











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkIntroduction
The growing demand for food in the world has led to an increase in the use of water bodies, depleting or contaminating rivers, lakes and surface waters, making access to drinking water more difficult, especially for human consumption (Santos & Spolador, 2022). This fact leads to restrictions on the use of drinking water in agriculture, increasing the use of water of inferior quality, such as saline water, in irrigation. Thus, the difficulty in accessing high quality water has accelerated the search for viable alternatives for food production (Zhang & Shen, 2019; Singh, 2021).
For the production of vegetables, the continuous use of soil and the high number of production cycles can lead to an increase in salt concentrations in the soil (Libutti & Monteleone, 2018). Also, wastewater from agro-industrial processes or low-quality water from other sources has been increasingly frequent in agriculture. This action results in significant productivity losses due to the presence of residues that can limit the development of plants, including lettuce (Adhikari et al., 2019; Zhang et al., 2021).
Irrigation with saline water is possible for many plant species, which have different levels of tolerance to this condition. However, many commercially important species have low salinity tolerance, and excess salts can lead to plant death (Hajihashemi et al., 2020). For these species, reduced fertility, changes in the physical and chemical characteristics of soils, in addition to the accumulation of Na both in the plants and in the soil, reduce germinative efficiency and promote physiological disorders, affecting the photosynthetic system, cell division and, consequently, the development of plant structures (Singh, 2021).
The impact of excessive use of soil and water resources can be reduced by applying alternative solutions that allow maintaining development without further damage to natural resources. The exogenous application of vitamins in plants can lead to increase in vigor, higher reproductive rates, and resistance to edaphoclimatic conditions (Contieri et al., 2018; Vendruscolo et al., 2019), among other physiological increments that allow cultivation under conditions of increased stress.
Thiamine, known as vitamin Bl, has been used as a potential biostimulant in plant production (Vendruscolo et al., 2019; Vendruscolo & Seleguini, 2020; Jabeen et al., 2021). This vitamin acts directly on plant organelles as an enzymatic cofactor and indirectly on the respiratory and energy generation cycles of plants (Goyer, 2010; Taiz et al., 2017). The application of exogenous thiamine can stimulate the action of defense mechanisms in plants, in addition to promoting development and growth (Kaya et al., 2015).
Another substance with application to crop systems is niacin (vitamin B3), which is also found in several physiological systems of plants. It helps in vegetative growth and accumulation of reserves and minimizes adversities that the plant may undergo from stress (Taiz et al., 2017; Colla et al, 2021).
Based on the hypothesis that the application of the В vitamins, niacin and thiamine can change the characteristics of vegetables and mitigate the effects of abiotic stresses, the objective of the study was to evaluate the effects of the application of vitamins В on lettuce plants submitted to irrigation with saline water.
Materials and methods
Plant material and treatments
The experiment was conducted in a greenhouse covered with a 150-micron low-density polyethylene film and a thermoreflective screen (LuxNet®) with 42/50% shading under the film, in April 2021, in the experimental area of the State University of Mato Grosso do Sul, Cassilândia University Unit, Brazil.
The climate of the region, according to the Koppen classification, is tropical rainy (Aw), with rainy summers and dry winters (winter precipitation less than 60 mm), with annual precipitation of 1,520 mm and an average temperature of 24. FC. During the experimental period, the climatic data were collected daily using equipment installed inside the greenhouse. (Fig. 1).

A completely randomized design was used, in a 2x4 factorial scheme, with four replicates. The treatments were composed of two lettuce cultivars (Valentina and Pira Roxa) and the four growing conditions, Control: irrigated with water and without application of vitamins; NaCl: irrigated with saline solution and without application of vitamins; NaCl+Bl: irrigated with saline solution and application of vitamin thiamine; and NaCl+B3: irrigated with saline solution and application of vitamin niacin.
The lettuce seedlings were purchased from a certified producer (Agromudas, Jales, Brazil), ensuring phytosanitary quality and homogeneity in size. Before planting in the containers, the seedlings received a foliar application of water (control) and solutions of thiamine (100 mg I/1) and niacin (100 mg I/1), depending on the treatment. The vitamin solutions were prepared in water, and the treatments were applied with a foliar spray, using a plastic hand pump. One ml of solution was applied per plant and per treatment. After 24 h of application, the plants, containing four leaves and 15 cm high, were transferred to the containers with substrate. One treatment without application of vitamins was irrigated with fresh water (0.48 dS m-1) (control), while another treatment without vitamins was irrigated with saline solution (50 mM NaCl, 5.18 dS m-1), used as "stress control" (NaCl); the other treatments containing thiamine (NaCl+B1) and niacin (NaCl+B3) application were irrigated with saline solution for 15 d. Four replicates were used, consisting of two plants each.
