Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBoletín Científico. Centro de Museos. Museo de Historia Natural
Print version ISSN 0123-3068
Bol. Cient. Mus. Hist. Nat. Univ. Caldas vol.12 no.1 Manizales Jan./Dec. 2008
William Rojas Vinasco1
Jaime V. Estévez-Varón1
Néstor Roncancio1,2
*Recibido 25 de febrero de 2008, aceptado 1 de julio de 2008.
1Grupo de Investigación en Ecosistemas Tropicales, Universidad de Caldas.
2Maestría en Ciencias – Biología, Universidad Nacional de Colombia, sede Bogotá.
Se estudio la estructura y composición florística de cinco remanentes de bosque húmedo tropical de la vertiente norte del río Moro y San Antonio en el municipio de Samaná, (Caldas, Colombia). Se censaron todos los individuos con DAP > 2,5 cm en 0,08 ha por sitio. Se registraron 994 individuos pertenecientes a 123 morfoespecies, 91 géneros y 42 familias de plantas leñosas. La familia con mayor riqueza de especies fue Euphorbiaceae (11 especies), seguido de Fabaceae (10), Melastomataceae (8), Moraceae (8) y Rubiaceae (7). Los remanentes dos, cuatro, cinco y tres presentaron una composición florística similar. Se presentaron diferencias significativas en la estructura de la vegetación entre remanentes de bosque en la clase diamétrica DAP > 10 cm. Los resultados sugieren que tanto la estructura como la composición florística son afectadas por la perturbación natural o antrópica ya sea por la presencia de claros o por la tala selectiva.
Estructura, composición florística, perturbación, riqueza, bosque húmedo tropical.
The floristic structure and composition of five tropical humid forest remainders of the north basin of the Moro and San Antonio rivers (Samana, Caldas, Colombia) were studied. All the individual plants with DAP > 2.5 cm in 0.08 ha were sensed, resulting in 994 individual belonging to 123 morphospecies, 91 genera and 42 families of woody plants. The Euphorbiaceae family presented the highest richness of species (11 species), followed by Fabaceae (10), Melastomataceae (8), Moraceae (8) and Rubiaceae (7). The remainders two, four, five and three showed a similarity in their floristic composition. Significant differences were seen in the floristic structure among the forest remainders in the diametric class DAP > 10 cm. The results suggest that the floristic structure, as well as the composition, is influenced by natural or human disturbances, either due to the presence of gaps, or due to selective cutting of trees.
Structure, floristic composition, disturbance, richness, tropical humid forest.
INTRODUCCIÓNLos bosques húmedos tropicales (bh-T) representan casi un 25% de la superficie total de bosques en el mundo. América tropical posee alrededor del 40%, Asia el 32% y África 10% (BUNDESTAG, 1990). Estos bosques se encuentran entre las latitudes 10°N y 10°S, en regiones donde la precipitación media anual es superíor a los 1000 mm (FAO, 1993).
Colombia con una extensión de 31'057.800 ha de bosque húmedo tropical, ha perdido la tercera parte de éstos por debajo de 1000 m (ESPINAL & MONTENEGRO, 1963; ETTER, 1993). Una de las principales causas de este fenómeno es la transformación del bosque por actividades humanas como la extracción de madera, la ampliación de la frontera agrícola (KATTAN, 1997), estos factores junto con el desarrollo de actividades ilegales como cultivos ilícitos vinculados al narcotráfico propician la desaparición de estos ecosistemas (VÉLEZ, 1990).
La transformación de los bosques, en su gran mayoría es parcial, lo que resulta en la creación de paisajes fragmentados en donde su composición con respecto al bosque original es diferente (KATTAN, 1997; TABARELLI et al., 1999). En vista de que la fragmentación incide en la desaparición de muchas especies arbóreas, ya sea por factores bióticos como limitaciones en la polinización y diseminación de semillas por animales o abióticos como cambios en la temperatura y la humedad relativa, ésta con el tiempo da lugar a una reducción en la riqueza de especies (ASQUITH, 2001).
