











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroducción
Entre los insectos acuáticos, el orden Plecoptera constituye un componente importante de la fauna de ecosistemas dulceacuícolas debido a su alta abundancia y su amplia distribución. Este grupo de insectos cumple un papel destacado en la descomposición y recirculación de nutrientes y contribuye en la red trófica como alimento de otros insectos y vertebrados (Stewart y Stark 2002). No obstante, el conocimiento que se tiene del orden, a nivel nacional y regional, corresponde principalmente a estudios taxonómicos siendo escasos los trabajos sobre su biología y ecología en el Neotrópico (Roldán et al. 2014, Zúñiga 2010 y Zúñiga y Cardona 2009), a pesar de su amplio uso como indicadores de calidad del agua (Roldán 1998, Roldán y Ramírez 2008).
La biomasa es un importante atributo de la estructura de las comunidades o poblaciones; este atributo se puede evaluar como una propiedad colectiva o emergente (Breckling et al. 2005) debido a que permite cuantificar la cantidad de energía disponible por un organismo o grupo de organismos, en un determinado momento, al ser consumidos por organismos superiores en la red trófica (Chará-Serna et al. 2012, Tomanova et al. 2006), o bien pueden tomar la vía del detritus (Caraballo et al. 2014, Vander-Zanden et al. 2006).
Benke et al. (2001) y Waters (1979) resaltaron la importancia de conocer la historia de vida y el recambio de la biomasa en los organismos que conforman los sistemas acuáticos. Estas estimaciones resultan valiosas porque permiten analizar los aspectos ecológicos de una población y los datos pueden usarse para realizar estimaciones acerca de su producción en los ecosistemas. De esta forma, el conocimiento de la dinámica de la biomasa aporta información relevante sobre el flujo de energía en los ecosistemas (Benke et al. 2001, Benke y Huryn 2007). Por otra parte, el análisis de los hábitos alimentarios permite conocer el rol trófico de muchas comunidades de invertebrados en los sistemas acuáticos (Benke 1993, Benke y Wallace 1980). Lo anterior es útil para generar información relacionada con los flujos energéticos (captura y destino) y la dinámica de los ecosistemas acuáticos continentales (Dunne et al. 2004, Motta y Uieda 2004, Tomanova et al. 2006).
Aunque en el Neotrópico es poco lo que se ha estudiado sobre los hábitos alimentarios del orden Plecoptera, se conoce que la dieta ingerida por las ninfas puede variar dependiendo de la especie, el estado de desarrollo, la hora del día y la disponibilidad de recursos (Dorvillé y Froehlich 2001). Algunas especies, por ejemplo, son detritívoras o depredadoras durante todo su desarrollo, mientras otras podrían presentar cambios en sus hábitos alimentarios durante el proceso de desarrollo. El análisis del contenido estomacal de plecópteros de zonas templadas evidencia que son comunes los cambios de herbívoros-detritívoros en los primeros estadios a omnívoros-carnívoros en los periodos siguientes (Merritt et al. 2008). Estudios realizados previamente en la Sierra Nevada de Santa Marta los han definido como depredadores desde estados juveniles (Guzmán-Soto y Tamaris-Turizo 2014, Tamaris-Turizo et al. 2007). Sin embargo, estos trabajos no relacionaron la composición de las dietas con la talla de los organismos.
Con el fin de conocer 1) las posibles variaciones en la distribución de tallas entre los microhábitats y los muestreos, 2) la dinámica en las densidades entre microhábitats y las clases de talla, 3) las fluctuaciones temporales de la biomasa de los plecópteros y entre las clases de talla y 4) los posibles cambios en las dietas entre las clases de tallas del género Anacroneuria, se estudió la dinámica temporal del género Anacroneuria durante cuatro meses, en el río Gaira, Sierra Nevada de Santa Marta (SNSM). Además, se exploraron las relaciones entre la dinámica de tallas y la abundancia del género y las precipitaciones en un tramo del río Gaira. Se espera que los mayores aportes de biomasa estén dados por organismos de tallas pequeñas, debido a sus altas densidades, así como una preferencia por el microhábitat hojarasca por tener alta disponibilidad alimentaria y heterogeneidad. Con relación a las dietas, se pretende detectar diferencias en las clases de tallas de acuerdo con su desarrollo, las cuales, considerando la literatura, varían de detritívoras al inicio del desarrollo a depredadoras en la etapa madura (Merritt et al. 2008).
