous forest of Serranía de Macuira, Colombia. Biotropica 22(4): 346-352. [ Links ]
7. CLYMO, R.S. & E.J.F. REDDAWAY. 1971. Productivity of Sphagnum (bog-moss) and peat accumulation. Hidrobiologia 12: 181-192. [ Links ]
8. DAVIDSON, E.A., L.V. VERCHOT, J.H. CATTANIO, I.L. ACKERMAN & J.E.M. CARVALHO. 2000. Effect of soil water content of soil respiration in forests and cattle pastures of eastern Amazonia. Biochemistry 48: 53-69. [ Links ]
9. DAVIDSON, E.A., K. SAVAGE, P. BOLSTAD, D.A. CLARK, P.S. CURTIS, D.S. ELLSWORTH, P.J. HANSON, B.E. LAW, Y. LUO, K.S. PREGITZER, J.C. RANDOLPH & D. ZAK. 2002. Belowground carbon allocation in forest estimated from litterfall and IRGA-based soil respiration measurements. Agricultural and Forest Meteorology 113: 39-51. [ Links ]
10. GIARDINA, C.P. & M.G. RYAN. 2000. Evidence that decomposition rates of organic carbon in mineral soil do not vary with temperature. Nature 404: 858. [ Links ]
11. GIBLIN, A.E., K.J. NADELHOFFER, G.R. SHAVER, J.A. LAUNDRE & A.J. MCKERROW. 1991. Biogeochemical diversity along a riverside toposequence in artic Alaska. Ecol. Monogr. 61: 415-435. [ Links ]
12. HABER, W. 1958. Okologische untersuchugen der bodenatmung. Flora 146: 109-157. [ Links ]
13. HAIR, J.F., R.E. ANDERSON, L. TATHAM, & C. BLACK. 1998. Multivariate data analysis. Prentice Hall, New Jersey. [ Links ]
14. HERNÁNDEZ, I., E. MEDINA & D. LÓPEZ. 1995. Respiración edáfica y aportes de materia orgánica por las raíces y la hojarasca en un cultivo de caña de azúcar. Agronomía Trop. 45(1): 121-142. [ Links ]
15. HERNÁNDEZ, M. & M.A. MURCIA. 1995. Estimación de la productividad primaria de Espeletia grandiflora H & B y Pinus patula Schl & Cham en el páramo “El Granizo” Cundinamarca, Colombia. En: Estudios ecológicos del páramo y del bosque altoandino cordillera oriental de Colombia. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Jorge Álvarez Lleras 6: 503-520. [ Links ]
16. HOUGHTON, J.T., G.J. JENKINS & J.J. EPHRAUMS. 1990. Climate change: The IPCC assessment. Cambridge/U.K.: Cambridge Univ. Press. [ Links ]
17. JASSAL, R.S., T.A. BLACK, T. CAI, K. MORGENSTERN, Z. LI, D. GAUYMONT-GUAY & Z. NESIC. 2007. Components of ecosystems respiration and an estimate of net primary productivity of an intermediate-aged Douglas-Fir stand. Agric. For. Meteorol. 144: 44-57. [ Links ]
18. KANE, E.S., D.W. VALENTINE, E.A.G. SCHUUR, & K. DUTTA. 2005. Soil carbon stabilization along climate and stand productivity gradients in black spruce forests of interior Alaska. Can. J. Forest Res. 35: 2118-2119. [ Links ]
19. KURSAR, T.A. 1989. Evaluation of soil respiration and CO2 concentration in a lowland moist forest in Panama. Plant Soil 113: 21-29. [ Links ]
20. LIETH, H. & R. OULLETTE. 1962. Studies on the vegetation of the Gaspe Peninsula. II The soil respiration of some plants communities. Can. J. Bot. 40: 127-140. [ Links ]
21. LIU, H., LI, L., HAN, X., HUANG, J., SUN, J. & WANG, H. 2006. Respiratory substrate availability plays a crucial role in the response of soil respiration to environmental factors. Applied Soil Ecology 32: 284- 292. [ Links ]