La caracterización local de la vegetación representa el primer paso hacia el entendimiento de la estructura y dinámica de un bosque (BAWA & MCDADE, 1994), también es una herramienta importante para el manejo a largo plazo de áreas reducidas o fragmentadas, de las cuales en su mayoría no se cuenta con información necesaria para revertir estos procesos (CASCANTE & ESTRADA, 2001); por lo expuesto anteriormente, se evidencia la necesidad de realizar estudios que conduzcan al conocimiento de la diversidad y riqueza de estos ecosistemas (REDFORD et al., 1990), por tal razón el presente estudio tiene como objetivo determinar la estructura y composición florística de remanentes de bosque húmedo tropical en la cuenca del Río Moro en el oriente de Caldas.
MATERIALES Y MÉTODOSÁrea de estudio: la investigación se realizó en la vereda La Primavera, jurisdicción del corregimiento de Florencia, municipio de Samaná, departamento Caldas, Colombia, en las coordenadas 5030´29" N, 74059´18" W. La zona de estudio se encuentra en la vertiente oriental de la Cordillera Central en un sector aledaño a los terrenos de inundación del embalse Amaní sobre el Río Moro y corresponde a una zona de vida bosque húmedo tropical (bh-T) según la clasificación de HOLDRIDGE (1979).
Método de muestreo: el estudio se llevó acabo durante los meses de octubre y noviembre de 2005. Se realizó un muestreo estratificado preferencial (MATTEUCCI & COLMA, 1982), el cual presenta buenos resultados para evaluar la riqueza de especies en paisajes rurales de los Andes Centrales de Colombia (MENDOZA et al., 2005). Se seleccionaron cinco remanentes de bosque, el remanente uno (R1) tiene 6 ha, el remanente dos (R2) 8,2 ha, el remanente tres (R3) 5,5 ha, el remanente cuatro (R4) 7,1 ha, y el remanente cinco (R5) con un área de 11,2 ha.
En cada remanente se establecieron dos unidades de muestreo de 400 m2 separadas 50 m. En cada unidad se establecieron dos transectos de 50 x 4 m separados 5 m, en ellas se evaluaron las especies con DAP > 5 cm. Para evaluar los individuos de DAP entre 2,5 y 5 cm, se realizaron cuatro transectos de 50 x 1 m al interíor de cada transecto de 50 x 4 m. MENDOZA et al., 2005.
Se censaron todas las plantas leñosas de crecimiento libre (sin incluir epifitas, hierbas y lianas) que se encontraban con al menos el 50% de su tronco enraizado dentro del transecto. A cada individuo se le midió el DAP a 1,3 m, altura y diámetro de copa, así como la presencia de exudado, olores, sabores, colores y descripción de la corteza. De cada individuo se colectaron tres muestras, las cuales se etiquetaron con un número consecutivo. Las muestras se conservaron en alcohol al 70%. Se transportaron al herbario de la Universidad de Caldas (FAUC) –Colombia–, donde fueron secadas, saneadas e identificadas hasta género y en lo posible especie por comparación con muestras del mismo o mediante el empleo de claves para familias, géneros y especies de árboles, o se recurrió a la asesoría de especialistas.
Análisis de datos: para la evaluación de la estructura vegetal de cada remanente, se calculó el área basal, el valor de importancia para familias (IVF), el valor de importancia para especies (IVI), y área de copa sólo para individuos con DAP > 10 cm (MATTEUCCI & COLMA, 1982; MORÍ & BOOM, 1987). Para analizar las diferencias en la estructura vegetal de todos los remanentes se comparo el área basal, la altura y el área de copa por clases diamétricas. Estas últimas se establecieron mediante una modificación de las propuestas por el Instituto Humboldt para evaluar la riqueza de especies en paisajes rurales (MENDOZA et al., 2005). Este método consiste en dividir los árboles en tres clases: individuos con DAP entre 2,5 y 4,99 cm, individuos con DAP entre 5 y 9,99 cm e individuos con DAP > a 10 cm. A estos datos se les realizó un análisis de varianza de una vía, previa comprobación del supuesto de normalidad y homogeneidad de varianzas. Para datos que no cumplían los supuestos se realizó la prueba de Kruskall-Wallis (K-W). En los casos donde se presentaron diferencias se realizó un prueba de Tukey o Nemenyi para determinar entre qué remanentes existían diferencias (ZAR, 1999).
