











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkINTRODUCCIÓN
Las macrófitas acuáticas son plantas que pueden, bajo condiciones normales, germinar y crecer con al menos su base en el agua, y son lo suficientemente grandes para ser vistas con el ojo desnudo (Fasset 1940). Bajo este término, entonces, se designan un gran conjunto de plantas pertenecientes las divisiones Cyanobacteria, Chlorophyta, Rhodophyta, Xanthophyta, Bryophyta, Pteridophyta y Spermatophyta (Chambers et al. 2008). Dado este amplio espectro taxonómico y filogenético, estas plantas exhiben gran variabilidad morfológica. No obstante, su dependencia al hábitat acuático ha marcado estrategias de vida convergentes que permiten reconocerlas como un grupo definido y clasificarlas según dichas estrategias. Es así como se han propuesto diferentes sistemas de clasificación ecológica de acuerdo con sus formas de vida y crecimiento (Den Hartog y Segal 1964, Schmidt-Mumm 1998). Los diferentes esquemas propuestos basan su clasificación según la relación de la planta con el sustrato y su localización en la columna de agua (Velásquez 1994, Esteves 1998, Cronk y Fennessy 2001, Rial 2003). En la actualidad, el sistema más utilizado es el propuesto por Sculthorpe (1967) en el que se distinguen cuatro formas de vida: sumergidas, flotantes libres, flotantes arraigadas y emergentes arraigadas, donde los tres primeros grupos son considerados hidrófitos y el último helófito (Velásquez 1994, Schmidt-Mumm 1998, Rial 2003). Si bien, hasta ahora las macrófitas parecen un grupo fácilmente distinguible, es preciso indicar que su delimitación es un tanto compleja (Velásquez 1994, Schmidt-Mumm 1998, Rial 2003). En los sistemas acuáticos pulsátiles, dentro de un ciclo de inundación-sequía, muchas plantas acuáticas pueden expresar variaciones fenotípicas y tolerancia a suelos secos como mecanismos de adaptación a las condiciones de su ambiente, llamadas ecofases. Así, en una misma especie se pueden observar variaciones en tamaño, morfología, fisiología y comportamiento reproductivo, denominadas ecofenos (Rial 2003). En consecuencia, se plantea la necesidad de considerar el ciclo hidrológico en la delimitación de las plantas acuáticas y la evaluación de su riqueza.
Las macrófitas acuáticas representan entre el 1 y el 3 % de todas las plantas conocidas (Cronk y Fennessy 2001, Chambers et al. 2008). Su riqueza global, de acuerdo con Chambers et al. (2008), se estima en 2724 especies de las cuales 110 corresponden a briófitos, 171 a helechos y afines, y 2443 son angiospermas. Si bien gran cantidad de especies tienen amplios ámbitos de distribución, estas plantas alcanzan sus mayores niveles de endemismo y diversidad en los trópicos (Chambers et al. 2008). Su importancia biológica está asociada a su rol en los ecosistemas acuáticos. Son productoras primarias, participan en la formación de detritos, el ciclado de nutrientes y en la fijación de la energía que sustenta todos los componentes de los ecosistemas acuáticos (Cook et al. 1974, Esteves 1998, Cronk y Fennessy 2001, Thomaz et al. 2009). Son determinantes de la diversidad, estructura y dinámica de comunidades animales (Gregg y Rose 1985, Grenouillet et al. 2002, Lansac-Tôha 2003, Thomaz y Cunha 2010). Además, tienen la capacidad de absorber y acumular metales pesados (Miretzky et al. 2004, Peng et al. 2007, Skinner et al. 2007), por lo tanto, son bioindicadoras, biomonitoras y depuradoras del agua. Pese a esta enorme importancia, las macrófitas conforman un grupo biológico poco conocido, tanto a nivel global como local, si se comparan con las plantas terrestres.