22. LLOYD, J. & A. TAYLOR.1994. On the temperature dependence of soil respiration. Functional Ecology. 8: 315-323. [ Links ]
23. MEDINA, E. 1966. Producción de hojarasca, respiración edáfica y productividad vegetal en bosques deciduos de los llanos altos centrales. In: Prog. Biol. del suelo. Monografía I. Centro Coop. Cient. UNESCO Amer. Lat. E A. Rapport (Ed). Montevideo. Pags. 97-108. [ Links ]
24. MEDINA, E., H. KLINGE, H.C. JORDAN, & R. HERRERA. 1980. Soil respiration in amazonian rain forest in the Rio Negro basin. Flora 170: 240-250. [ Links ]
25. MEDINA, E. & M. ZELWER. 1972. Soil respiration in tropical plants communities. In: PM. Golley and F. B. Golley (Eds). Proceedings of the second international symposium of tropical ecology, Athens, Georgia, pp 245-269. University of Georgia Press, Athens, Georgia. [ Links ]
26. MORA-OSEJO, L.E. 2001. Contribuciones al estudio comparativo de la conductancia y de la transpiración foliar de especies de plantas del páramo. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Jorge Álvarez Lleras Nº 17. [ Links ]
27. MORA-OSEJO, L.E. 2002. El ciclo climático circadiano, los cambios intempestivos del clima durante el fotoperiodo y las respuestas adaptativas de las plantas del páramo. Simposio: Cambio climático y su potencial impacto en los páramos, pp. 132-143. I Congreso Mundial de Páramos. [ Links ]
28. MORA-OSEJO, L.E. 2004. Respuestas adaptativas de plantas del páramo de bioforma contrastante, frente a las fluctuaciones acentuadas del clima y del foto período. Simposio: Ecología y fisiología de plantas tropicales de alta montaña, pp. 102-114. VIII Congreso latinoamericano y II Colombiano de Botánica. [ Links ]
29. MORA-OSEJO, L.E. & H. STURM. 1995. Estudios ecológicos del páramo y del bosque altoandino cordillera Oriental de Colombia. Tomos I y II. Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Colección Jorge Álvarez Lleras Nº 6. [ Links ]
30. MOORE, T.R. 1986. Carbon dioxide evolution from subartic peatlands in eastern Canada. Arctic Alpine Res. 18: 189-193. [ Links ]
31. MURCIA-RODRÍGUEZ, M.A. 2001. Aislamiento térmico resultante de la bioforma caulirrósula de Espeletia spp en los páramos de Monserrate, Chingaza, Ocetá, Nevados del Tolima y el Ruiz. Tesis de MSc en Biología línea Ecología, Universidad Nacional de Colombia, Departamento de Biología, Bogotá. [ Links ]
32. MURCIA-RODRÍGUEZ, M.A. & L.E. MORA. 2003. Aislamiento térmico del manto de hojas de Espeletiinae de Colombia de bioforma caulirrosular. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales. Vol XXVII, número 105. [ Links ]
33. POOLE, D.K. & P.C. MILLAR. 1982. Carbon dioxide flux from three arctic tundra types in north-central Alaska, U.S.A. Arctic Alpine Res. 14: 27-32. [ Links ]
34. RAICH, J.W. & K.J. NADELHOFFER. 1989. Belowground carbon allocation in forest ecosystems: global trends. Ecology 70: 1346-1354. [ Links ]
35. RAICH, J.W. & W.H. SCHLESINGER. 1992. The global carbon dioxide flux in soil respiration and its relation to vegetation and climate. Tellus. 44B: 81-99. [ Links ]
