











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
The global demand for nitrogen (N) fertilizers is increasing. In the case of urea, consumption was 174.29 million t worldwide in 2016 (IFA, 2017). In Peru, the import volume of urea for agricultural purposes was 398 961 t in 2019 (AGRODATA PERÚ, 2020).
Key factors determine crop productivity and one of them is N fertilizers such as urea (Campillo et al., 2007). Intensive use of N fertilizers can cause negative effects such as acidification or nitrification of soils (Tong and Xu, 2012; FAO, 2014).
The existence of heavy metals, such as lead (Pb), in agricultural soils is mainly due to irrigation with contaminated water (Méndez et al., 2009). Additionally, the use of pesticides, such as Pb arsenate, in Peruvian agriculture (Iannacone et al., 2009) until its ban in 2013 (MINAM, 2013) and the atmospheric deposition of industrial and vehicular air particles are some of the causes of the presence of these metals in the soil (Nolasco, 2001; Bellido, 2018).
The bioaccumulation of heavy metals, such as Pb, in crops is influenced by factors such as weather, atmospheric depositions, soils with concentrations of other heavy metals, use of fertilizers, the nature of the soil in which vegetables are grown, and agricultural areas with a long-term history of treated or untreated wastewater for irrigation (Sinha et al, 2005).
Anthropogenic factors, such as fertilizers, can contribute to the absorption of heavy metals in crops by modifying the physical-chemical properties of the soil, such as pH, organic matter, and bioavailability of heavy metals in the soil (Sharma et al., 2008). According to Yu-kui et al. (2009), corn (Zea mays L.) absorbs mineral elements and heavy metals due to the application of N fertilizers. In the case of cabbage (Brassica oleracea L.), the plant has heavy metal hyperaccumulator characteristics (Yanqun et al., 2005). Also, spinach (Spinacia oleracea L.) has a high potential to accumulate heavy metals (Kumar et al, 2013).
The absorption of heavy metals by the different parts of plants (root, leaves, and fruit) is the first step for their incorporation in the trophic chain (Méndez et al., 2009). Therefore, it represents a health hazard due to the negative effects these heavy metals can cause (Salazar et al, 2012).
The objective of this research was to determine the effect of urea on the absorption of Pb in corn (Z. mays), spinach (S. oleracea) and cabbage (B. oleracea var. capitata), and to determine which of these plant species absorbs more Pb in its foliage and how urea influences soil acidification.
Materials and methods
The study was carried out in the Environmental Engineering greenhouse of the Universidad Cientifica del Sur (UCSUR), Lima, Peru, and in the Laboratory of Ecology and Animal Biodiversity of the Universidad Nacional Federico Villarreal (LEBA), Lima, Peru. A total of112.5 kg of greenhouse substrate was prepared with the following characteristics: 40% peat moss, 30% farm soil, 10% coarse sand, 10% compost, and 10% horse manure. Two samples were analyzed for the selection of farm soil; one belonged to the district of Huachipa and the other to the district of Lurin, Lima, Peru. The samples were analyzed in the Laboratory of Soil, Plants, Water and Fertilizer Analysis (LASPAF) of the Faculty of Agronomy of the National Agrarian University La Molina (UNALM), Lima, Peru (Tab. 1). The protocol for the determination of the field capacity (FC) followed the procedure of Rai et al. (2017).
TABLE 1 Analysis of the farm soil for the preparation of the greenhouse substrate.

EC - electrical conductivity; CaCO3 - calcium carbonate; OM - organic matter; P - Phosphorus; K - Potassium; Al+ 3 + H+ - exchangeable acidity; Pb - Lead; FC - Field capacity; N - Nitrogen.
The soil chosen for the substrate was from the district of Lurin because the concentration of Pb was below the environmental quality standard (EQS) of Peru for agricultural land (0.07 g kg-1). In contrast, the soil from Huachipa had a Pb concentration above the EQS of Peru for agricultural land. The greenhouse substrate was mixed with the respective concentration of Pb (0.5 g Pb kg-1 of soil) in a plastic container. The contaminant used was Pb acetate, and the urea doses were added to the substrate according to the treatments listed in Table 2.
TABLE 2 Treatments to evaluate the effect of urea on Pb absorption in corn (Zea mays), spinach (Spinacia oleracea), and cabbage (Brassica oleracea var. capitata).