For plant growth, recipients (0.35 dm3) contained a mixture of Typic Quartzipsamments (pH 5.0, P 14.0 mg dm-3, K 3.0 mmolc dm-3, Ca 24.0 mmolc dm-3, Mg 14.0 mmolc dm-3, cation exchange capacity 58.0 mmolc dm-3, base saturation 71.0%, organic matter 13.0 g dm-3, 95 g kg-1 of clay, 50 g kg-1 of silt and 855 g kg-1 of sand), vermiculite and tanned bovine manure, in a proportion of 3:1:1 (v/v).
Gas exchange and plant weight accumulation
Fourteen days after the application of the vitamins, during the morning hours, the net photosynthesis (A; μmol CO2 m-2 s-1), stomatal conductance (gs; mmol rn2 s-1), intracellular CO2 concentration (Ci; μmol mol-1) and transpiration (E; mol H2O m-2 s-1) were evaluated, using a portable infrared gas exchange meter (LCi, ADC Bioscientific, Hertfordshire, UK). Also, water use efficiency (A/E) and the instantaneous carboxylation efficiency (A/Ci) were calculated.
After obtaining the physiological characteristics, the aerial part of the plants was harvested and dried in a forced ventilation oven at 65°C, until a constant dry mass (SDW) was obtained. Also, the percentage loss of dry mass was calculated based on the control treatment.
Statistical analysis
Data were submitted to preliminary normality and ho-moscedasticity tests. Then, the means were submitted to analysis of variance (ANOVA) and the Scott-Knott test, at the 10% probability level. The analysis was performed using the SISVAR statistical software (Ferreira et al., 2014).
Results
The treatments affected the net photosynthesis variable, with the highest average obtained in the control treatment, followed by the treatments composed of the application of vitamins B1 and B3, which differed significantly from the treatment composed only by irrigation with saline solution (Fig. 2A). Differences in the intracellular concentration of CO2 only occurred between the cultivars, with Pira Roxa being superior in this variable. In addition, for both transpiration and stomatal conductance, the superiority of the control treatment over the others was observed, and of the cultivar Pira Roxa over Valentina, without considering the treatments (Fig. 2C-D).
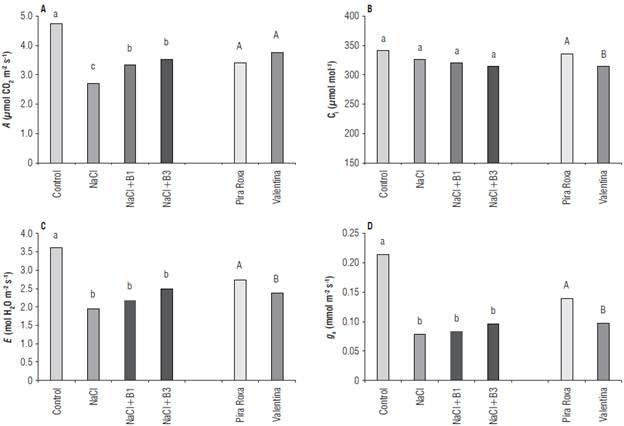
FIGURE 2 A) A - net photosynthesis; В) C¡ - intracellular C02 concentration; C) E- transpiration, and D) g s - stomatal conductance in lettuce plants submitted to foliar application with vitamins and irrigation with saline solution. Bars represent means; n=4. Control: irrigated with water and without application of vitamins; NaCl: irrigated with saline solution and without application of vitamins; NaCI+B1: irrigated with saline solution and application of vitamin thiamine; NaCI+B3: irrigated with saline solution and application of vitamin niacin. Lowercase letters compare treatment means and uppercase letters compare cultivar means.
For water use efficiency, higher values were found for treatments in which irrigation with saline solution was used and, among the cultivars, Valentina was superior (Fig. 3A). In addition, for the efficiency of instant carboxylation, the control treatment stood out, followed by the treatments composed by the application of vitamins, both superior to the treatment composed only by irrigation with saline solution, while among the cultivars Valentina stood out (Fig. 3B).