La estimación de la riqueza específica (S), entre los remanentes se realizó mediante una curva de acumulación de especies, basada en la abundancia de individuos, con el estimador (ACE) (COLWELL & CODDINGTON, 1994; LEE AND CHAO, 1994; CHAZDON et al., 1998; COLWELL, 2000; MAGURRAN, 2004). Los datos se analizaron con el programa EstimateS 8.0 (COLWELL, 2000). Para determinar la similitud de las especies vegetales entre los remanentes de bosque se realizó un análisis de Clúster con base en abundancias. Éste se basó en la distancia Euclidiana con el algoritmo UPGMA (SNEATH & SOKAL, 1973).
RESULTADOS Riqueza Florística
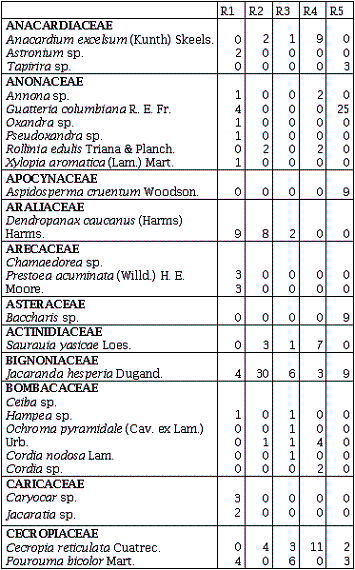
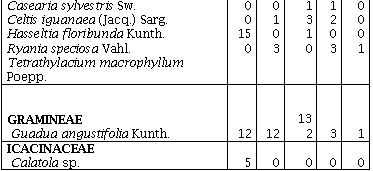
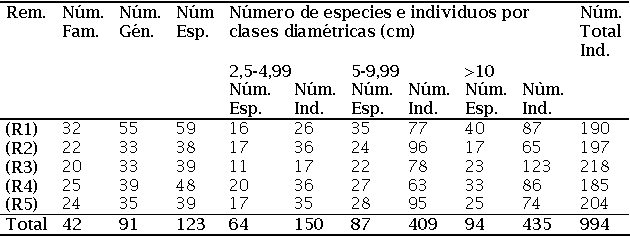
Se registraron 994 individuos leñosos, pertenecientes a 42 familias, 91 géneros y 123 morfoespecies. De ellas 77 se determinaron hasta especie (62,6%), y las restantes 46 fueron determinadas hasta género (37,4%). De las 42 familias encontradas, 41corresponden a la clase Magnoliopsida, representada por 122 especies (99,2%), y sólo se registró una familia con una especie (0,8%) perteneciente a la clase Liliopsida, esta especie es Guadua angustifolia Kunth (Gramineae).
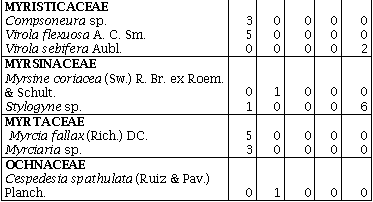
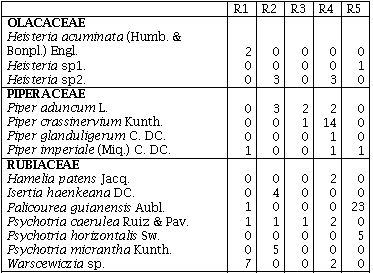
La familia Annonaceae presentó la mayor cantidad de géneros (5) en el R1, seguida por Flacourtiaceae con cuatro géneros en R3 y R4 y Euphorbiaceae con cuatro géneros en R1, R2 y R4. La familia Melastomataceae en los R2 y R3 y Euphorbiaceae en el R4, presentaron el mayor número de especies (6), seguidas por Annonaceae con cinco especies en el R1 (Anexo 1). El mayor número de especies (59) para un sitio se presentó en el R1 distribuidas en 33 familias y 55 géneros, mientras el menor número se presentó en el R2 con 38 distribuidas en 22 familias y 33 géneros (Tabla 1).
En las curvas de acumulación de especies (Figura 2) se puede observar que el cambio en la riqueza de especies incrementa con el número de individuos muestreados, pero que el tamaño de la muestra de 0,08 ha por sitio, no fue suficiente para lograr la asíntota.