(*) This value (0 g) corresponds to the application of Pb acetate on the soil and not to the value of Pb in the composition of the soil of the farm (0.026 g kg-1) which is below the Environmental Quality Standard (EQS) of Peru (0.07 g Pb kg-1 soil) for agricultural land.
The plant materials used for the present experiment were spinach (S. oleracea), and cabbage (B. oleracea var. capitata) seeds acquired at the nursery of the Universidad Nacional Agraria La Molina (UNALM), Lima, Peru. Corn (Z. mays) seeds were purchased from the Corn Program of UNALM, Lima, Peru. The corn cultivar was Indurata; for spinach, the cultivar was Viroflay, and for cabbage, the cultivar was Brunswick.
A completely randomized block design was established with five treatments (Tab. 2) with three replicates per crop. The experimental unit was a 15 L plastic container with 2.5 kg of greenhouse substrate. The seeds were sown directly in the substrate except for the cabbage that was transplanted after two weeks of germination. Three seeds were sown in each pot, and after three weeks of germination, only one plant was left per pot. Only a general linear model (GLM) factorial two-way ANOVA was performed to evaluate the effects of urea and Pb application on foliar Pb and N contents in corn, spinach, and cabbage.
Plant growth and development were under greenhouse conditions. Temperature and relative humidity data were collected from the nearest SENAMHI weather Von Humboldt station located in La Molina district, Lima, Peru (12°05 S, 76°57' W, and 243.7 m a.s.l.). The following temperature (16.62°C, range = 12.0-27.9°C) and humidity parameters (82.27%, range = 75.44-84.71%) were recorded.
Plants were irrigated with demineralized water from the time of sowing to one week before harvest. The volume of irrigation water per pot was enough to approximate the humidity of field capacity (Rai et al., 2017). The field capacity of the substrate was not modified by the use of peat moss and coarse sand, which prevented leachate of excess water and, therefore, a possible loss of N, Pb, or other nutrients from the substrate. Each pot was irrigated twice a week; the first irrigation had a volume of 1000 ml, and then plants were irrigated with a volume between 200 and 500 ml according to the need of each plant (Ugaz et al., 2000). In total, each plant of corn utilized 7600 ml, spinach 6800 ml, and cabbage 5800 ml.
During plant growth and development, the soil pH was evaluated with a potentiometer (AAMS, MHS Iberica, Madrid, Spain) (1:1 soil:water ratio) every 7 d to determine its behavior (Collins et al., 1970). Biometric measurements were performed at the end of the trial, at the time of harvest at 93 d for corn, spinach, and cabbage, without considering their different periods of growth of each crop but implementing the same period of exposure to urea and Pb. A portable electronic balance (0.1 g readability, SPJ6001, Ohaus, Melrose, USA) was used to determine the fresh weight and dry weight of plants. For measuring the height of the plant, a measuring tape (±1 mm) was used. For corn, plants were measured from the ground level to the flag leaf (the last leaf that emerges from the stem), and the spinach and cabbage plants were measured from the ground level to the highest leaf.
A portable chlorophyll meter (SPAD-502, Konica Minolta, Tokyo, Japan) was used to determine the chlorophyll index. Three measurements were taken between the main nerve and the edge of the central leaflet of each plant to obtain the average chlorophyll index in SPAD units (Yllanes et al., 2014; Hurtado et al., 2017). The ImageJ program version 1.50i was used to determine the leaf area. Photographs of all the leaves of each plant were taken before sending them to the laboratory.
The plant shoot of each crop was placed in paper bags and then taken to the LASPAF of UNALM, Lima, Peru, where the concentration of Pb and N in leaves was determined. The reading or measurement of the concentration of Pb was performed on an atomic absorption spectrophotom-eter (AAnalyst 200, Perkin Elmer, Waltham, USA) using a wavelength of 283.3 nm. The analysis of plant tissue for the determination of total N was carried out using the method described by Kjeldahl, as described in Sáez-Plaza et al. (2013), which is based on transforming the entire organic fraction of N (proteins, amino acids) into the ammonium sulfate, and which can be distilled to obtain the N contained in the leaf sample.