Materiales y métodos
Los muestreos se realizaron en la hacienda La Victoria, localizada en la parte media de la cuenca del río Gaira (departamento del Magdalena - norte de Colombia), ubicada a 900 m de altitud entre los 11°07’44,2” N y 74°05’35,8” O. El río tiene una longitud total de 32,53 km (con pendientes variables en la parte media) y atraviesa cinco tipos de clima, desde el cálido árido al nivel del mar hasta el frío muy húmedo a los 2850 m (Frayter et al. 2000, Tamaris-Turizo y López-Salgado 2006). En el bosque ribereño predominan especies con estrato arbóreo superior a los 24 m. El cauce posee corrientes rápidas y es un sistema pedregoso con una heterogeneidad de microhábitats favorable para el asentamiento de grupos de macroinvertebrados bentónicos (Grimaldo 2001, Rueda-Delgado et al. 2005). De acuerdo con los datos de la estación pluviométrica del IDEAM (código: 15010010), en la zona de estudio se registraron precipitaciones de 0, 107,5, 258,2 y 437,9 mm en los meses de marzo, abril, mayo y junio del 2005. Una mayor descripción del sitio de muestreo puede ser consultada en Tamaris-Turizo et al. (2013).
Recolecta del material biológico . Se realizaron recolectas cada 20 días durante cuatro meses, desde marzo hasta junio del 2005, lo cual correspondió a la finalización del periodo seco e inicio de las lluvias respectivamente. Se muestrearon tres sustratos: gravas, hojarasca y piedras, considerando que en dichos sustratos se han registrado las mayores abundancias del género (Tamaris-Turizo et al. 2007). En las gravas se utilizó una red Surber de 0,09 m2 y 250 µm de poro y se tomaron cuatro réplicas en el microhábitat de la grava, en cada sección del río: ribera izquierda, ribera derecha y centro del cauce. Dos réplicas de cada sección se usaron para el análisis de biomasa; las dos réplicas restantes se usaron para evaluar los contenidos estomacales. Posteriormente, las muestras se rotularon y fijaron en etanol al 96%.
De forma aleatoria se tomaron 100 g (peso húmedo) de hojarasca y se pesaron con un dinamómetro. Para determinar el área muestreada en la hojarasca se tomó una réplica en cada sector del río; las réplicas fueron transportadas al laboratorio, en donde se lavaron y secaron al aire. Posteriormente se midió el área de cada hoja empleando un planímetro manual Sokkia (Planix 6); se calculó el área promedio de los 300 g de hojarasca medida y se multiplicó por el número de réplicas por muestreo (12) para obtener el área total. Las piedras recolectadas tenían diámetros entre 10−20 ± 5 cm y alturas entre 18−23 ± 7 cm. La estimación del área superficial de cada piedra se realizó, mediante una regresión lineal simple, entre áreas de papel aluminio conocidas (cada piedra se forró con papel aluminio en campo) y su respectivo peso, lo que permitió obtener la ecuación de tipo Y = mX + b. El peso del papel aluminio (utilizado durante el primer muestreo para forrar las 12 piedras) se reemplazó en la ecuación anterior para determinar el peso (Bergey 2006).