La investigación científica acerca de la biología y ecología de los ecosistemas acuáticos continentales tiene una historia relativamente reciente. Si bien, el conocimiento y estudio de las plantas acuáticas inició desde hace varios siglos con Theophrastus (371-287 a. C.), es a partir de la década de los 60 del siglo XX cuando empiezan a generarse una gran cantidad de trabajos que involucran no solo la dinámica de los humedales, sino también la ecología y aspectos taxonómicos de los organismos que habitan dichos ecosistemas, particularmente las macrófitas (Rial 2003, Thomaz y Bini 2003, Padial et al. 2008, Machado-Filho et al. 2014). En las últimas décadas se ha evidenciado un aumento en las publicaciones dedicadas a las plantas acuáticas neotropicales y el conocimiento del grupo es adelantado en países como Brasil (Pott y Pott 2000, Moura-Júnior et al. 2013, Moura-Júnior et al. 2015), México (Lot et al. 2013, Lot 2017), Paraguay (Mereles et al. 2015), Perú (Leon y Young 1996, Kahn et al. 1993) y Venezuela (Velásquez 1994).
En Colombia, el estudio de las macrófitas acuáticas ha tenido creciente interés en los últimos años. La vegetación acuática del país ha sido bien documentada principalmente en las regiones Andina (Rangel-Ch y Aguirre-C 1983, Schmidt-Mumm 1998, Durán-Suárez et al. 2011, Posada y López 2011, Ramos-Montaño et al. 2013, Gómez et al. 2017), Caribe (Rangel-Ch 2010, Cortés-Castillo y Rangel-Ch 2013, Pérez-Vásquez et al. 2015, Cortés-Castillo 2017) y Orinoquía (Fernández et al. 2015, Madriñán et al. 2017, Vera 2017). Aunque se cuenta con un buen número de publicaciones dedicadas al grupo, muy poco se han explorado aspectos relacionados con su taxonomía y fitogeografia. Por ello, se sabe muy poco acerca de su riqueza y patrones de distribución. El reciente catálogo de Plantas y Líquenes de Colombia muestra que en este país las plantas acuáticas están representadas por 168 especies (Bernal et al. 2016). Estos datos lejos de mostrar la realidad de la flora acuática colombiana dan cuenta del escaso conocimiento florístico del grupo. Se hace evidente su bajo nivel de muestreo y poca representatividad en los herbarios nacionales.
La serranía de La Lindosa hace parte de los afloramientos rocosos del escudo Guayanés colombiano (Etter 2001, Córdoba 2014). Se encuentra localizada en la parte norte del municipio de San José del Guaviare (Guaviare, Colombia), en una zona considerada de transición entre las sabanas orinocenses y la selva amazónica. Este macizo rocoso se encuentra bañado por numerosos cuerpos de agua representados por una red de caños y charcos temporales que albergan una importante flora acuática hasta hoy desconocida. En el área se han realizado inventarios florísticos de la vegetación terrestre (López 2005, Cárdenas et al. 2008) y recientemente Madriñán et al. (2017) realizaron muestreos de la vegetación acuática, sin embargo, aún no se ha evaluado la composición de macrófitas de este ecosistema.
El poco conocimiento florístico entorno a las plantas acuáticas en Colombia, dada la tendencia al enfoque ecológico en los trabajos que involucran este grupo de plantas, pone de manifiesto la necesidad de generar estudios básicos, particularmente aquellos que involucran la taxonomía, florística y fitogeografia. En vista de esto, el presente estudio tuvo como objetivo realizar un inventario de las macrófitas acuáticas en los ambientes lóticos y lénticos de la serranía de La Lindosa, y clasificar sus formas de vida de vida. De esta manera, este trabajo contribuirá con el conocimiento de la flora acuática regional y nacional.
MATERIALES Y MÉTODOS
Área de Estudio
La serranía de La Lindosa es una formación rocosa ubicada entre los 02°27'-02°34' Norte y 72°50'-72°36 Oeste. Se encuentra en el municipio de San José del Guaviare, al norte del departamento del Guaviare, en la zona norte de la región amazónica de Colombia. Este macizo pertenece a la Guayana colombiana y dada su afinidad florística con el sur de La Macarena conforman una unidad fitogeográfica: complejo Macarena-Lindosa (Córdoba 2014). En conjunto las áreas del escudo guayanés colombiano hacen parte de la provincia occidental de la región biogeográfica de la Guayana, la cual hace parte del Escudo Guayanés sudamericano (Etter 2001, Córdoba 2014). Esta serranía tiene una extensión aproximada de 12 000 hectáreas, con altitudes entre 225 y 470 m (Cárdenas et al. 2008). Posee clima tropical lluvioso, con precipitación promedio anual de 2800 mm, de régimen monomodal. El periodo lluvioso se presenta entre marzo y octubre, y el periodo seco entre noviembre y febrero; la temperatura promedio es 25,7 °C y la humedad relativa 78-90 % (Cárdenas et al. 2008). Hidrológicamente, los cuerpos de agua en la serranía de La Lindosa hacen parte del sistema de ríos de aguas claras (Lasso 2014), que son afluentes del río Guaviare, el cual pertenece a la cuenca del Orinoco. La vegetación en la zona está conformada por bosques riparios y bosques asociados a grietas de afloramientos rocosos, arbustales sobre afloramientos y arenas blancas, y sabanas de tipo orinocense (López 2005).