Data were analyzed using the statistical program SPSS, version 24.00. An analysis of variance (ANOVA) with a confidence of 95% was performed to analyze the differences between the control and the treatments as well as the relationship between the parameters of the plants. Additionally, the non-parametric Kruskal-Wallis test was used for those variables that did not meet all the requirements (normality and homogeneity of variance). Posteriorly, the Tukey test was performed to determine the significant differences for each of the parameters. The observed effect concentration (NOEC) and lowest observed effect concentration (LOEC) were also determined. The negative effects of LOEC and NOEC refer to any increase or reduction for each of the evaluated variables. In addition, for NOEC and LOEC, the highest statistically divergent values were used compared to the control in the case of observing a non-linear relationship. A general linear model (GLM) factorial two-way ANOVA was performed to evaluate the effects of urea and Pb application on crops of corn, spinach, and cabbage.
The relationship between the urea dose, acidity, and soil pH were also analyzed as well as the relationship between the soil pH, and the concentration of Pb were analyzed by Pearson's correlation, with 95% confidence.
The data obtained from the LASPAF of UNALM, Lima, Peru regarding the concentration of Pb in the leaves of corn, spinach, and cabbage were compared with the maximum permissible levels (MPLs) of Codex Alimentarius and of other countries (MINCOTUR, 2017). Codex Alimentarius is the regulation that governs Peruvian legislation because there was no MPL for food until now.
The Bioconcentration factor (BCF) is the ratio of the elements contained in the foliar part of the plant concerning the soil (Vyslouzilova et al., 2003). If the quotient is greater than or equal to one, the plant is considered a Pb accumulator (Santoyo-Martinez et al., 2020). Pb concentration in the leaf structure was estimated for the foliar parts. In the case of the control treatment, the data from the soil analysis of the used farmland (22.32 mg kg-1) was considered for determining Pb concentration in the soil, and for the other treatments, the Pb acetate concentration (500 mg kg-1) that was applied.
Results
Fifteen experimental units were evaluated for corn, 15 experimental units for spinach, and only nine experimental units for cabbage, because in the third week the plants of T3 and T4 died, possibly due to the application of dose levels of urea and Pb totaling 39 experimental units that grew and developed for 93 d.
Pb and foliar nitrogen
There were significant differences between treatments for foliar Pb in spinach, with T4 showing the highest concentration of Pb compared to corn and cabbage. Significant differences were evident between treatments for foliar N in cabbage, with T2 showing the highest concentration compared to the control and T1. NOEC and LOEC had the lowest statistical values for foliar Pb and N in cabbage (Tab. 3). The GLM factorial two-way ANOVA indicated significant differences in the treatments with urea and Pb application on foliar Pb (F=4.09, P<0.05), but no differences were observed on the foliar N (F=1.70, P=0.18) of corn, spinach, and cabbage. Also, the factorial two-way ANOVA indicated significant differences between crops of foliar Pb (F=10.51, P<0.05) and foliar N (F=16.82, P<0.05). Finally, the factorial two-way ANOVA indicated significant differences in the interaction between treatments of urea and Pb application and the crops on foliar Pb (F=2.78, P<0.05) but no differences were observed on foliar N (F=0.88, P=0.52) of corn, spinach, and cabbage.
TABLE 3 Comparison of the effects of urea and Pb application on foliar Pb and N in corn (Zea mays), spinach (Spinacia oleracea), and cabbage (Brassica oleracea var. capitata).

F - Fisher statistic; *the Kruskal Wallis test was used because the data did not meet the normality test. P- Level of significance; NOEC - maximum concentration in which no negative effects are observed compared to the control; LOEC - minimum concentration in which negative effects are observed compared to the control; (±) Standard error; ND - not determined; **for NOEC and LOEC, the highest statistically different values were used in relation to the control in the case of a non-linear relationship; equal lowercase letters in the same column indicate that the means are statistically equal according to the Tukey test (P<0.05).
Corn
For the chlorophyll index, the treatments in which urea was applied (T2, T3, and T4) had the highest SPAD values compared to the treatments without urea application (control and T1). T2 and T3 showed greater fresh weight than the control, and T1, T2, T3 and T4 showed greater dry weight than the control. T2 plants showed greater height compared to the control, and the leaf area for T1, T2, and T3 was greater than in the control. NOEC and LOEC had the statistically lowest values for chlorophyll index, fresh weight, and height in corn (Tab. 4).
TABLE 4 Progressive effect of urea and Pb application on the five physiological variables evaluated in leaves of corn (Zea mays).