Análisis de muestras. La identificación del material biológico se confrontó con las descripciones de Zúñiga et al. (2007) y Gutiérrez-Fonseca y Springer (2011). Los organismos se determinaron hasta el nivel de género y se separaron de acuerdo con las clases de tallas registradas (0,1−0,5; 0,6−1,0; 1,1−1,5; 1,6−2,0 y 2,1−2,5 cm). Para los cálculos de la biomasa se usaron 644 individuos, los cuales se secaron en estufa a 60 °C por 24 horas para obtener el peso seco constante y luego se llevaron a la mufla a 500 °C por 2 horas para determinar, por combustión, el peso seco libre de ceniza (PSLC) (Benke y Huryn 2007, Shieh et al. 2003). Todos los organismos se pesaron en una balanza analítica Ohaus Adventurer-Pro® (precisión ± 0,0001 g). Para los análisis de contenidos estomacales se extrajo el estómago a 640 individuos; el órgano fue fragmentado sobre un portaobjetos con glicerina y se realizaron observaciones en aumentos de 100X y 400X en un microscopio, discriminando los ítems alimentarios en cinco categorías: restos vegetales (RV), restos animales (RA), hongos (H), microalgas (M) y detritos (D) (Tamaris-Turizo et al. 2007). A cada estómago se le cuantificó la frecuencia de aparición de cada ítem alimentario, por cada campo ocular, hasta cubrir el total de la muestra.
Procesamiento de datos. Para explorar diferencias en el número de individuos entre los muestreos y en el número de individuos encontrados entre las tallas, así como también para establecer posibles diferencias en el número de individuos entre los microhábitats, y dado que los datos no cumplieron los supuestos de normalidad de residuos y/o homogeneidad de las varianzas, se utilizó la prueba de Kruskal-Wallis (H’) (Guisande et al. 2016). Para identificar cuáles fueron los grupos significativamente distintos se usó la prueba de contrastes de medias de Tukey (Guisande et al. 2016). Para conocer las relaciones entre las precipitaciones y el caudal con las tallas promedio y con la biomasa en los muestreos, se realizaron regresiones lineales; previamente se comprobó el cumplimiento de la homogeneidad y homocedasticidad de residuos mediante las pruebas Kolmogorov-Smirnov (Gross 2013) y Breusch-Pagan (Hothorn et al. 2013) respectivamente, y la ausencia de autocorrelación a través de la prueba de Durbin-Watson (Durbin y Watson 1951). Finalmente, para conocer la posible relación entre los ítems alimentarios y las clases de tallas, se realizó una tabla de contingencia con una prueba de Chi-cuadrado (X 2 ). Todos estos análisis se realizaron en el programa R (http://cran.r-project.org).
Resultados
Se recolectaron en total 1284 organismos (se utilizaron 644 para estimar la biomasa y 640 para evaluar el contenido estomacal). Los individuos se dividieron en cinco clases según el rango de las tallas. Las mayores abundancias coincidieron con los menores registros de caudales en los meses de marzo y abril (0,4 y 0,5 m3/s respectivamente); caso contrario se evidenció en el mes de junio cuando el caudal promedio fue 2,3 m3/s (Figura 1).

Figura 1 Abundancia promedio de Anacroneuria en los microhábitats durante los muestreos y los caudales promedio mensuales. M: muestreo. H: hojarasca, G: grava, P: piedra.
Frecuencia y distribución de tallas. Se detectaron diferencias en la distribución de las tallas durante los muestreos (H’=72; p<0,05; n=644). Las tallas menores de 1,15 cm presentaron altas abundancias durante todo el estudio, mientras que las tallas mayores de 2 cm no se encontraron en los muestreos 5 y 6 (Figura 2). También se registraron diferencias en el número de individuos entre muestreos (H’=82,9; p<0,05; n=644). El número de individuos recolectados en el segundo muestreo (inicio de abril) presentó diferencias significativas con respecto al registrado en los demás muestreos, en el cual las tallas menores de 1,1 cm se diferenciaron de las demás clases de tallas por tener la mayor abundancia. La comparación del número de individuos entre las tallas evidenció diferencias significativas (H’=100,3; p<0,01; n=644), de tal forma que las tallas pequeñas e intermedias (0,1−0,5 y 0,6−1,0 cm) presentaron, en todos los muestreos, el mayor número de individuos, mientras que las tallas mayores (1,6−2,0 y 2,1−2,5 cm) presentaron menores abundancias. No se encontraron diferencias significativas en la distribución de las tallas entre los diferentes microhábitats (p>0,05), pero si en el número de individuos encontrados en cada microhábitat (H’=24,8; p<0,05, n=644), siendo la hojarasca el microhábitat con mayores abundancias, seguido de la grava y la piedra (Figura 3).