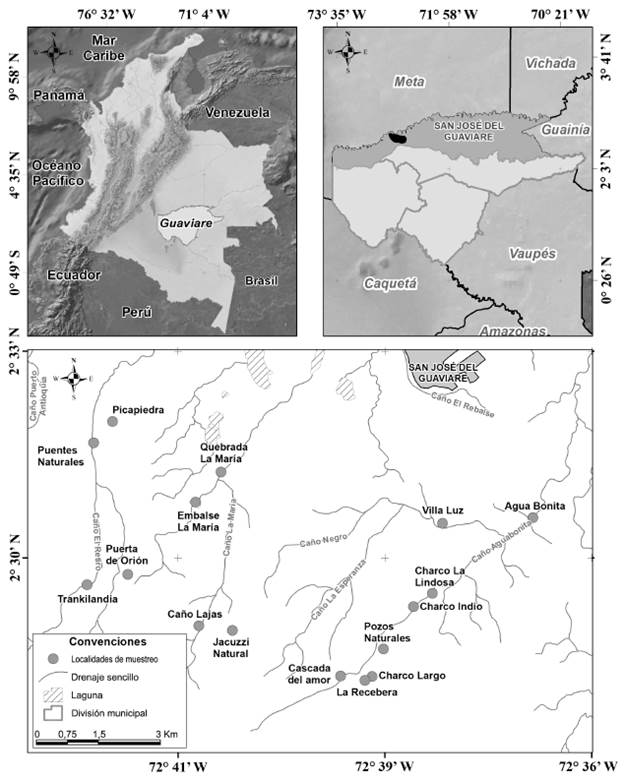
El área de estudio hace parte de las microcuencas de los caños Agua Bonita, La Lindosa y La María, las cuales pertenecen a las veredas Agua Bonita y El Retiro, en el sector nororiental de la serranía de La Lindosa. Se muestrearon los cuerpos de agua lóticos y lénticos de 16 localidades (Fig. 1). Dichas localidades corresponden a los sitios más frecuentemente visitados por la comunidad guaviarense y los turistas, por lo tanto, constituyen las áreas más vulnerables dentro de la serranía dada la intensa actividad humana. De ahí la importancia de evaluar la riqueza de macrófitas del sector, con el fin de obtener información preliminar que pueda servir para el monitoreo y futuras estrategias de conservación del área.
Inventario florístico
Se realizaron tres expediciones de campo durante el año 2016, abarcando los diferentes periodos del ciclo hidrológico: temporada seca (enero), inicio de lluvias (marzo) y máximo nivel de precipitación (julio). Adicionalmente, en el año 2017 se realizaron visitas cortas a las localidades durante los meses del ciclo anual. Los ambientes acuáticos muestreados incluyeron charcos temporales sobre lajas, bajos, embalses y caños. El muestreo se realizó al azar mediante recolecciones generales, procurando abarcar la mayor área posible. En cada localidad se realizaron recorridos a pie y se recolectó material tanto al interior como en ambas márgenes de los cuerpos de agua. De cada especie se registró su forma de vida según el sistema de Sculthorpe (1967): enraizada emergente, enraizada flotante, flotante libre y sumergida.