F - Fisher statistic; *the Kruskal Wallis test was used because the data did not meet the normality test; SPAD - chlorophyll index value; P- level of significance; NOEC - maximum concentration to which no negative effects are observed concerning the control; (LOEC) minimum concentration where negative effects are observed for the control; ± standard error; ND - not determined; **for NOEC and LOEC, the highest statistically different values were used in relation to the control in the case of a non-linear relationship; equal lowercase letters in the same column test indicate that the means are statistically equal according to the Tukey (P<0.05).
Spinach
There were no significant differences between treatments for dry weight, height, and leaf area. Regarding the chlorophyll index and fresh weight, T2 was higher compared to the control. NOEC and LOEC had the statistically lowest values for fresh weight in spinach (Tab. 5).
TABLE 5 Progressive effect of urea and Pb application on the five physiological variables evaluated in leaves of spinach (Spinacia oleracea).

F - Fisher statistic; *the Kruskal Wallis test was used because the data did not meet the normality test; SPAD - chlorophyll index value; P Level of significance; NOEC - maximum concentration to which no negative effects are observed concerning the control; LOEC - minimum concentration where negative effects are observed concerning the control; ± standard error; ND - Not determined; **for NOEC and LOEC, the highest statistically different values were used in relation to the control in the case of a non-linear relationship; equal lowercase letters in the same column indicate that the means are statistically equal according to the Tukey test (P<0.05).
Cabbage
In cabbage, there were no significant changes between treatments for fresh weight, dry weight, height, and leaf area. For the chlorophyll index, T2 was higher compared to the control. NOEC and LOEC had the statistically lowest values for the chlorophyll index in cabbage (Tab. 6).
TABLE 6 Progressive effects of urea and Pb application on the five physiological variables evaluated in leaves of cabbage (Brassica oleracea var. capitata).

F - Fisher statistic; *the Kruskal Wallis test was used because the data did not meet the normality test; SPAD - chlorophyll index value; P- level of significance; NOEC - maximum concentration to which no negative effects are observed concerning the control; (LOEC) minimum concentration whe e negative effects are observed for the control; ± standard error; ND - not determined; **for NOEC and LOEC, the highest statistically different values were used in relation to the control in the case of a non-linear relationship; equal lowercase letters in the same column test indicate that the means are statistically equal according to the Tukey (P<0.05).
Soil pH
Soil pH in the control and Tj for corn was slightly alkaline during the time of the experiment. T2 remained slightly acidic until day 36, and then acidity increased. Treatments T3 and T4 were slightly acid pH until day 50 (Fig. 1).

FIGURE 1 Comparison of the five treatments in corn (Zea mays) for soil pH behavior. Treatment 1 (T,): urea application level (0 g urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil); Treatment 2 (T2): (0.5 urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil); Treatment 3 (T3): (1.0 g urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil); Treatment 4 (T4) (1.5 g urea kg1 soil) and Pb application level (0.5 g Pb kg-1 soil). Bars represent the standard error.
The soil pH in spinach in control and T[ was slightly alkaline during the 93 d. However, T2, T3 and T4 remained slightly acidic until days 29,50, and 64, respectively (Fig. 2).

FIGURE 2 Comparison of the five treatments on spinach (Spinacia oleracea) for soil pH behavior. Treatment 1 (T1): urea application level (0g urea kg-1 soil) and Pb application level (0.5 g Pb kg-1 soil); Treatment 2 (T2): (0.5 urea kg-1 soil) and Pb application level (0.5 g Pb kg-1 soil); Treatment 3 (T3): (1.0 g urea kg-1 soil) and Pb application level (0.5 g Pb kg-1 soil); Treatment 4 (T4) (1.5 g urea kg-1 soil) and Pb application level (0.5 g Pb kg-1 soil). Bars represent the standard error.
Regarding the soil pH in cabbage during all the assays, the control and T[ were slightly alkalized. T2 remained slightly acidic until day 22, and from day 29, it was somewhat alkalized; T3 and T4 were slightly acidified until day 15 (Fig. 3).

FIGURE 3 Comparison of the five treatments in cabbage {Brassica olerácea var. capitata) for soil pH behavior. Treatment 1 (T,): urea application level (0 g urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil); Treatment 2 (T2): (0.5 urea kg1 soil) and Pb application level (0.5 g Pb kg-1 soil); Treatment 3 (T3): (1.0 g urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil); Treatment 4 (T4) (1.5 g urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil). Bars represent the standard error.