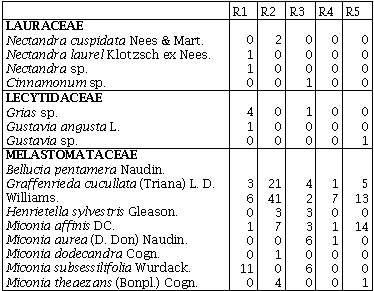
Los remanentes de bosque húmedo tropical comparten sólo 10 familias de las 42 encontradas, y 11 familias sólo se encuentran en uno de los cinco remanentes. En cuanto a las especies sólo seis (4,9%) se encuentran presentes en los cinco remanentes de bosque: Jacaranda hesperia Dugand; Casearia arborea (Rich.) Urb.; Guadua angustifolia Kunth; Bellucia pentamera Naudin; Graffenrieda cucullata (Triana) L. D. Williams, y Miconia affinis DC. En el R1 se encontraron 25 especies exclusivas, ocho en el R2, seis en el R3, en el R4 y R5 doce especies fueron exclusivas (Anexo 1).
Los remanentes R2, R3, R4 y R5, fueron similares en la composición florística. Los remanentes R4 y R5 comparten 14 especies; los remanentes R2 y R3, 17 especies, mientras que el R1 presentó la menor similitud con respecto a los otros sitios (Figura 1).


Estructura
Las comparaciones de las variables estructurales (análisis de varianza) entre los cinco remanentes de bosque presentaron diferencias significativas para las clases diamétricas con DAP entre 2,5-4,99 cm, para la variable altura en metros (F=2,11; p=0,0406). Para árboles con DAP > 10 cm, se presentó diferencias significativas con el test de Kruskal-Wallis (K-W) para las tres variables: área basal en m2 (K-W; p=0,00001329), altura en metros (K-W; p=0,000) y área de copa en m2 (K-W; p=0,0014).
En la comparación entre sitios (Test de Tukey-Nemenyi) para las variables estructurales que presentaron diferencias significativas con la prueba de Kruskal-Wallis, se presentaron diferencias entre el R3 y los demás sitios para la variable altura (m) en individuos con DAP > 10 cm, y entre el R4 vs. el R3 y el R1 para individuos con DAP < 5 cm. La variable área basal en m2 presentó diferencias entre el R3 vs. el R1 y el R4. Entre el R1 y el R5 se presentó diferencias para la variable área de copa en m2.
Los individuos que presentaron mayor altura fueron: Jacaranda hesperia Dugand en R1 con 30 m, en el R2 y R3 con 29 m; Tetrorchidium rubrivenium Poepp en el R2 con 30 m; Inga tenuistipula Ducke en el R1 con 28 m. En el R4 todos los individuos presentaron una altura inferior a 24 m, y la especie con mayor altura fue Cecropia reticulata Cuatrec., con 23,5 m. El R5, presentó las menores alturas, la especie más alta fue Himatanthus sp. con 19,5 m.
El mayor número de especies (40) para una clase diamétrica (árboles con DAP > 10 cm) se presentó en el R1, mientras el menor se presentó en el R2 con 17. El mayor número de individuos (123) para una clase diamétrica se presentó en el R3 (Tabla 1). Las especies que presentaron mayor diámetro a la altura del pecho (DAP) fueron: Jacaranda hesperia Dugand con 68,8 cm en el R1; Turpinia occidentalis (Sw.) G. Don con 65,2 cm en el R4; Albizia niopoides (Spruce ex Benth.) Burkart, con 62,5 cm en el R4; Erytrina poeppigiana (Wale) O.F. Cook con 59,8 cm en el R4, y en el R5 se presentaron los individuos con los menores DAP, y la especie que presentó el mayor diámetro fue Maquira sp., con 43,8 cm.
La mayor área basal la presentó el R1 con 5 m2 y la menor fue para los R2 y R5 ambos con 2,5 m2, mientras R3 y R4 presentaron valores de 3 y 4 m2, respectivamente. A nivel de especies se destacaron Guadua angustifolia Kunth con el 50,2% de dominancia relativa en el R3, seguida de Graffenrieda cucullata (Triana) L.D. Williams con el 19% en el R2 y Jacaranda hesperia Dugand con el 14,2% del área en el R1, 16,9% en el R2, 13,8% en el R3, y 10,4% en el R5.