Las técnicas de recolección y preservación de las muestras variaron de acuerdo con las características de los taxones teniendo en cuenta los lineamientos de Lot y Chiang (1986) y Warrington (1994). Las plantas sumergidas fueron prensadas mediante la técnica de flotación (Taylor 1977, Haynes 1984) y se eliminó el exceso de agua usando papel absorbente. El secado se realizó manualmente ubicando las muestras entre papel periódico y láminas de cartón, y colocando un objeto pesado para ejercer presión sobre los especímenes. Se permitió la exposición al sol y viento para favorecer el rápido secado del material, además, se cambió el periódico diariamente para evitar el crecimiento de hongos. Finalmente, el material colectado fue procesado y depositado en el Herbario Nacional Colombiano (COL). La determinación de los ejemplares se realizó mediante revisión de literatura botánica especializada: Fernández-Pérez (1964), Velásquez (1994), Berry et al. (1995), Churchill y Linares (1995), Berry et al. (1997), Berry et al. (1998), Berry et al. (1999), Berry et al. (2001), Gradstein et al. (2001), Berry et al. (2003), Berry et al. (2004), Berry et al. (2005), Bernal et al. (2016). Además, las determinaciones fueron confirmadas mediante comparación con ejemplares de referencia de COL y consulta con especialistas.
RESULTADOS Y DISCUSIÓN
Riqueza florística
En los ambientes acuáticos de la serranía de La Lindosa se encontraron 110 especies pertenecientes a 70 géneros y 46 familias (Tabla 1). Rial (2014) estima la riqueza de plantas acuáticas de la cuenca del Orinoco en 300 a 400 especies. Fernández et al. (2015) y Madriñán et al. (2017) por su parte reportan para la porción colombiana de esta cuenca 341 especies en 221 géneros y 95 familias. De esta manera, en La Lindosa se encontraría el 32 % de las macrófitas de la Orinoquia colombiana y alrededor del 28 % para toda la cuenca. La velocidad del agua es un importante determinante de la riqueza y composición de plantas acuáticas. De acuerdo con Rial (2014) la velocidad de la corriente limita la presencia de muchas especies, de manera que la riqueza es mayor en ambientes lénticos que en los lóticos. Así pues, los valores de riqueza obtenidos para el área de estudio son considerables si tenemos en cuenta su pequeña extensión estimada en 12 000 hectáreas (Cárdenas et al. 2008) y la ausencia de lagos y lagunas, ambientes lénticos donde proliferan más las plantas acuáticas. A esta riqueza entonces contribuyen la presencia de numerosos charcos temporales y áreas de los caños donde se interrumpe su cauce debido a la presencia de árboles caídos o represas construidas para fines turísticos.
Tabla 1 Riqueza florística, bioforma y ejemplar testigo de las plantas acuáticas de La serranía de La Lindosa. * Nuevo registro para el Guaviare, ** nuevo registro para Colombia. Su = Sumergida, AF = Arraigada flotante, AE = Arraigada emergente.

Las plantas no vasculares estuvieron representadas por quince especies de las cuales cinco fueron hepáticas y diez musgos (Tabla 1). Estos valores son considerables si tenemos en cuenta que para el departamento del Guaviare se registran solo tres especies de hepáticas y 21 de musgos (Alvaro-Alba et al. 2011). Además, se declara la importancia de realizar muestreos en ambientes acuáticos dada su pobre exploración. En cuanto a la riqueza a nivel de familias se encontró que Sematophyllaceae, con cuatro especies en tres géneros, fue la más importante. Esto no es sorprendente pues dicha familia es una de las más ricas en especies en el neotrópico, particularmente en tierras bajas (Gradstein et al. 2001).
Las plantas vasculares incluyeron 96 especies las cuales estuvieron representadas por seis pteridófitos y 90 angiospermas (Tabla 1). Las familias con mayor riqueza fueron Cyperaceae (seis géneros/17 especies), Poaceae (diez/ catorce), Lentibulariaceae (uno/catorce) y Eriocaulaceae (tres/seis). En conjunto estas cuatro familias contienen el 46 % de las especies de la flora acuática de la serranía de La Lindosa. La mayor riqueza de las familias Cyperaceae y Poaceae es consistente con lo presentado por Fernández et al. (2015) y Rial (2014) para los llanos colombo-venezolanos. Además, estas familias típicamente dominan los ambientes acuáticos de Suramérica, incluyendo cuencas hidrográficas, lagos, lagunas, humedales y ciénagas (Leon y Young 1996, Matias et al. 2003, Terneus 2007, Henry-Silva et al. 2010, Moura-Júnior et al. 2013). Esto constituye una tendencia no solo en el neotrópico (Machado-Filho et al. 2014) sino para todas las regiones biogeográficas del mundo con excepción de la Antártida (Chambers et al. 2008). Tal éxito en estas familias puede ser atribuido a su plasticidad ecológica. La familia Cyperaceae está constituida por cerca de 5000 especies (Goetghebeur 1998), mientras que Poaceae tiene alrededor de 10 000 (Giraldo-Cañas 2010). Para la Guayana colombiana esta última familia se encuentra representada por el 18,7 % de las especies y el 43,7 % de los géneros de gramíneas del país (Giraldo-Cañas 2010) mostrándonos la alta riqueza en la región. La plasticidad ecológica explica la gran riqueza de estas familias en ambientes acuáticos asociada a la eficiencia en los mecanismos de reproducción vegetativa por medio de sistemas subterráneos formados por rizomas y estolones (Goetghebeur 1998, Matias et al. 2003, Moura-Júnior et al. 2013). Además, su naturaleza perenne, tolerancia al periodo de sequía (Bove et al. 2003) y mecanismo de dispersión anemófila les permiten colonizar con éxito numerosos ambientes, tanto terrestres como acuáticos.