The pH behavior among the treatments for the three plants together was similar; the control and T1 were slightly alkalized, and T2, T3 and T4 were acidified temporarily to approach neutrality or become somewhat alkalized (Fig. 4).

FIGURE 4 Comparison of the five treatments combined in corn (Zea mays), spinach [Spinacia oleracea), and cabbage [Brassica oleracea var. capitata) for soil pH behavior. Treatment 1 (T,): urea application g urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil); Treatment level (0 g urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil] 2 (T2): (0.5 urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil); treatment 2 (T2): (0.5 urea kg1 soil) and Pb application level (0.5 g Pb Treatment 3 (T3): (1.0 g urea kg1 soil) and Pb application level (0.5 g Pb kg1 soil); Treatment 3 (T3): (1.0 g urea kg1 soil) and Pb application kg"1 soil); Treatment 4 (T4) (1.5 g urea kg1 soil) and Pb application level level (0.5 g Pb kg1 soil); treatment 4 (T4) (1.5 g urea kg1 soil) and Pb (0.5 g Pb kg"1 soil). Bars represent the standard error. application level (0.5 g Pb kg1 soil). Bars represent the standard error.
We observed that the urea dose had a direct relationship with the acidity time of the soil for corn, spinach, and cabbage, and the urea dose had a negative relationship with the soil pH for corn and spinach (Tabs. 7 and 8). We also observed that the soil pH had a negative correlation with foliar Pb contents only for spinach (Tab. 9).
TABLE 7 Pearson correlation coefficient values among the doses of urea and the acidity time in corn [Zea mays), spinach [Spinacia oleracea), and cabbage [Brassica oleracea var. capitata).
| Dose of urea (g urea kg1 soil) | Soil acidity time (d) | P |
|---|---|---|
| Corn | r = 0.98 | 0.00 |
| Spinach | r = 0.98 | 0.00 |
| Cabbage | r = 1.00 | 0.00 |
P<0.05 is statistically significant, indicating a relationship between the dose of urea and the acidity time of the crops.
TABLE 8 Pearson correlation coefficient values illustrating the dose of urea and its effect on the soil pH in corn (Zea mays), spinach (Spinacia oleracea), and cabbage (Brassica oleracea var. capitata).
| Dose of urea (g urea kg-1 soil) | pH of the soil | P |
|---|---|---|
| Corn | r = - 0.97 | 0.01 |
| Spinach | r = - 0.97 | 0.01 |
| Cabbage | r = -0.98 | 0.14 |
P<0.05 is statistically significant, indicating a relationship between the dose of urea and the soil pH.
TABLE 9 Pearson correlation coefficient values showing the relationship between the soil pH and the foliar Pb contents in corn (Zea mays), spinach (Spinacia oleracea) and cabbage (Brassica oleracea var. capitata).
| pH of the soil | Foliar Pb contents (mg kg-1) | P |
|---|---|---|
| Corn | r = -0.17 | 0.79 |
| Spinach | r = -0.95 | 0.01 |
| Cabbage | r = 0.73 | 0.48 |
P<0.05 is statistically significant, indicating a relationship between soil pH and foliar Pb contents of the crop.
Maximum permissible limit (MPL) of Pb in food
We observed that the concentration of Pb in the leaf structure of spinach, corn, and cabbage in all treatments and control exceeded the Pb MPL of both the Codex Alimentarius (Peruvian legislation) and the limits of other countries (Tab. 10).
TABLE 10 Comparison of Pb contents in leaves of corn (Zea mays), spinach (Spinacia oleracea), and cabbage (Brassica oleracea var. capitata) with the maximum permissible limits of the Codex Alimentarius (Peruvian legislation) and legislation of other countries.
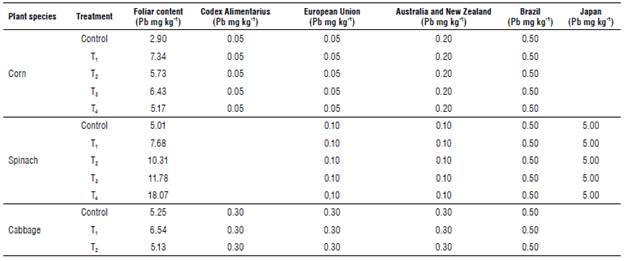
Treatment 1 (T,): urea application level (0 g urea kg-1 soli) and Pb application level (0.5 g Pb kg-1 soli); Treatment 2 (T2): (0.5 urea kg-1 soli) and Pb application level (0.5 g Pb kg-1 soli); Treatment 3 (T3): (1.0 g urea kg-1 soil) and Pb application level (0.5 g Pb kg-1 soil); Treatment 4 (T4) (1.5 g urea kg-1 soil) and Pb application level (0.5 g Pb kg-1 soil).
