











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCCIÓN
Las abejas son insectos esenciales para procesos de polinización tanto en ambientes naturales como en sistemas agrícolas y contribuyen al mantenimiento de servicios ecológicos y resiliencia ecológica en estos paisajes fragmentados (Decourtye et al., 2009). De esta manera, las abejas juegan un rol fundamental para la productividad alimentaria, puesto que cerca de un 80 % de las plantas de interés agrícola son polinizadas por estos insectos (Klein et al., 2007). De acuerdo con Kleijn et al. (2015), las abejas silvestres aportan cerca de USD $3251/ha anuales a la producción de cultivos, lo que demuestra la importancia de la fauna silvestre como mediadores de servicios ecosistémicos (Kleijn et al., 2015). Asimismo, los sistemas productivos junto con las áreas naturales son importantes para las abejas, porque otorgan una diversidad de recursos que pueden suplir sus necesidades (Plaza-Ortega et al., 2017).
En estudios orientados a entender la dinámica y salud de los ecosistemas naturales y su interacción con sistemas intervenidos, es importante entender el aporte de la diversidad taxonómica y funcional en los procesos del ecosistema. En Colombia se tienen registros de aproximadamente 600 especies que se han evaluado a partir de parámetros como la riqueza o número de especies, la composición o especies que conforman un ensamblaje y la estructura o valor de abundancia de las especies, tanto en sistemas naturales como agrícolas (Nates-Parra, 2006; Medina-Gutiérrez et al., 2012; Nates-Parra, 2016; Sepúlveda-Cano et al., 2017; Gutiérrez-Chacón et al., 2018). Sin embargo, para los agroecosistemas en Colombia la información de diversidad taxonómica de abejas es poca y se encuentra dispersa, por lo cual no se han identificado patrones generales de esta dimensión de la diversidad biológica.
La dimensión funcional de abejas ha sido abordada de manera global, desde la perspectiva del papel ecológico, con varios estudios sobre polinización en cultivos (Forrest et al., 2015) y sobre los efectos de ambientes urbanos sobre la diversidad de abejas (Buchholz et al., 2020). No obstante, son pocos los estudios que abordan la funcionalidad desde las métricas de la diversidad de rasgos funcionales y cómo estos ayudan al desempeño de las abejas para cumplir con sus roles ecológicos. La diversidad funcional de abejas, entendida como el valor, rango, distribución y abundancia relativa de los caracteres funcionales de los organismos en un ecosistema (Díaz et al., 2007), juega un papel importante en la dinámica de los sistemas intervenidos. De esta manera se ha identificado que los rasgos funcionales aumentan el funcionamiento de los agroecosistemas en todo el mundo (Garibaldi et al., 2011). Las abejas presentan características morfológicas y etológicas, como la presencia de corbícula, el grado de sociabilidad y la anidación, que les permite llevar a cabo sus funciones ecológicas (Ollerton, 2017) y favorecer la producción de los sistemas agrícolas, de países como Colombia, cuyo PIB depende en gran parte de esta (FAO, 2008).
Colombia cuenta con grandes extensiones de tierra dispuesta para la siembra de cultivos estructuralmente simples, como los de papa en ecosistemas de alta montaña, hasta más complejos como los de café en la media montaña y palma y maracuyá en las tierras bajas (FAO, 2016). Sin embargo, pese a la alta representatividad de los sistemas agrícolas en el país, no se cuenta con información que documente el aporte del componente agrícola al mantenimiento de la diversidad taxonómica y funcional de abejas. Por esta razón, se requiere tener inventarios unificados del conocimiento de la riqueza, composición y abundancia y representatividad de rasgos funcionales de abejas en sistemas productivos del país.