Los géneros más representativos fueron Utricularia con catorce especies y Eleocharis con seis, seguidos por Cyperus, Rhynchospora, Syngonanthus y Xyris con cuatro especies cada uno; además, 54 géneros estuvieron representados por una sola especie (Tabla 1). La mayor riqueza del género Utricularia es similar a la presentada por Fernández et al. (2015) para toda la región Orinoquia de Colombia y estaría asociada a su mecanismo de nutrición carnívora que le ha permitido diversificarse en zonas con baja disponibilidad de nutrientes como es el caso de los afloramientos rocosos guayaneses. Asimismo, hay que considerar que dicho género tiene centros de diversificación en Suramérica, en regiones de Brasil, Venezuela y Guayana (Fischer et al. 2004). De acuerdo con los datos del Sistema de Información sobre Biodiversidad de Colombia (SIB Colombia c2018), en el país las Utricularias se encuentran representadas por 32 especies. Así pues, La Lindosa contiene alrededor del 44 % de las especies de este género. Este dato es bastante sorprendente pues con una mejor exploración de la zona encaminada a conocer la riqueza del grupo seguramente el número de especies aumentará. De esta manera, este macizo constituye un importante reservorio de lentibulariáceas en Colombia.
La mayor cantidad de géneros o familias representas por una única especie en algunos casos puede estar relacionada con el hábitat predominante de la familia. Por ejemplo, las Bromeliaceae son típicamente terrestres, epífitas o litófitas (Berry et al. 1997), Amaranthaceae predomina en zonas cálidas y secas (Agudelo-H y Rosselli 1991), mientras que las especies de Burmanniaceae generalmente son saprofitas (Maas-Van de Kamer 1998). Por lo tanto, encontraremos muy pocas especies de dichas familias en ambientes acuáticos. En contraposición, las familias cuyos géneros presentaron mayor riqueza, Lentibulariaceae (Utricularia) y Cyperaceae (Eleocharis, Cyperus, Rhynchospora), contienen gran cantidad de especies acuáticas o de hábitats húmedos (Berry et al. 1998). Las familias Eriocaulaceae y Xyridaceae son típicas de aguas negras de la región guayanesa en Colombia y Venezuela, donde algunas de sus especies caracterizan fisionómicamente ciertos ambientes (Rial 2014). Lo anterior puede explicar la alta riqueza de especies de Syngonanthus (Eriocaulaceae) y Xyris (Xyridaceae).
Se reporta un nuevo registro para la flora de Colombia, Eleocharis alveolatoides S. González & Reznicek, y 38 para el departamento del Guaviare (Tabla 1). Los datos muestran además que el 35 % de las plantas acuáticas encontradas en este estudio eran desconocidas para la región. Este alto porcentaje de nuevos registros demuestra el poco conocimiento que se tiene de las plantas acuáticas debido a la escasez de muestreo en la zona. Lo anterior es más evidente para el caso de los briofitos pues once de los quince especímenes encontrados constituyen nuevos registros.