The BCF values were less than 1 in all the treatments for spinach, corn, and cabbage that indicate that these plants are not considered efficient accumulators of Pb (Tab. 11).

Discussion
Effect of treatments on chlorophyll contents, fresh weight, dry weight, and leaf area
Chlorophyll contents of corn, spinach (T2), and cabbage were higher in the treatments where urea was applied and when Pb concentration increased in the soil. T3 and T4 of spinach showed a decrease in chlorophyll contents, both fresh or dry weight, and leaf area, possibly due to the presence of high doses of urea and the presence of Pb taken from the soil (Tabs. 4-6). According to Sonbai et al. (2013), the increase in chlorophyll levels is because N from urea can be absorbed through the roots and used to form more chlorophyll (Ai and Banyo, 2011; Surya, 2013). In Cannabis sativa, an increase in leaf number, plant height, and root length is observed at high doses of urea with contaminated soil with Pb, when compared to a control (Hadi et al., 2014). Khan et al. (2016) noted that Pb could alter chlorophyll biosynthesis and N metabolism.
The fresh weight in corn (T2 and T3) was higher in the treatments where urea was applied, and in spinach the greater fresh weight was observed in T2 (Tabs. 4-6). Nitrogen is an essential element since it is a component of proteins and chlorophyll. Therefore, it is essential for photosynthesis, vegetative (fresh weight), and reproductive growth, and often determines the yield of corn (Iqbal et al., 2006). According to Pangaribuan et al. (2018), increasing N levels can also increase the fresh weight per plant, which can be attributed to the greater availability of N with the higher rate of N fertilizer.
In spinach, T4 had the highest concentration of foliar Pb (Tab. 3) and, at the same time, the lowest fresh weight (Tab. 5). Similar effects are observed in studies with Plan-tago major L. and spinach, where reduction in fresh and dry weight is observed in the different parts of the plant when exposed to Pb (Kosobrukhov et al., 2004; Lamhamdi et al., 2013). The specific symptoms of exposure to Pb in plants are characterized by the reduction of dry weight in the shoot since Pb causes a decrease in the protein content in tissues and an alteration in the lipid composition (Cao et al., 2015). Alia et al. (2015) note that, in S. oleracea, an increase in the doses of Pb in the soil with different doses (mg kg-1) reduces the growth parameters (shoot and root lengths, biomass and number of leaves). Hart et al. (2005) and Zhou et al. (2018) studied the effects of Pb doses in the soil on plant shoot and root growths and photosynthesis in various types of plants. Under Pb stress, the chloroplasts are deformed, and the thylakoid lamellae are gradually expanded, resulting in the separation from the cell wall and the eventual shrinkage of the nucleus (Zhou et al., 2018).
In corn, the treatments where Pb was applied (T1, T2, and T3) obtained a greater dry weight compared to the control (Tab. 4). According to Hu et al. (2015), the reduction in dry weight in plants that are exposed to Pb shows variability depending on the species, as shown in this research for corn, spinach, and cabbage that had different Pb concentration in the leaves. This was also observed for P. major plants, where a decrease in dry weight was observed. This effect was also noted in Z. mays, where there was an apparent increase in dry weight with the increase in the synthesis of polysaccharides in the cell wall.
In cabbage, there were no significant changes between treatments for fresh weight, dry weight, height, or leaf area. Paschalidis et al. (2013) observed that cabbage plants do not show growth disturbance, and the plants show no symptoms of phytotoxicity as a result of Pb application. These studies note the tolerance of cabbage plants to high concentrations of Pb in the soil. These authors found that fresh weights of leaves of cabbage significantly increase in the lowest applications (10 mg Pb kg-1) compared to the control treatment (Paschalidis et al., 2013). The stimulatory effect of Pb on chlorophyll contents in cabbage in T2 could be attributed to the fact that metal ions can serve as activators of enzymes in cytokinin metabolism that accelerates the growth of plants. Cytokinins can delay senescence while maintaining the chlorophyll and photosynthetic activity of leaf tissues of cabbage (Paschalidis et al., 2013).