La mayor cobertura de copa la presentó el R1 y la especie Trophis sp. (Moraceae) con 264 m2 presentó la mayor cobertura de copa para un sólo individuo en el mismo remanente, seguida de Inga acuminata Benth (Fabaceae) con 216 m2 en el R4, Hura crepitans L. (Euphorbiaceae) con 214 m2 también en el R4, Caryocar sp. (Caryocaraceae) con 148 m2 y Pourouma bicolor Mart. (Cecropiaceae) con 121,6 m2, ambas en el R1; en el R5 se presentaron los individuos con menor cobertura de copa y la especie con mayor cobertura fue Vismia lauriformes (Lam.) Choisy (Clusiaceae) con 48,3 m2.
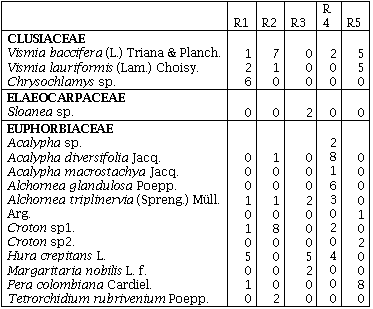
Índice de Valor de Familias (IVF)Las familias de mayor importancia ecológica fueron: Gramineae con un IVF 113,3 (37,9%) en el R3, seguida de Melastomataceae con un IVF de 84,3 (28,2 %) en el R2 y 42 (14%) en el R5. Euphorbiaceae fue la familia que presentó el mayor número de especies en el R4 con un IVF de 45, obtuvo el 15% de importancia para este sitio. La familia Melastomataceae en todos los sitios presentó los géneros: Miconia, Bellucia, Graffienrienda, mientras que Henrietella sólo se presentó en los R2 y R3.
Índice de Valor de Importancia para cada especie (IVI)El mayor valor de importancia ecológica lo presentó Guadua angustifolia Kunth en el R3 a 650 msnm con un valor de 115 (38,3%). Seguida de Graffenrieda cucullata (Triana) L.D. Williams con 44,3 (14,8%) y Jacaranda hesperia Dugand con 36,5 (12,3%) ambas en el R2, y 20,8 en el R3. Otras especies importantes fueron Bellucia pentamera Naudin; Croton sp1. y Maquira sp. (Tabla 2).

DISCUSIÓN
Riqueza Florística
Las familias y géneros de plantas con mayor número de especies encontradas en los cinco remanentes son las mismas reportadas por GENTRY (1982a) en tierras bajas triopicales, por BALCAZAR et al., (2000) en la Serranía de Las Quinchas Magdalena medio, entre 380 y 1000 m, y por CUATRECASAS (1958) en gradientes cordilleranos andinos. Esto puede ser consistente con la noción de que diferentes especies de estas familias se adaptan a diferentes condiciones ecológicas, o que se dispersaron de un área remota, seguida de una amplia radiación adaptativa en ausencia de posibles competidores (WHITTAKER, 1956; WHITMORE, 1975; GENTRY, 1982a, 1982b, 1988; TEBORGH et al., 1996; VÁZQUEZ & GIVNISH, 1998).
La variable número de especies encontradas por género para los cinco remanentes de bosque fue baja, en promedio 1,5 ± 0,3 para cada una de las familias más ricas, este resultado contrasta con el patrón observado en muchos bosques lluviosos tropicales de tierras bajas, en que pocos géneros (como inga, Piper, Psychotria) contribuyen sustancialmente para la riqueza de especies (GENTRY, 1988; VÁZQUEZ & GIVNISH, 1998). Esta variación se puede explicar por las variaciones en el hábitat y las perturbaciones que, junto con los efectos naturales y las condiciones del suelo, controlan la diversidad de los árboles (RICHARDS, 1952; WHITMORE, 1998).
La mayor riqueza del remanente uno (R1) se puede explicar porque éste hace parte de la zona de amortiguación del embalse Amaní, por lo que presenta un mejor estado de conservación y la presión sobre este remanente disminuye caso contrario ocurre en los R2 y R5 donde existe la presencia de caminos muy transitados o en el R2 donde se observó la tala selectiva. Esta disminución de la riqueza florística en los remanentes estudiados es similar a lo sugerido por LOVETT et al. (2006) donde esta disminuye por la presencia de viviendas cerca a bosques donde sus habitantes utilizan el bosque como recurso.
La presión humana que ha existido y que existe por los recursos maderables de estos sitios se comprobó, al encontrarse una baja diversidad de especies en las familias de importancia económica como Lauráceae y Meliaceae, ya que sólo se presentaron con una especie, en uno de los cinco remanentes de bosque (Anexo 1), esta misma presión sobre el recurso bosque puede ser la responsable de la alta similaridad de los remanentes R2, R4, R5 y R3 (Figura 1).