Esta investigación tiene como objetivo hacer una revisión de los estudios que aborden la diversidad taxonómica y funcional de abejas en sistemas agrícolas característicos de Colombia. Se buscó compilar el inventario de las abejas asociadas a cultivos y determinar la representatividad de las dimensiones taxonómica y funcional de las abejas en estos sistemas. Se partió de la hipótesis de que los cultivos de papa por presentar una estructura vegetal simple, presentan valores bajos de diversidad taxonómica y funcional, contrario a aquellos sistemas productivos cuya estructura es más compleja. Para contribuir al mejoramiento del estado actual del conocimiento de la diversidad taxonómica y funcional de abejas en sistemas productivos agrícolas se plantean perspectivas de investigación que se enfoquen en determinar la importancia de hábitats semitransformados (interacción cultivo-vegetación natural) sobre el comportamiento de la diversidad de abejas.
MATERIALES Y MÉTODOS
Sistemas productivos escogidos
Se seleccionaron seis cultivos asociados a diferentes zonas de vida a lo largo del gradiente altitudinal de los Andes colombianos. Estos fueron el cultivo de palma (Elaeis guineensis Jacq; Arecaceae), maracuyá (Passiflora edulis Sims; Passifloraceae), café (Coffea arabica Chev; Rubiaceae), granadilla (Passiflora ligularis Juss; Passifloraceae), gulupa (Passiflora pinnatistipula Cav; Passifloraceae) y papa (Solanum tuberosum Linneo; Solanaceae) (FAO, 2016). Estos sistemas productivos son representativos de la economía colombiana y se ha documentado que albergan una alta diversidad de abejas (Nates-Parra, 2016).
Revisión y búsqueda de la información
Se realizó la búsqueda de información entre febrero y agosto de 2020, de abejas asociadas a cultivos de interés comercial en Colombia. Se utilizaron bases de datos como Google Scholar, ScienceDirect, EBSCOhost, Web of Science y Scopus. Se buscaron artículos mediante combinaciones de palabras clave en inglés como: "Bees", "Apoidea", "Crops", "Coffe" "Palm", "Potato", "Passion fruit", "passionflower", "Gulupa", "Agroecology", "Pollination", "paramos", "lowland", "tropical zone" y "Colombia" y sus equivalentes en castellano; para ambos casos se utilizaron operadores boo-leanos como AND, NOT, OR (Dodge, 2005). Cada artículo se filtró de acuerdo al tipo de cultivo, año de publicación y área de estudio (zona de vida) en el que se llevó a cabo (Tabla 1). Además, solo se tuvieron en cuenta aquellos artículos que evaluaron exclusivamente la caracterización de abejas asociadas a los cultivos escogidos independientemente del año de publicación.
Análisis de datos
Se realizó una matriz con datos de presencia/ausencia en la cual se registraron las especies de abejas asociadas a cada cultivo. Se excluyeron los registros de morfoespecies presentes en dos o más cultivos, para evitar sobreestimar la riqueza de especies. La diversidad taxonómica de abejas se tomó como la riqueza de especies para cada cultivo. Se construyeron curvas análogas a la acumulación de especies, por periodos de publicación de los artículos, mediante el estimador Jackniffe 2, que permite realizar análisis de datos de incidencia y logró esquematizar de mejor manera la acumulación de la riqueza por cada estudio adelantado. El análisis se desarrolló en programa EstimateS versión 9.1. Adicionalmente se realizó un análisis de complementariedad para evaluar el grado de disimilitud en la composición de especies entre cultivos (Moreno, 2001).
Para los análisis de diversidad funcional, se utilizaron tres rasgos funcionales, uno morfológico (presencia/ausencia de corbícula), y dos etológicos (grado de sociabilidad y sitio de anidación) (Tabla 2). Se utilizó un análisis de conglomerados para identificar tipos funcionales mediante el método de Ward y distancias euclidianas, en el programa INFOSTAT (Di Rienzo et al., 2008). Adicionalmente, se utilizó el índice de riqueza funcional (FRic) (Villéger et al., 2008), para evaluar la representatividad de la diversidad funcional en los cultivos. Se excluyó de los análisis funcionales el cultivo de maracuyá, por su baja representatividad en la muestra. Los análisis se llevaron a cabo en programa FDiversity con extensión a R con una interfaz escrita en Delphi® y DCOM-R.