Nas and Ali (2018) explain that Pb accumulation in the soil inhibits germination of seeds and retards growth of seedlings, decreases the germination percent, germination index, root/shoot length, number of leaves, tolerance index and dry mass of roots and shoots. Lead inhibits photosynthesis, decreases mineral nutrition, water balance, and enzyme activities. Lead inhibits chlorophyll synthesis by causing reduced uptake by plants of essential elements such as magnesium and iron. However, the presence of urea in this study caused a different behavior with an increase in dry and wet weight, and a decrease in leaf area with higher urea concentrations and constant Pb values according to the treatments in corn. In spinach, the highest values in all variables analyzed were seen in T2. In cabbage, most of the variables showed no changes compared to the control, and only the chlorophyll index showed an increase in T1 and T2 compared to the control.
Crop response to treatments
A decrease in height was observed in corn in T3 and T4 that were the treatments with the highest rates of urea application (Tab. 4). On the other hand, different studies show that increasing N doses has positive increased plant height in corn (Kaplan et al., 2016; Chauhdary et al., 2017).
In cabbage, T3 and T4 were not successful because the plants died at 21 d after sowing. The Pb uptake may have influenced this effect under greater urea concentrations than by the Pb concentration in the soil itself when it existed at concentrations that do not exceed 1 g of Pb kg-1 soil. Pb accumulates in higher amounts in the different organs of the plant, and non-tolerant plants may die (Rodriguez et al., 2006). Likewise, the acidity of the soil may also have influenced because both treatments had the lowest pH value (5.87) during the first weeks. Cabbage has an optimum pH range between 6.2-6.5 (FAXSA, 2002) and is intolerant of acidity (Ugaz et al., 2000). Bioavailability of Pb in plants depends on the acidity of the soil and pH values (Lopez et al., 2005). Paschalidis et al. (2013) observed that 500 mg Pb kg-1 significantly increases Pb accumulation in roots of cabbage compared to the control treatment. Pb accumulation in Pb-treated plants was not dose-related, but it significantly increased only with the greatest dose of Pb, compared to the control treatment. The lower Pb accumulation was probably due to the relatively low mobility of this metal caused by the binding of Pb to clay minerals. Pb is taken up by plants only in very low amounts, with its largest proportion being accumulated in roots, and only a small portion is transported to the shoots.
Spinach obtained higher concentrations of foliar Pb compared to corn and cabbage (Tab. 3). Spinach was the Pehafiel-Sandoval and lannacone: Effect of urea on lead absorption in corn (Zea mays L.), only plant species that increased Pb levels in the leaves by increasing urea concentrations and the presence of Pb in the soil. This could be explained by the fact that spinach differs from corn and cabbage in its uptake of heavy metals and its sensitivity to Pb, which is a factor that depends on certain soil conditions and root morphology (Hart et al., 2005). There are plant species with the capacity to store higher concentrations of specific metals in the shoots (Pilon-Smits, 2005; Callahan et al., 2006). In spinach, this could be related to a greater capacity to resist the oxidative stress generated by Pb (Lamhamdi et al., 2010). Spinach also can accumulate heavy metals, such as Pb, due to a greater translocation capacity for Pb compared to other plant species (Kumar et al., 2013). Thanh et al. (2013) and Khan et al. (2016) point out that the capacity of plants to absorb Pb depends on each plant species, so this was observed in the present research with spinach showing higher levels of foliar Pb than in the other two plant species.
A higher concentration of foliar Pb was observed in spinach in T4 (Tab. 3) and, at the same time, a greater reduction in the leaf area compared to T1 (Tab. 5) where the concentration of foliar Pb was lower (Tab. 3). This symptom may be related to the inhibitory effect of Pb on the growth of the different plant shoots (Kopittke et al., 2007), where a lower number of leaves, a smaller size, and a greater fragility are observed (Gupta et al., 2009). This can be attributed to metabolic disorders of nutrients (Kopittke et al., 2007; Gopal and Rizvi, 2008) and a disturbance of photosynthesis (Islam et al., 2008). All correlation relationships for Pb concentrations within shoot and leaves from a variety of plants were positive. The increasing concentrations of Pb within roots also correspond to increasing concentrations within the shoots (Alia et al., 2015). Most of the Pb ions absorbed by plants from the soil remain concentrated in the roots, and a small portion is transferred to the stems and leaves (Zhou et al., 2018).