EstructuraLas especies que presentaron los mayores valores del IVI (Tabla 2) se encuentran entre las especies más abundantes presentes para todos los sitios (Anexo 1). De ellas Jacaranda hesperia Dugand, Bellucia pentamera Naudin y Miconia affinis DC se encuentran entre las especies reportadas por BALCÁZAR et al. (2000) para la Serranía de Las Quinchas, Magdalena medio, Colombia, entre 380 y 1000 m. La existencia de estas especies dominantes puede reflejar tendencia aleatoria sobre la evolución del control denso dependiente para enemigos naturales especialistas, lo que puede acontecer para la distribución observada de la abundancia de árboles y para la dominancia reiterada de taxas particulares en sitios separados pero similares ecológicamente (GIVINHS, 1999).
Las diferencias significativas (Test de Kruskal-Wallis; p=0,05) entre sitios para individuos con DAP > a 10 cm, nos sugieren que estos sitios se encuentran en fases de regeneración natural con desarrollos intermedios influenciados por disturbios naturales o humanos, ya que las diferencias en el área basal son características de sitios que presentan algún grado de regeneración natural provocado por algún tipo de perturbación como la caída de arboles (obs. pers.) ya que el incremento en área basal en muestras de tamaño similar refleja la capacidad potencial de los individuos para llenar el área superficial o espacio disponible en el bosque (ENQUIST & NIKLAS, 2001).
Las diferencias significativas en la altura de los árboles con DAP < a 5cm y > 10 cm, nos sugieren variaciones en el grado de las perturbaciones tanto naturales como antropogénicas (RICHARDS, 1952), que a pequeña escala causan vacíos en el dosel y alteran la estructura del bosque (HARTSHORN, 1978, 1980; WHITMORE, 1978), es el caso de perturbaciones causadas por caminos muy transitados (R2 y R5) o la tala selectiva en el remanente dos (R2), estas perturbaciones pueden ser un factor en la estructura de las comunidades (SUMINA, 1994; BURSLEM & WHITMORE, 1999; HUBBELL et al., 1999; SHEIL & BURSLEM, 2003).
Las diferencias estructurales entre los remanentes de bosque, también podrían deberse, en parte, a las actividades que demandan la extracción de madera como la construcción de cercas, viviendas, el procesamiento de la panela y la cocción de los alimentos. Estas actividades pueden propiciar que las especies extraídas, presenten un menor DAP y altura que las especies no utilizadas de menor valor maderable como individuos de Bombacaceae, Moraceae, Fabaceae, Bignoniaceae y Caryocaraceae, los cuales presentaron mayor altura, DAP y cobertura de copa en los remanentes de bosque. Lo anterior comprueba el hecho de que especies poco comercializadas, o de bajo valor, tiendan a mantenerse como emergentes, además de presentar baja abundancia, alta dominancia y grandes dimensiones en sitios perturbados (VALVERDE, 1998).
AGRADECIMIENTOSEste proyecto fue realizado con recursos del convenio C 099-2004 entre CORPOCALDAS-Universidad de Caldas. Los autores manifiestan su agradecimiento a Oscar Ospina de CORPOCALDAS por el apoyo brindado para el desarrollo del proyecto. A Luis Miguel Álvarez por sus asesorías en la identificación de las muestras y el préstamo de las instalaciones del herbario de la Facultad de Agronomía de la Universidad de Caldas (FAUC). Y a todas las personas de la vereda La Primavera por su ayuda y hospitalidad. A Gabriel Jaime Castaño Villa, Martha Sofía González Insuasti y María Fabiola Ospina Bautista por las sugerencias realizadas al manuscrito.