RESULTADOS
Aspectos generales
Se encontraron 19 publicaciones con inventario de abejas asociadas a los seis cultivos escogidos (Tabla 1). Para maracuyá se encontraron cinco estudios realizados en las zonas de vida Subandina y Andina, entre los 1300 y 2200 m.s.n.m. (Calle et al., 2010; Arias-Suárez et al., 2014; González y González, 2015; Rodríguez-Calderón, 2016; Bezerra et al., 2019). Para el cultivo de palma se registraron cuatro estudios en la zona de vida tropical de la región amazónica (Guerrero-Olaya y Núñez, 2017; Núñez y Carreño, 2017), de la Orinoquia, el Pacífico (Núñez y Carreño, 2016) y de la región Caribe (Brieva-Oviedo y Núñez, 2020). Para abejas asociadas a cultivos de papa, se encontraron tres estudios de entre los 2400 y 2700 m.s.n.m. en región Andina (Barrientos-Restrepo, 2012; Sepúlveda-Cano, 2013; Sepúlveda-Cano et al., 2017). Para el cultivo de café se registraron tres estudios en la zona de vida Subandina entre los 1200 y 1900 m.s.n.m. (Jaramillo-Delgado, 2012; Bravo-Monroy et al., 2015; Herrera y Sabogal, 2016). Se encontraron tres estudios enfocados a los cultivos de granadilla en las regiones de vida Subandina y Andina entre los 1800 y los 2300 m.s.n.m. (Franco et al., 2007; Suárez et al., 2016; Gutiérrez-Chacón et al., 2018) y para abejas asociadas a cultivos de gulupa, se encontró un estudio realizado en la región Subandina y Andina, entre los 1600 y 2200 m.s.n.m. (Medina-Gutiérrez et al., 2012).
Representatividad de la diversidad taxonómica
Se obtuvo un inventario de 116 especies, representadas en 49 géneros, 16 tribus y cinco familias. La riqueza más alta se registró en el cultivo de palma con 48 especies, 15 géneros y dos tribus; seguido del cultivo de papa con 44 especies, 31 géneros y 14 tribus y café con 41 especies, 25 géneros y diez tribus. En los cultivos de granadilla y gulupa se registraron 16 y 11 especies respectivamente y en el cultivo de maracuyá se registró una especie, Xylocopa sp. (Anexo 1). De acuerdo al estimador de diversidad jackknife 2, los cultivos de palma y granadilla presentan mayor potencial para albergar más especies de abejas (Fig. 1). Para los cultivos de papa, café y gulupa la curva fue asintótica.
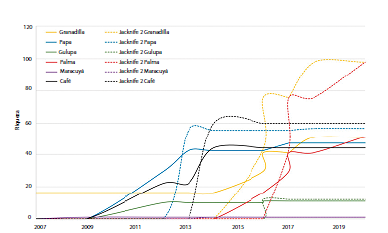
Figura 1 Riqueza de especies de abejas en cada cultivo. En línea continua las especies registradas. En línea discontinua estimadores de diversidad.
La disimilitud en la composición de especies entre los cultivos fue alta y osciló entre 1 para los cultivos de palma y maracuyá y 0,77 entre la gulupa y la granadilla. Los cultivos con más especies compartidas fueron café y papa con 15, seguido de café y palma con 11 y granadilla y papa con ocho. Se registraron cinco especies compartidas entre los cultivos de gulupa y papa, gulupa y granadilla y gulupa y café (Anexo 1). Finalmente, el cultivo de maracuyá presentó una especie compartida con todos los cultivos (Xylocopa sp.), excepto con el cultivo de palma. Los cultivos de palma, papa y café, presentaron la mayor cantidad de especies exclusivas con 36, 24 y 16 especies respectivamente y los cultivos de granadilla y gulupa presentaron seis y tres especies exclusivas.