Soil acidification
In corn, spinach, and cabbage, in the treatments where urea (T2, T3 and T4) was applied, the soil was slightly alkalized during the first 7 d and then temporarily acidified (Figs. 1-5). This is because when urea is applied to the soil, it has two behaviors, a basic behavior during the first days when it goes from amide to ammonium carbonate, and an acidic behavior when it passes from the ammoniacal to the nitric form (releasing H+ to the medium); therefore, urea behavior is acidic at the end (FERTIBERIA, 2002). However, the acidification of the soil can be long-lasting or temporary, and the degree or intensity with which it varies is due to the continuous applications of N fertilizers such spinach (Spinacia oleracea L.) and cabbage (Brassica oleracea L.) as urea (Baldoncini, 2015). In our research, acidification was temporary because the dose of urea was only applied once. Additionally, this temporality had a direct relationship with the dose of urea, and the greater the dose of urea, the longer the soil remains acidic (Boccolini, 2016). Paschalidis et al. (2013) observed that the application of Pb significantly increases the total N content in the soil compared to the control treatment, perhaps because of the application of the metal in the nitrate form. In our research, the contaminant used was Pb acetate. The availability of Pb for uptake by plants is generally lower at high pH than at low pH (Paschalidis et al., 2013).
Maximum permissible limit (MPL) and bioconcentration factor (BCF) of Pb
In corn, spinach and cabbage, all treatments including the control exceeded the MPL for Pb (Tab. 10) of the Codex Alimentarius (0.05 mg kg-1 for corn and 0.3 mg kg-1 for cabbage) and other countries (0.1 mg kg-1 for spinach for the European Union, Australia and New Zealand, 0.5 mg kg-1 for Brazil, and 5 mg kg-1 for Japan). It should be noted that, in the control, Pb was not applied in the form of Pb acetate. Still, the soil already contained 0.026 mg kg-1 (a concentration lower than the MPL for Pb in agricultural land for Peruvian legislation) in the soil of the farm that was used to make the greenhouse substrate.
The possible source for the high levels of Pb, even in the controls, could be related to some possible variables not evaluated in this research that could be irrigation water (although it was demineralized water), the concentration of Pb in the soil at 93 d of exposure, or even atmospheric contamination and foliar deposition of Pb; however, it is not certain which of the factors listed was the main source of Pb in the leaves of the crops (Natasha et al., 2020). Some researchers point out that Pb is present in plants under natural conditions (Tóth et al., 2014). These results can be observed in the crops where agrochemicals, organic fertilizers, and pesticides are frequently used (Pesca, 2015). This overdose in both the amount and frequency of application (Torres and Ramírez, 2014) increases the probability of transfer or absorption of heavy metals in food, soil, and water (CONPES, 2014). In Peru, research has been carried out where it is evident that foodstuffs such as potatoes (Luna and Rodríguez, 2016) and eggs (González, 2015) exceed the MPL for Pb in food. Therefore, the spinach analyzed in this research is not suitable for consumption and poses a real risk of Pb exposure in the human body.
The BCF of Pb is the best way to know the availability of important metals transferred from soil to grown vegetables (Ugulu et al., 2020). The BCF values were less than 1 in all the treatments for the three plant species, which indicated that they were not accumulators of heavy metals in the agroecosystem. Similarly, Pakchoi cabbage (Brassica campestris L.) shows BCF values of less than 1 for Pb with the influence of green waste compost (Liu et al., 2020).
Conclusions
We observed that Pb increased the dry weight in corn and decreased the fresh and dry weights and leaf area in spinach (T4). Likewise, urea increased the values of chlorophyll in corn, spinach, and cabbage, indicating that urea influences the absorption of Pb in the leaf structure in the three plant species under study.
We determined that spinach is the only plant species that absorbed by the roots and translocated more Pb to the shoot of the plant by N fertilization (T4). Spinach showed a greater Pb storage capacity in its leaves (10.57 mg kg-1 average in all treatments) compared to corn (5.51 mg kg-1 average in all treatments) and cabbage (5.64 mg kg-1 in all treatments).
In corn and spinach, we determined that there was a direct relationship between the dose of urea and the acidity time of the soil, and an indirect relationship between the dose of urea and the pH value, which means that urea influenced soil acidification.
The idiosyncratic nature of the crop responses to the combined effects of Pb and N highlights the need for more research on the subject.