BIBLIOGRAFÍA ASQUITH, N. M., 2001. La dinámica del bosque y la diversidad arbórea: 337- 406 (en) Libro universitarío regional (1 ed.): Ecología y Conservación de Bosques Neotropicales. Instituto Tecnológico. Costa Rica. [ Links ]
BALCÁZAR, V. M. P., RANGEL, CH. J. O. & LINARES, C. E. L., 2000.- Diversidad florística de la serranía de Las Quinchas, Magdalena medio (Colombia). Caldasia, 22 (2): 191-224. [ Links ]
BAWA, K. S. & MCDADE, L., 1994.- The plant community: Composition, dynamics, and life-history processes-commentary: 68 p. (en) L. MCDADE, K. S. BAWA, H. A. HESPENHEIDE & G. S. HARTSHORN (eds.) La Selva: Ecology and Natural History of a Neotropical Rain Forest. The University of Chicago, Chicago, Illinois. [ Links ]
BUNDESTAG, G., 1990.- Protecting the tropical forests: A high-priority international task. Second report of the enquete commission "Preventive Measures to Protect the Earth's Atmosphere" of the XI German Bundestag. Bonn. [ Links ]
BURSLEM, D. F. R. P. & WHITMORE, T. C., 1999.- Species diversity susceptibility to disturbance and tree population dynamics in tropical rain forest. J. Veg. Sci., 10: 767-776. [ Links ]
CASCANTE, M. A. & ESTRADA A. CH., 2001.- Composición florística de un bosque húmedo premontano en el valle central de Costa Rica. Rev. Bio. Trop., 49 (1): 213-225. [ Links ]
CHAZDON, R. L., COWELL, R. D. DENSLOW, J. S., GUARIGUATTA, M. R., 1998.- Statistical methods for stimating species richness of woody regenertation in primary and secondary rain forest of northeastern Costa Rica: 285-309 (in) DALLMEIER, F., COMINSKY, J. A. (eds.) Forest Biodiversity Research, Monitoring and Modeling Conceptual Background and Old World Cases Studies. Parthenon publishing, Paris. [ Links ]
COLWELL, R. K. & CODDINGTON, J. A., 1994.- "Estimating terrestrial biodiversity through extrapolation". Philosophical Transaction of the Royal Society of London. Series B, 345: 101-118. [ Links ]
COLWELL, R. K., 2000.- Estimates statistical estimation of species richness and shared species from samples. User's guide and application, http://viceroy.eeb.uconn.edu/estimates. [ Links ]
CUATRECASAS, J., 1958.- Aspectos de la vegetación natural de Colombia. Ex. Fis. Nat., 10 (40): 221- 268. [ Links ]
ENQUIST, B. J. & NIKLAS, K. J. N., 2001.- Invariant scaling relations across tree-dominated communities. Nature, 410: 655-660. [ Links ]
ESPINAL, T. S. & MONTENEGRO, M. E., 1963.- Formaciones vegetales de Colombia. Instituto Geográfico Agustín Codazzi. Bogotá, D.C. Colombia. [ Links ]
ETTER, A., 1993.- Diversidad ecosistémica en Colombia hoy: 43-61 (en) Nuestra Diversidad Biológica, CEREC, Fundación Alejandro Ángel Escobar, Bogotá, D. C. Colombia. [ Links ]
FAO, 1993.- The challenge of sustainable forest management. What Future for the World's forests FAO? Roma, 128 pp. [ Links ]
GENTRY, A. H., 1982a.- Patterns of neotropical plants species diversity. Evol. Biol., 15: 1-84. [ Links ]
GENTRY, A. H., 1982b.- Neotropical floristic diversity: Phytogeographical Connections between Central and South America. Pleistocene climatic fluctuations, or an accident of the Andean orogeny. Annals of the Missouri Bot. Garden, 69: 557-593. [ Links ]
GENTRY, A. H., 1988.- Changes in plant community diversity and floristic composition on environmental and geographical gradients. Ann. Missouri Bot. Garden, 75: 1-34. [ Links ]
HARTSHORN, G. S., 1978.- Tree falls and tropical forest dynamics: 617- 638 (en) By P. B. TOMLINSON & M. H ZIMMERMANN (eds.) Tropical Trees as living system. Cambridge University Press, Cambridge. [ Links ]
HARTSHORN, G. S., 1980.- Neotropical forest dynamics. Biotropica, 12 : 23-30. [ Links ]