Representatividad de la Diversidad Funcional
Las abejas se clasificaron en 11 tipos funcionales (Fig. 2). El grupo con presencia de corbícula, sociales y que anidan en cualquier cavidad (ScEuCc), agrupó el 43,9 % de especies. Las abejas sin corbícula, solitarias y que anidan en el suelo (Nc-SoSu) y las abejas con corbícula, solitarias y que anidan en cualquier cavidad (ScSoCc) representaron el 18,1 % y 7,75 % respectivamente. Finalmente, los tipos funcionales menos representados fueron las abejas sin corbícula, solitarias y que anidan en madera (NcSoMa) representados por Xylocopa sp., y abejas con corbícula, sociales y cleptoparásitas (ScEuCl) con Exaerete smaragdina como su única especie (Anexo 1, Fig. 2).
Los cultivos de papa y café presentaron la mayor cantidad de tipos funcionales (diez cada uno), mientras que los cultivos de granadilla y gulupa, estuvieron representados por siete tipos. El cultivo de palma presentó tres tipos funcionales (NcSoCc, NcSoSu y ScEuCc) y el cultivo de maracuyá estuvo representado por un tipo funcional (NcSoMa) (Fig. 3). A nivel de riqueza funcional (FRic), se encontró una alta riqueza, congruente con la representatividad de los tipos funcionales; en los cultivos de café (FRic =3,33), papa (FRic = 2,83) y gulupa (FRic = 2,00), mientras que la granadilla y la palma presentaron valores más bajos FRic = 1,33 y FRic = 1,12 respectivamente.
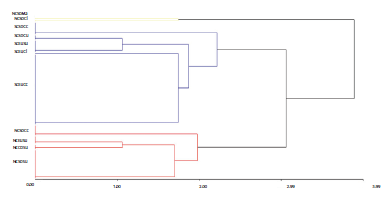
Figura 2 Dendrograma del análisis de conglomerados generado con el método de Ward y utilizando distancias euclidianas para tres rasgos funcionales de abejas asociadas a cultivos en Colombia. Se forman once tipos funcionales: NoCorbícula-So-litaria-Madera (NcSoMa), NoCorbícula-Solitaria-Cleptoparásita (NcSoCl), SiCorbícula-Solitaria-CualquierCavidad (ScSoCc), Si-Corbícula-Comunal-Suelo (ScCoSu), SiCorbícula-Eusocial-Suelo (ScEuSu), SiCorbícula-Eusocial-Cleptoparásita (ScEuCl), SiCor-bícula-Eusocial-CualquierCavidad (ScEuCc), NoCorbícula-Solitaria-CualquierCavidad (NcSoCc), NoCorbícula-Eusocial-Suelo (NcEuSu), NoCorbícula-Comunal-Suelo (NcCoSu) y NoCorbícu-la-Solitaria-Suelo (NcSoSu).
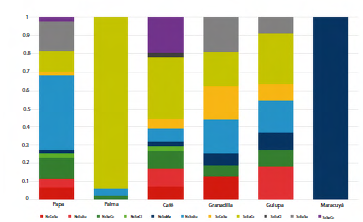
Figura 3 Conjunto de abejas en cultivos de interés en Colombia. Proporción de especies en cada tipo funcional asociado a cada cultivo. NcCoSu: NoCorbícula-Comunal-Suelo; NcEuSu: NoCorbícula-Eusocial-Suelo; NcSoCc: NoCorbícula-Solitaria-CualquierCavidad; NcSoCl: NoCorbícula-Solitaria-Cleptoparásita; NcSoMa: NoCorbícu-la-Solitaria-Madera; NcSoSu: NoCorbícula-Solitaria-Suelo; ScCoSu: SiCorbícula-Comunal-Suelo; ScEuCc: SiCorbícula-Eusocial-Cualquier-Cavidad: SiCorbícula-Eusocial-Cleptoparásita; ScEuSu: SiCorbícula-Eusocial-Suelo; ScSoCc: SiCorbícula-Solitaria-Cualquier Cavidad.