Los humedales de la región guayanesa se caracterizan por presentar suelos arenosos y rocosos de historia geológica antigua, donde las aguas son trasparentes y oligotróficas (Rial 2014). Estas características han favorecido la expresión de una flora singular, con elementos de distribución restringida. Los ambientes acuáticos incluyen áreas de morichales, sabanas inundables donde habitan especies como Drosera cayennensis y Utricularia simulans, lajas rocosas en ríos de aguas negras donde son frecuentes Abolboda pulchella y Portulaca pusilla, raudales, saltos y zonas torrentosas donde son comunes las especies de podostemáceas (Rial 2014). En la serranía de La Lindosa se destaca la presencia de especies típicas de esta región guayanesa y de distribución geográfica restringida o reducida: Rhyncholacis clavigera (=Macarenia clavigera) en serranías La Macarena y Lindosa (Colombia), Utricularia schultesii Este de Colombia y Oeste de Venezuela, Eleocharis alveolatoides serranía La Lindosa en Colombia, afloramientos en Bolívar (Venezuela) y Roraima (Brasil), y Utricularia chiribiquetensis sudeste de Colombia, sudoeste de Venezuela y noroeste de Brasil. De manera que se muestra la importancia de monitorear y generar adecuadas medidas de preservación de este afloramiento rocoso.
Formas de vida
La forma de vida arraigada emergente fue la más abundante con 82 especies, le siguieron las plantas sumergidas con 20 y por último las arraigadas flotantes con tres (Tabla 1). Además, seis especies manifestaron ecofases pudiendo encontrarse sumergidas al interior de los cuerpos de agua o emergentes en las orillas: Calyptrocarya glomerulata, Eleocharis alveolatoides, Eriocaulon spruceanum, Syngonanthus humboldtii, Tonina fluviatilis y Mayaca sellowiana.
En los ambientes acuáticos las áreas de ribera contienen la mayor diversidad de macrófitas. Sus representantes dominantes son las plantas helófitas, las cuales deben su éxito a la capacidad de tolerar la inundación y la sequía, manteniendo una vida anfibia. De acuerdo con Cronk y Fennessy (2001) este tipo de plantas logran dominar el hábitat acuático debido a su habilidad para interceptar la luz solar antes que esta alcance la superficie del agua. La mayor proporción de la bioforma arraigada emergente en las plantas acuáticas de La Lindosa es consistente con lo encontrado en otros estudios (Leon y Young 1996, Matias et al. 2003, Terneus 2007, Henry-Silva et al. 2010, Moura-Júnior et al. 2013) y representa una tendencia en el neotrópico (Machado-Filho et al. 2014). La gran riqueza de las familias Cyperaceae y Poaceae contribuye con el predominio de la forma de vida arraigada emergente. En estos grupos, sus complejos sistemas subterráneos cumplen una importante función de estabilización de sedimentos en las márgenes de los cuerpos de agua (Matias et al. 2003), además, su ajuste morfológico y fisiológico les permite vivir entre el ambiente acuático y el terrestre (Moura-Júnior et al. 2013).
Los hidrófitos se encuentran a lo largo de todo el gradiente de profundidad, desde las orillas hasta unos pocos metros de profundidad, en aguas cristalinas. Siempre crecen hacia el interior del espejo de agua y bajo condiciones favorables logran colonizar rápidamente su hábitat. La riqueza de plantas sumergidas estaría asociada a las características de los ambientes acuáticos de la serranía de La Lindosa. Los caños son de aguas cristalinas poco profundas con suelos rocosos y en muchas áreas se presentan rápidos y caídas de agua donde proliferan especies reófitas como Rhyncholacis clavigera y Utricularia neottioides.
Así mismo, destacan las especies de briófitos sumergidos, muchos de los cuales constituyen elementos propios de las regiones Guayana y Amazonia como Hydropogon fontinaloides, Potamium lonchophyllum y Sphagnum negrense (Churchill y Linares 1995, Gradstein et al. 2001).
Finalmente, la plasticidad ecológica de las especies de plantas acuáticas refleja su capacidad de ajuste morfo-fisiológico (Matias et al. 2003, Moura-Júnior et al. 2013). Dicha plasticidad se manifiesta como ecofases que incluyen dos estrategias principales: 1) modificaciones anatómicas, fisiológicas y morfológicas para permanecer en el mismo ambiente durante los períodos de lluvia y sequía (p. ej. Sagittaria guayanensis), 2) capacidad de adoptar más de una bioforma durante su ciclo de vida (p. ej. Calyptrocarya glomerulata). Estas estrategias adaptativas hacen de estas especies componentes abundantes en las áreas de humedal de La Lindosa.