HOLDRIDGE, L. R., 1979.- Ecología basada en zonas de vida. Instituto Interamericano de Ciencias Agrícolas, San José, Costa rica. [ Links ]
HUBBELL, S. P., FOSTER, R. B., O'BRIEN, S. T., HARMS, K. E., CONDIT, R. WECHSLER, B., WRIGHT, S. J. & LOODE LAO, S., 1999.- Light-gap disturbance, recruitment limitation, and tree diversity in a neotropical forest. Science, 283: 554-557. [ Links ]
KATTAN, G., 1997.- Transformación de paisajes y transformación de hábitats (en) MARÍA ELFI CHAVES & NATALIA ARANGO (eds.) Causas de Pérdida de Biodiversidad. Instituto de Investigaciones Biológicas Alexander Von Humboldt, Bogotá D.C. 76-87 pp. [ Links ]
LEE, S. M., CHAO, A., 1994.- Estimating population size via sample coverage for closed capture-recapture models. Biometrics, 50: 88-97. [ Links ]
MAGURRAN, A. E., 2004.- Measuring biological diversity, Backwell Science. Malden, MA, U.S.A. [ Links ]
MATTEUCCI, S. & COLMA, A., 1982.- Metodología para el estudio de la vegetación. Secretaría General de la Organización de Estados Americanos (OEA). Washington D.C., USA. [ Links ]
MENDOZA, J. E., JIMÉNEZ, F. H., LOZANO-ZAMBRANO, P., CAYCEDO-ROSALES & RENJIFO, L. M., 2005.- Identificación de elementos del paisaje prioritarios para la conservación de biodiversidad en paisajes rurales de los andes centrales de Colombia (en) SAENZ, J. & C. A. HARVEY, Conservación de Biodiversidad en Paisajes Fragmentados y Agropaisajes. Editorial UNA. Heredia. Costa rica. [ Links ]
MORÍ, S. & BOOM, B., 1987.- The Lecythidaceae of a lowland neotropical forest: La Fumeé Motain, French Guiana. Mem. New York. Bot. Garden, 44: 9-29. [ Links ]
REDFORD, K. H., TABER, A. & SIMONETTI, J. A., 1990.- There is more to biodiversity than the tropical rain forest. Conservation Biology, 4: 238-330. [ Links ]
RICHARDS, P. W., 1952.- The tropical rain forest. Cambridge University Press, Cambridge. [ Links ]
SHEIL, D. & BURSLEM, D. F. R. P., 2 003.- Disturbing hypotheses in tropical forests. Ecol. and Evol., 18(1): 18-26. [ Links ]
SNEATH, P. H. & SOKAL R. R., 1973.- Numerical taxonomic. Freeman, San Francisco. [ Links ]
SUMINA, O. I., 1994.- Plant communities on anthropogenically disturbed sites on Chukotka Peninsula, Russia. J. Veg. Sci., 5: 885- 896. [ Links ]
TABARELLI, M., MANTOVANI, W. & PERES C. A., 1999.- Effects of habitat fragmentation on plant guild structure in the montane Atlantic forest of southeastern Brasil. Biological Conservation, 91: 119-127. [ Links ]
TEBORGH, J., FOSTER, R. B. & NUÑEZ, V. P., 1996.- Tropical tree communities: A test of the nonequilibrium hypothesis. Ecology, 77: 561-567. [ Links ]
VALVERDE, B. O., 1998.- Estructura forestal y patrones florísticos de dos bosques tropicales húmedos de la cordillera de Talamanca, Costa Rica. Brenesia, 49 (50): 39-60. [ Links ]
VÁZQUEZ, G., J. A. & GIVNISH, T. J., 1998.- Altitudinal gradients in tropical forest composition, structure, and diversity in the Sierra of Manantlán. J. of Ecol., 86: 999-1020. [ Links ]
VÉLEZ, E. N., 1990.- Desarrollo y Conservación de Bosques Tropicales: 41-52 (en) Guadalupe (ed.): Seminario de Bosque Tropicales. Bogotá D. E., Colombia. [ Links ]
WHITMORE, T. C., 1975.- Tropical rain forests of the far east. Oxford University Press, Oxford. [ Links ]
WHITMORE, T. C., 1978.- Gaps in the forest canopy: 639-655 (en) by P. B. TOMLINSON & M. H. ZIMMERMANN (eds.) Tropical Trees as Living System. Cambridge University Press, Cambridge. [ Links ]
WHITMORE, T. C., 1998.- An introduction to tropical rain forest. Oxford University Press, New York. [ Links ]
WHITTAKER, J. A., 1956.- Vegetation of the great smoky mountains. Ecological Monographs, 26: 1-80. [ Links ]
ZAR, J. H., 1999.- Biostatistical Analysis. Fourth Edition. Prentice Hall. 663 pp. [ Links ]