DISCUSIÓN
El bajo número de publicaciones encontradas sobre abejas asociadas a los cultivos evaluados, demuestra el vacío de información sobre la importancia de las dimensiones funcionales y taxonómicas de la diversidad de estos insectos en la producción agrícola de Colombia. Recientemente se han promovido iniciativas importantes sobre la relevancia de las abejas en cultivos agrícolas (Nates-Parra, 2016), sin embargo, la cantidad de información aún es escaza si se tiene en cuenta las grandes extensiones de tierra usadas para esta actividad en el país.
En términos generales, la diversidad de abejas en los sistemas agrícolas parece estar más relacionada a la cercanía de los cultivos con sistemas naturales, que a la complejidad estructural de los cultivos. De acuerdo con los artículos evaluados, muchos cultivos se encontraban cercanos a zonas boscosas o a fragmentos de vegetación, e incluso muchos de ellos estaban inmersos en matrices de vegetación natural (Barrientos-Restrepo, 2012; Jaramillo-Delgado, 2012; Sepúlveda-Cano, 2013; Bravo-Monroy et al., 2015; Herrera y Sabogal, 2016; Guerrero-Olaya y Núñez, 2017; Núñez y Carreño, 2017; Sepúlveda-Cano et al., 2017).
En este estudio se destaca que los cultivos de palma, papa y café hacen un aporte significativo al mantenimiento de la diversidad de abejas silvestres, por lo menos de manera preliminar; mientras que los cultivos de Passifloraceae (granadilla, gulupa y maracuyá), aportan el 27 % de la riqueza de abejas registradas. Sin embargo, la cantidad de estudios y la intensidad de los muestreos es un aspecto clave para dilucidar correctamente los patrones de diversidad. Adicionalmente, es un hecho que las flores de especies de Passifloraceae necesitan de insectos grandes para efectuar su polinización, como saltamontes (Tettigonidae), polillas y abejas de gran tamaño como las del género Xylocopa (Akamine y Girolami 1959), dada la incapacidad de las abejas pequeñas para acceder al néctar por la profundidad de los nectarios.
Representatividad taxonómica
En este estudio se encontraron 116 especies que corresponden al 19,33 % de las abejas registradas para Colombia (Nates-Parra, 2006). Según Ellis y Barbercheck (2015) y Núñez y Carreño (2016), los cultivos agrícolas ofrecen recursos alimentarios y de hábitat a las abejas que les permite cubrir sus requerimientos básicos, además de presentar un gran potencial como herramienta de conservación agrícola. La alta riqueza registrada podría deberse a la heterogeneidad estructural que ofrecen los cultivos y a la variabilidad de formas que presentan sus flores, así como también a la disponibilidad de néctar y polen (Leal et al., 2018). En los cultivos de palma se registró la mayor riqueza de abejas (48 especies), de acuerdo con Núñez y Carreño (2013), las inflorescencias de la palma son un recurso importante para las abejas silvestres debido a la gran cantidad de polen que producen. Sin embargo, la visita constante de abejas y el acceso a la inflorescencia de las palmas, no implica necesariamente que haya polinización ya que no todas las palmas presentan melitofilia (Núñez y Carreño, 2016).
Los cultivos de papa también presentaron una alta riqueza de abejas (44 especies), de las cuales se ha documentado que no todas llevan a cabo una polinización efectiva (De Luca y Vallejo, 2013). De acuerdo con Buchmann (1983), las solanáceas presentan polinización efectiva a través de un método conocido como "Buzz polination" o polinización por vibración, efectuada principalmente por abejas grandes de los géneros Bombus y Xylocopa. Sin embargo, estas plantas pueden ser visitadas por otras abejas (Halictidae, Euglossini, Meliponini), que aprovechan el recurso, pero no polinizan la planta, por lo que actúan como como robadoras de néctar y afectan la relación mutualista planta-polinizador (Inouye, 1980). A pesar de esto, en Colombia la propagación de cultivos de papa se hace de forma vegetativa (a través de tubérculos), por lo que la producción de cultivos no se vería afectada por la visita ilegítima de otras especies. Por lo tanto, los cultivos de papa proveen un recurso importante a las abejas de los ecosistemas circundantes sin verse afectado por la no polinización. Sin embargo, es importante dilucidar la función ecológica de tantas especies en un cultivo donde no realizan su mayor atributo ecológico, la polinización.