The cultivars had a similar response for the accumulation of dry mass, while, among the treatments, the superiority of the control was verified. However, among the treatments in which there was irrigation with saline solution, the superiority of the application of thiamine was observed (Fig. 4A), with lower percentage losses of dry mass (Fig. 4B).

FIGURE 3 (A) Water use efficiency and (B) instantaneous carboxylation efficiency in lettuce plants submitted to foliar application with vitamins and irrigation with saline solution. Bars represent means; n=4. Control: irrigated with water and without application of vitamins; NaCl: irrigated with saline solution and without application of vitamins; NaCI+B1 : irrigated with saline solution and application of vitamin thiamine; NaCI+B3: irrigated with saline solution and application of vitamin niacin. Lowercase letters compare treatment means and uppercase letters compare cultivar means.

FIGURE 4 (A) Shoot dry mass and (B) average values of dry mass loss In lettuce plants submitted to foliar application with vitamins and irrigation with saline solution. Bars represent means; n=4. Control: irrigated with water and without application of vitamins; NaCl: irrigated with saline solution and without application of vitamins; NaCI+B1: irrigated with saline solution and application of vitamin thiamine; NaCI+B3: irrigated with saline solution and application of vitamin niacin. Lowercase letters compare treatment means and uppercase letters compare cultivar means.
Discussion
The presence of salt concentrations in the irrigation water affected most of the parameters evaluated compared to the control treatment (Figs. 2-4); the resulting osmotic stress caused decreases in leaf area, transpiration, osmotic potential, dry biomass, and photosynthetic rate (Coelho et al., 2013; Silva Junior et al., 2018); the application of vitamins attenuated some of these effects (Figs. 2-4).
Thiamine is responsible for signaling the presence of stress acting on plants; its natural levels in plant tissues decrease as stress continues (Goyer, 2010). Thiamine has key roles in plant metabolic processes, including carbon assimilation and respiratory processes (Fitzpatrick & Chapman, 2020). The exogenous application of thiamine acts as priming, potentiating the responses to stress through the production of secondary metabolites and favoring the plant self-protection (Goyer, 2010; Kaya et al., 2015). The treatments with this vitamin showed lower losses of dry mass (Fig. 4), which may be an indication of the plant self-protection in response to stress. As well, the thiamine treatments with saline water irrigation led to a positive response for net photosynthesis (Fig. 2A).
When the plant is subjected to stress, the application of thiamine results in increases in glycine-betaine, the total levels of phenols, the activity of catalase and peroxidase enzymes and the levels of proline, which directly protect the plants by nullifying reactive oxygen species and helping to maintain photosynthetic pigments (Jabeen et al, 2021).
The application of thiamine also increases energy reserves (sugars) and nutritional reserves in plant tissues (Kaya et al., 2015). This increases plant resilience against the deleterious effects exerted by stress on the growing environment (Taiz et al, 2017).
The positive effect observed for the application of niacin on photosynthetic activity is related to its participation as a constituent of NAD+ and NADP+, acting directly in the transport of electrons in cellular and respiratory metabolism (Meyer-Ficca & Kirkland, 2016), with increases of the pigment photosynthetic agents and protection of the lipid layer in membranes of the leaf cells (Hussein et al, 2014). However, despite the positive effect reported in the literature, the application of niacin did little to alleviate the effects of salinity in the present study. This maybe related to the composition of lettuce plants, which might have high levels of niacin. Thus, the exogenous application of this vitamin may not have been sufficient to result in significant changes in the plants. The application of niacin to a curly mustard crop at concentrations close to 485.20 mg L-1 did increase plant development (Vendruscolo et al, 2017).
Other authors observed that the application of thiamine at different concentrations effectively affects and favors the development of rice plants (Vendruscolo et al, 2019) and sweet corn (Vendruscolo et al, 2018), especially when the plants are subjected to abiotic stresses or field production conditions, where environmental factors act on the plants. Additionally, the application of niacin has a biostimulant effect, promoting the development and productivity of species such as corn (Colla et al, 2021) and beans (Abreu et al, 2020) in the field.
Conclusions
The exogenous application of vitamins helps to mitigate the effects caused by irrigation with saline water on lettuce plants, reducing stress on photosynthetic mechanisms and increasing photosynthetic activity. In addition, thiamine helps to reduce the deleterious effects of salinity on the dry mass accumulation in plants.