Figura 2 Frecuencia de distribución de las tallas del género Anacroneuria en los muestreos realizados entre marzo y junio de 2005 en la parte media del río Gaira.

Figura 3 Abundancia de las clases de talla de Anacroneuria en los microhábitats hojarasca, grava y piedra.
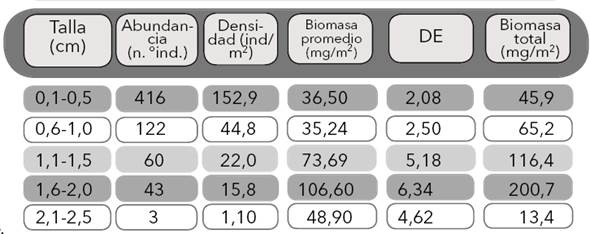
Biomasa. Los organismos de tallas pequeñas (0,1−0,5 y 0,6−1,0 cm) registraron las densidades más altas (152,9 y 44,8 individuos/m2 respectivamente), mientras que los individuos de tallas mayores (1,6−2,0 cm) registraron las menores densidades (1,10 individuos/m2). La biomasa presentó un patrón diferente a la densidad, debido a que los valores altos se registraron en los organismos con tallas intermedias (0,6−2,0 cm, promedio PSLC: 215,53 ± 4,67 mg/m2), en tanto que los valores bajos de biomasa coincidieron con los valores de densidades de los organismos con tallas grandes (2,1−2,5 cm, promedio PSLC: 48,90 ± 4,62 mg/m2) (Tabla 1). La prueba de comparación de la biomasa promedio de las tallas reflejó diferencias entre las clases de tallas (p<0,001; GL=17; F=10,83); la comparación de rangos múltiples evidenció diferencias altamente significativas entre las tallas 0,5−1,0 y 1,5−2,0 cm y 1,5−2,0 cm y 1,5−2,0 cm (p<0,001), lo cual indica que, aunque las tallas intermedias fueron las que más contribuyeron a la biomasa del género, las clases de talla 1,0−1,5 y 1,5−2,0 fueron las de mayor valor.
Tabla 1 Abundancia, densidad, biomasa promedio del género Anacroneuria (en peso seco libre de ceniza: PSLC) y biomasa total (PSLC) según las clases de tallas. DE: desviación estándar
Aunque en la Figura 4 se observa que la biomasa del género de Anacroneuria disminuyó durante los muestreos, pasando de 9,08 PSLC mg/m2 en el primero a 0,10 PSLC mg/m2 en el último, estos cambios no fueron significativos (H’=9,48; p>0,05; GL=5); la anterior tendencia se relacionó linealmente con el aumento de las precipitaciones en el sector (p<0,01; GL=4; r2=0,931). Una relación similar se observó entre la biomasa y el caudal (p<0,05; GL=4; r2=0,737), y entre la biomasa y la talla, pero en este último caso la relación no fue significativa (p>0,05; GL= 4; r2=0,102).

Figura 4 Biomasa promedio del género Anacroneuria en La Victoria y valores de precipitaciones mensual del corregimiento Minca. M: muestreo.
Hábitos alimentarios. En este análisis no se consideraron los organismos con tallas de 2,1 a 2,5 cm, debido a su baja abundancia y a que la mayoría de los estómagos se encontraron vacíos. Se observó que la dieta en las diferentes tallas fue variada y contenía todos los ítems alimentarios considerados en este trabajo, siendo los restos animales los más frecuentes (27,34%), mientras que los restos vegetales se presentaron en una menor frecuencia (10,79%) (Tabla 2). La prueba de Chi-cuadrado evidenció independencia entre el contenido estomacal y las diferentes tallas (p>0,05; GL=12; X 2 =7,27), lo cual sugiere que los ítems no están asociados con alguna categoría de talla en especial. Sin embargo, se destaca que la dieta de Anacroneuria incluyó restos animales entre los que dominaron simúlidos, quironómidos, partes esclerotizadas de efemerópteros y plecópteros, y en menor proporción, además de restos vegetales, hongos, microalgas y detritos.