La alta riqueza de abejas en cultivos de café (41 especies), permite resaltar su importancia en la conservación. De acuerdo con Jaramillo-Delgado (2012), los cultivos de café permanecen en floración (con diferente intensidad) durante todo el año, por lo que ofrecen un recurso alimentario continuo. Además, algunas variedades de café se establecen generalmente bajo sombrío, por lo que ofrecen mayor disponibilidad de hábitat para las abejas y una mayor interacción con los cultivos naturales (Klein et al., 2007). Cabe destacar que los cultivos de café también se benefician con la alta diversidad de abejas, ya que la polinización del café mediada por estos insectos mejora la producción hasta en un 15 % (Jaramillo-Delgado, 2012).
La alta disimilitud registrada puede estar dada por las condiciones ambientales en las que se establecen cada tipo de cultivo en el gradiente altitudinal de los Andes. Estas condiciones ambientales pueden actuar como importantes filtros que moldean los patrones de riqueza y composición de abejas (Hoiss et al., 2012). Otro factor importante en esta alta disimilitud, puede estar relacionada con los esfuerzos de muestreo que se han adelantado en cada cultivo.
Grupos funcionales de abejas en cultivos
La mayoría de las abejas registradas en estos cultivos (43,9 %) pertenecieron al tipo funcional ScEuCc, que se encontró en todos los cultivos (excepto en maracuyá). Las especies de abejas pertenecientes a este grupo podrían beneficiar los cultivos que visitan dado que la presencia de corbícula permite a las abejas almacenar mayor cantidad de polen (Nates-Parra, 2006) y de igual manera su comportamiento eusocial se ha relacionado con procesos de polinización más eficientes (Kaluza et al., 2017). Los cultivos evaluados no solo albergan una riqueza representativa de abejas en Colombia, sino que, además pueden ser un recurso necesario para el mantenimiento de múltiples poblaciones, a lo que algunos autores han denominado un proceso de codependencia abeja-cultivo (Janzen, 1985). Un ejemplo de esta situación lo representa el cultivo de maracuyá y su único visitante floral (Xylocopa sp.), del grupo ecológico NcSoMa. La declinación poblacional de Xylocopa sp. y por tanto de sus rasgos funcionales que intervienen en el proceso ecológico, podría traer pérdidas en la producción y aumento de costos en los cultivos de maracuyá (Rodríguez-Calderón, 2016). Esto también lo registraron Bezerra et al. (2009) en la Amazonia Brasilera y Da Silva et al. (2012) en México, quienes plantean un escenario donde la no co-ocurrencia entre Xilocopa sp. y cultivos de maracuyá, aumentará los costos de producción por el pago de polinización manual en este cultivo.
Se identificaron 11 grupos ecológicos, varios de ellos representados por más de 20 especies de abejas, lo que podría sugerir una posible redundancia de rasgos funcionales por tipo de cultivo. Sin embargo, dada la complejidad del término redundancia funcional (Ricotta et al., 2016), sería importante evaluar en futuras investigaciones si existe una verdadera redundancia funcional en el sentido que exponen Cadotte et al. (2011) y Casanoves et al. (2011), para los cultivos donde se registraron tipos funcionales con más de 20 especies (ScEuCc y NcSoSu). La presencia de grupos ecológicos de abejas cleptoparásitas (NcSoCl y ScEuCl) en cultivos de café y papa confirma que la comunidad de abejas es estable en estos sistemas agrícolas, ya que las abejas cleptoparásitas utilizan los recursos alimenticios de otras especies (Banaszak y Twerd, 2018).