36. RAICH, J.W. & C.S. POTTER. 1995. Global patterns of carbon dioxide emissions from soil respiration. Global Biochemistry. Cy. 9: 23-36. [ Links ]
37. RAICH, J.W., C.S. POTTER & D. BHAGAWATI. 2002. Interannual variability in global soil respiration. Global Change Biol. 8: 800-812. [ Links ]
38. RAMÍREZ, G., & B. RAMÍREZ. 2006. Colinealidad y mínimos cuadrados ponderados. Revista Venezolana de Análisis de Coyuntura 12(1): 297-299. [ Links ]
39. RANGEL-CH., J.O. 2000. Colombia. Diversidad Biótica III. La región de vida paramuna. Universidad Nacional de Colombia. Facultad de Ciencias. Instituto de Ciencias Naturales, Bogotá, D.C. [ Links ]
40. SÁNCHEZ, M.L.R., S.M. GÉLVIZ & F. SOLANO. 2007. Plantas con Flores de la Reserva El Volcán, Pamplona, Norte de Santander. Bistua 5(2): 27-34. [ Links ]
41. SCHELESINGER, W.H. 1977. Carbon balance in terrestrial detritus. Annu. Rev. Ecol. Syst. 8: 51-81. [ Links ]
42. SCHELESINGER, W.H. & J.A. ANDREWS. 2000. Soil respiration and the global carbon cycle. Biochemistry 48: 7-20. [ Links ]
43. SCHULZE, E.D. 1967. Soil respiration of tropical vegetation types. Ecology 48: 652-653. [ Links ]
44. SINGH, J.S. & S.R. GUPTA. 1977. Plant decomposition and soil respiration in terrestrial ecosystems. Bot. Rev. 43: 449-528. [ Links ]
45. SPSS. Inc. 2006. SPSS 16.0 for Windows Evaluation Version. Release 16.0.0. [ Links ]
46. SUBKE, J.A., I. INGLIMA & M.F. COTRUFO. 2006. Trends and methodological impacts in soil Co2 efflux partitioning: a metanalytical review. Global Change Biol. 12: 921-943. [ Links ]
47. SVENSSON, B.H. 1980. Carbon dioxide and methane fluxes from the ombrotrophic parts of a subarctic mire. Ecological Bulletin 30: 235-250. [ Links ]
48. VEENENDAAL, E.M., O. KOLLE & J. LLOYD. 2004. Seasonal variation in energy fluxes and carbon dioxide exchange for a broad - leaved semi - arid savanna – Mopane woodland- in Southern Africa. Global Change Biol. 10, 318-328. [ Links ]
49. VITTINGHOFF, E., D.V. GLIDDEN, S.C. SHIBOSKI & C.E. MCCULLOCH. 2005. Regression Methods in Biostatistics. Linear, Logistic, Survival, and Repeated Measures Models. Springer Science+Business Media, Inc., New York, USA. [ Links ]
50. WALTER, H. & W. HABER. 1957. Uber die Intensitat der Bodenatmung mit Bemerkungen zu den Lundegardhschen. Werten. Ber. Dtsch. Bot. Ges. 70: 275-282. [ Links ]
51. WALTER, H. 1960. Standortlershe (Phytologie III/1). Stuttgart. Ed. Ulmer Verlag. [ Links ]
52. WITKAMP, M. 1969. Cycles of temperature and carbon dioxide evolution from litter and soil. Ecology 50(5): 922-924. [ Links ]
53. YI, Z., FU, S., YI, W., ZHOU, G., MO, J., ZHANG, D., DING, M., WANG, X. & L. ZHOU. 2007. Partitioning soil respiration of subtropical forests with different successional stages in south China. Forests Ecology and Management 243: 178-186. [ Links ]
54. ZUUR, A.F., E.N. IENO & G.M. SMITH. 2007. Analysing Ecological Data. Statistics for Biology and Health. Springer Science + Business Media, LLC, New York, USA. [ Links ]
Recibido: 02/22/2008
Aceptado: 10/23/2008