Discusión
Frecuencia y distribución de tallas. Los cambios pluviométricos se relacionaron con cambios en la abundancia de los plecópteros. Durante los tres primeros muestreos (época seca e inicio de la época de lluvias) el número de individuos fue mayor. A partir del tercer muestreo se observó una disminución de la abundancia, coincidiendo con el inicio de las precipitaciones, las cuales provocaron cambios físicos en el lecho del río y la deriva de muchos organismos (Tamaris-Turizo et al. 2013, Wilcox et al. 2008). Durante los dos últimos muestreos (realizados en junio) se encontraron bajas abundancias. Sin embargo, como consecuencia de la estabilización de las precipitaciones y los niveles de caudal, no se observaron cambios importantes en el lecho durante las visitas a campo. Dichas relaciones entre las abundancias y las precipitaciones fueron registradas previamente en trabajos realizados en el trópico (Bispo et al. 2002, Bohórquez et al. 2011, Giacometti y Bersosa 2006, Rincón-H 1996, 2002, Tamaris-Turizo et al. 2007).
La abundancia alta de tallas pequeñas y la no diferencia en la distribución de las tallas durante todos los muestreos pueden ser evidencias de poblaciones que se encuentran en continuo crecimiento o que posiblemente no existe alternancia de generaciones entre las distintas especies. Sin embargo, esta información difiere de la registrada por Sweenney et al. (1995) en una especie semivoltina de Ephemeroptera, Euthyplocia hecuba (Hagen, 1861) de un río en Costa Rica y en trabajos realizados en zonas templadas donde la mayoría de las especies son univoltinas y evidencian dominio una o dos clases de tallas temporalmente por cohortes claramente diferenciadas (Landot et al. 2001, Meyer y Poepperl 2003). Usualmente, en las zonas templadas, los organismos no se encuentra en el sistema acuático durante el invierno; luego, al inicio de la primavera, dominan tallas pequeñas que incrementan su tamaño a lo largo del ciclo anual hasta alcanzar las mayores tallas en el otoño, donde se evidencian importantes emergencias de ninfas a sus estados adultos, tal como lo registró O´hop et al. (1984) en Estados Unidos, K’rno (1997) en Slovaquia, Benke et al. (2001) en el sureste de Estados Unidos y López-Rodríguez et al. (2009) en un río temporal de la Península Ibérica. Sin embargo, esta hipótesis debe ser comprobada con estudios poblacionales en ríos tropicales.
La mayor abundancia de individuos se encontró en el microhábitat hojarasca, similar a lo registrado en el río Guachaca (al norte de la SNSM) (Rueda et al. 2005), aunque difiere de lo encontrado en la parte alta del río Gaira (Tamaris-Turizo et al. 2007) y de los estudios realizados en el Valle, donde se observó preferencia por los sustratos hojarasca, piedra y grava respectivamente (Ballesteros 2004, Zúñiga y Cardona 2009). Merritt et al. (2008) sugieren que las ninfas de Plecoptera tienen especificidad por la temperatura del agua, tipo de sustrato y tipos de corrientes. Nuestros resultados difieren de las preferencias de sustrato observadas en el trópico. La preferencia por la hojarasca en este trabajo se debe posiblemente a que este sustrato ofrece disponibilidad de alimento pese a su poca estabilidad (están propensos a los arrastres por cambios en el caudal). Por su parte, el sustrato grava es más estable que las hojas, ofrece refugio y disponibilidad de alimento, en contraste con el sustrato piedra que generalmente presenta una biopelícula adecuada para la alimentación de los raspadores, pero es fácilmente lavada en las épocas de lluvias, debido a los aumentos de caudales, el arrastre de sedimentos y el aumento de la turbidez del agua.
Biomasa. La talla más pequeña (0,1−0,5 cm) presentó la densidad más alta, mientras que las tallas 2,1−2,5 cm fueron las menos abundantes. Aunque las tallas intermedias presentaron densidades bajas registraron los valores más altos de biomasa. La talla grande presentó un valor bajo de densidad y biomasa coincidiendo con lo reportado en especies de plecópteros del río Minnesota - Estados Unidos (Krueger y Cook 1984) y en efemerópteros de Estados Unidos (Taylor 2001). Ambos estudios coincidieron en que las tallas pequeñas e intermedias presentaron un aporte constante, debido a que se mantuvieron en continúa renovación durante el periodo de estudio. Lo anterior evidencia que las especies del género pueden ser multivoltinas y sus ciclos de vida menores de un año. En este trabajo, el género Anacroneuria mantuvo cantidades importantes de biomasa e individuos disponibles durante los meses de marzo a mayo, fechas que, según observaciones de campo, corresponden a periodos de emergencia (transformación de ninfa a adulto).