Los cultivos evaluados mantienen una buena dinámica funcional de abejas, pese a que se ha identificado que la intensificación del uso del suelo aumenta la vulnerabilidad de los grupos funcionales (Laliberté et al., 2010). Forrest et al. (2015) registraron que los sistemas agrícolas atraen a un importante número de abejas, pero funcionalmente similares entre sí, como se evidenció en cultivos de palma (FRic = 1,12), en el que se evidencia una baja congruencia entre las dimensiones taxonómica y funcional. Es por esto que, aunque los cultivos proporcionen un refugio importante para las abejas (Nicholls y Altieri, 2013), se debe profundizar en la conservación de hábitats naturales que amplíen el rango de las funciones ecológicas (Patel et al., 2020).
Perspectivas de investigación
A pesar de que los cultivos evaluados en esta revisión mostraron una alta riqueza de abejas, es posible que puedan albergar una mayor cantidad de especies (Fig. 1). Esto corrobora los resultados expuestos por Rao y Stephen (2010), donde los cultivos agrícolas aumentaron la diversidad de abejas nativas (género Bombus) en los Estados Unidos. Por esto, es importante incrementar los estudios que permitan conocer una diversidad real de abejas asociadas a paisajes agrícolas, en vista de la acelerada tasa de fragmentación y modificación del paisaje en Colombia (Instituto de Hidrología Meteorología y Estudios Ambientales de Colombia [IDEAM], 2020). Además, en los últimos años se ha intensificado la implementación de cultivos dependientes de polinizadores a nivel mundial (Aizen et al., 2019), por lo que conocer los polinizadores asociados a sistemas agrícolas se torna indispensable.
Muchos cultivos agrícolas son polinizados por abejas con rasgos funcionales específicos, como se evidencia en los cultivos de maracuyá (Calle et al., 2010; Arias-Suárez et al., 2014; González y González, 2015; Rodríguez-Calderón, 2016; Bezerra et al., 2019). Por esto, se deben ampliar las investigaciones enfocadas a la diversidad funcional de abejas y su relación con la polinización efectiva de los sistemas agrícolas donde actúan como visitantes florales, ya que los rasgos funcionales de las especies determinan la legitimidad de la polinización (Shaw et al., 2017). Además, estos estudios permitirán conocer las funciones ecológicas de abejas asociadas a cultivos en los que no actúan como polinizadores eficaces.
El papel ecológico de las abejas puede verse afectado por la interacción abeja-cultivo, debido a que muchos sistemas agrícolas, están expuestos a diferentes agroquímicos, que alteran el comportamiento y la morfología de las abejas (Chauzat et al., 2009). Por esta razón, se recomienda incrementar los estudios enfocados a la ecología funcional, no solo por los efectos adversos en la polinización, sino también en otros servicios ecosistémicos como el aporte en biomasa a las redes tróficas (Rodríguez y Velásquez, 2013). La ampliación de estos estudios es esencial para garantizar la conservación de abejas y sus servicios ecosistémicos en un mundo cambiante.
CONCLUSIONES
De acuerdo con la revisión realizada sobre la representatividad de las dimensiones taxonómica y funcional de la diversidad de abejas asociadas a cultivos, se encontró que ambas escalas de la diversidad son altas y con un buen potencial de ser mayor, a pesar de la asociación entre la fragmentación y pérdida de hábitat con la disminución de polinizadores. Es importante aclarar que esto no significa que los cultivos promuevan la diversidad de abejas, sino que estas pueden usarlos como recurso alternativo debido a la disponibilidad de flores. La variedad de rasgos funcionales y la riqueza y composición a escala taxonómica guarda congruencia entre sus valores para varios de los cultivos estudiados, sin embargo, esto no ocurre en otros como la palma, donde se favorece la ocupación de hábitat para un número alto de especies, pero con un mismo patrón de rasgos funcionales. Este comportamiento de las diversidades taxonómica y funcional tiene fuertes implicaciones para la conservación de las abejas y su función ecológica, por lo que requiere más profundidad de análisis con estudios in situ. Se identifica la necesidad de más estudios ecológicos que evalúen las variaciones en las métricas taxonómicas y funcionales de la diversidad de abejas, que analicen la congruencia entre ambas dimensiones de la diversidad a la luz de predictores locales y regionales propios de un paisaje agrícola y de cada cultivo en particular.