Los valores de biomasa registrados en este trabajo son mayores que los registrados por Ramírez y Pringle (1998) en varios taxones de insectos acuáticos distintos a plecópteros en un río neotropical de Costa Rica. Las biomasas de Anacroneuria se asemejan a las registradas por el género Tricorythodes (7,28 PSLC mg/m2) y a la familia Elmidae (7,18 PSLC mg/m2), pero son mucho menores que los observados en organismos de mayores tallas como la familia Corydalidae (1,19 PSLC mg/m2). Esto demuestra que el género Anacroneuria es un grupo de gran importancia para la disponibilidad de energía en los niveles tróficos superiores o inferiores (vía descomponedores) y que es probable que aporte más energía que organismos de tallas mayores.
En este estudio se observó una fuerte relación entre el aumento de las precipitaciones y los caudales con la disminución de la biomasa del género. Se puede considerar que el género Anacroneuria posee una alta tasa de crecimiento, evidenciado en el rápido incremento de la biomasa durante un periodo corto de tiempo. No obstante, estas dinámicas pueden ser variables según las condiciones ambientales e interacciones ecológicas, lo cual abriría el campo para el análisis de este aspecto desde otras perspectivas como la producción secundaria (Benke 1993). Desde el punto de vista trófico, esta biomasa disponible mantiene constante el suministro energético para depredadores de niveles tróficos superiores (Añón-Suárez y Albariño 2001), hecho que podría considerar a este grupo como fuente potencial de alimentación disponible entre marzo y junio.
Análisis de Contenido Estomacal. Los restos animales fueron el componente de mayor importancia en la dieta del género Anacroneuria, lo que confirma su comportamiento depredador de otros grupos de insectos, seguido por los detritos. Al parecer, la relación entre estos ítems puede ser explicada por el hecho que los restos animales o presas encontradas fueron en su mayoría larvas de simúlidos, quironómidos (recolectores, filtradores) y efemerópteros (recolectores o raspadores), importantes intermediarios en la conversión del material vegetal y de los detritos para la biomasa de los plecópteros y que dominan en estos sustratos (Siegfried y Knight 1976). Estos grupos de organismos tienen una alta abundancia en el microhábitat hojarasca, hecho posiblemente asociado con las altas densidades de plecópteros en dichos sustratos. En menor proporción se encontraron fragmentos de otros organismos del género Anacroneuria, sugiriendo el canibalismo referenciado por Tamaris-Turizo et al. (2007), lo cual debe ser confirmado a través de estudios específicos. En cuanto al detrito, se considera que este podría haber sido ingerido accidentalmente con la presa animal durante su actividad alimentaria (Riaño et al. 1997), y podría estar relacionado con el material consumido por la presa de manera incidental o puede ser el producto de restos de animales y vegetales ya procesados por plecópteros o sus presas. Aunque no se puede descartar la posibilidad que estos organismos cambien sus dietas de acuerdo con la disponibilidad de recursos en el medio y su capacidad de seleccionar algún recurso en particular.
Los contenidos estomacales mostraron ítems alimentarios, tal como se había registrado previamente para la familia Perlidae (Dorvillé y Froehlich 2001). Los cambios de las dietas podrían ocurrir durante el desarrollo, al parecer dependiendo de la estación o los cambios en la abundancia de la presa (Teslenko 1997), coincidiendo con lo encontrado en las ninfas de la parte alta (Tamaris-Turizo et al. 2007) y media del río Gaira (Guzmán-Soto y Tamaris-Turizo 2014). Una de las limitantes de este trabajo fue que la preservación en alcohol puede producir que los insectos regurgiten. Sin embargo, durante todo el estudio menos de 20 individuos presentaron estómagos vacíos.

