











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
PermalinkCacao (Theobroma cacao L.) is a species native to the tropical rain forests of South America whose beans are the basic input for the chocolate industry and derivatives, cosmetic and pharmaceutical industries. Wild and cultivated cacao populations exhibit genetic variability due to sexual reproduction and natural cross-pollination. The pressure of evolutionary forces, particularly, the mutations have given rise to recessive genes that have survived for generations in heterozygous condition. Mutations occur at very low frequencies modifying the genotype causing unfeasibility of the embryo or premature death of some seedlings. In cacao seedlings from Parinari series was observed leaf chlorosis, followed of necrosis and finally death of seedling, at approximately 30 to 40 days (Almeida et al., 1998). It has been reported that Pa-30, Pa-121 and Pa-169 varieties when they were self or cross-fertilized and their progenies subjected to Chi-square (X2) test, segregated in a ratio 3:1 (normal: lethal), inferring that are heterozygous genotypes carrying the lethal gene ‘luteus-Pa’ (Bartley et al., 1983) and this gene is expressed only in recessive homozygous, being a character of monogenic inheritance (Bartley, 1969). Self-pollination or alopolinización between carriers’ heterozygous trees produce a phenotypic ratio 3:1 (normal and lethal) to the emergency and a ratio 2:1 (2 normal heterozygotes and 1 dominant homozygote) to adulthood (Yamada, 1982). Another study using six varieties from Parinari series pollinated with Pa-169 variety (carrying the lethal gene ‘luteus-Pa’), only Pa-44 variety showed segregation 3:1 verifying again monogenic inheritance (Yamada et al., 2005). A physiological study of cacao seedlings carriers of lethal gene ‘luteus-Pa’, identified differences between normal and mutant seedlings at 15 days after sowing. Variations in growth and subsequent death of seedlings to 60 days were related to the depletion of reserves from cotyledons, inactivity of the photosynthetic apparatus and oxidative stress (Rehem et al., 2011). Cacao as allogamous species, the lethal gene flow among populations can increase the frequency of this gene in the progeny and may become a limiting factor for mass propagation of seeds as rootstocks increasing production costs in the nursery and the inevitable replant new seeds. In some populations of common cacao exist heterozygous trees carriers of lethal gene ‘luteus-Pa’ who may have inherited from some ancient variety from Parinari series. The lethal gene ‘luteus-Pa’ may be useful for identifying seedlings belonging to the Pa series, in order to determine relationships between genotypes constituting a potential genetic marker and a tool for paternity tests (Bartley, 2005; Rehem et al., 2010). The aim of this study was to verify the type of inheritance of the ‘luteus-Pa’ gene in progenies resulting from self and cross fertilization of three varieties from Peruvian cacao.
MATERIALS AND METHODS
The study involved three places: EEA-Tulumayo, genebank cacao and deckhouse mesh of the Agronomy Faculty (Tingo María). The experimental soil consisted of a mixture of organic substrate of forest and sand (ratio 2:1). The internal temperature ranged from 22.2 to 25.6 °C at 08: 00h and from 28.4 to 34.5 °C at 14:00 pm with a brightness of 65% and 85% of relative humidity. The water requirements of the seedlings were satisfied with intraday irrigation. The genetic material consisted of three clones: S-5, C-25 and Pa-150, and this one had two components: selfing (S-5 Ø and C-25 Ø) and outcrossing (Pa-150 x C-25), utilizing eight pods from five trees for each clone. The seeds of the resulting progeny were planted in plastic containers and placed in the deckhouse mesh. The number of seeds per treatment was 200 (two replications of 100 seeds each one) sowed in four trays 25 seeds per container. For statistical analysis, the Chi-square test (X2) was chosen in order to test hypotheses about the kind of segregation from resulting progeny to one or a few pairs of genes of Mendelian inheritance. Its formula is as follows:
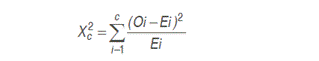
where:
Oi = i-th observed data
Ei = i-th data expected
C = number of phenotypic classes
The Xc 2 value will be contrasted with the Xt 2 (0.05, 1 df) value to test Ho.
The methodology used in the self-pollinations and cross-pollination involved (i) the choice of flower bud and isolation. To this purpose, 20 clear plastic tubes properly packed were placed above the flower buds and fixed to the tree until anthesis and hand pollination, (ii) for the self-pollinations, pollen from a drawn flower on the same tree was applied and then it was covered with the protective tube for eight days to avoid contamination with foreign pollen, and among eight to ten days the fruit was covered with a transparent plastic bag cutted in the bottom ends to evacuate transpired water and protect of the infection from fungi spores and (iii) cross-pollination, it was similar to that described for the self-pollinations, with the difference in the pollen donor parent. Pollen receptors flowers corresponded to Pa-150 variety and as pollen donors to C-25 variety. Monitoring the growth of fruits fertilized from self and cross pollination was performed every two weeks during six months. At harvest the fruits were labeled, seeds were washed with sawdust to remove the mucilage and then disinfected with Cupravit (5 g kg-1 seed) just before sowing. The date of sowing and seedling emergence, the identifying of emergent seedlings: normal and chlorotic (lethal) and intraday monitoring of chlorosis evolution until the senescence and death of mutant plants (lethal), were recorded.
RESULTS AND DISCUSSION
Chi-square test (X 2 ) for progenies from selfing
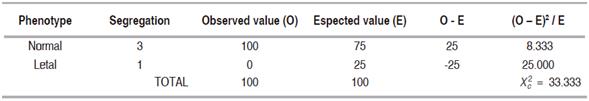
The Table 1, shows that the S-5 progeny resulting from self-pollination, no exhibited none lethal seedling, being all normal. The 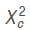 = 33.333 value was higher to Xt
2
(0.05, 1df) = 3.841 value, and as falls out of the region of Ho acceptation, so we inferring this progeny not be adjust to ratio 3:1 (normal: lethal) of the expected Mendelian segregation and consequently, this genotype is not carrier of lethal gene ‘luteus-Pa’ in heterozygote condition.
= 33.333 value was higher to Xt
2
(0.05, 1df) = 3.841 value, and as falls out of the region of Ho acceptation, so we inferring this progeny not be adjust to ratio 3:1 (normal: lethal) of the expected Mendelian segregation and consequently, this genotype is not carrier of lethal gene ‘luteus-Pa’ in heterozygote condition.
Instead in Table 2, the progeny of the C-25 variety resulting from self-pollination, the segregation of seedlings: normal and lethal in a ratio 3:1 was observed. On average 100 emergent seedlings (replicate 1), 71 seedlings exhibited normal green colour (synthetizing chlorophyll) and 29 seedlings not exhibited (non-synthetizing chlorophyll). The 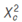 = 0.853 value was inferior to Xt
2
(0.05, 1df) = 3.841 value and as falls out of the region of acceptation of Ho, it can be concluded that observed segregation ratio in the resulting progeny from selfing C-25 variety, it fits the pattern of expected Mendelian segregation 3:1. Likewise, similar = 0.853 values were obtained in the replicate 2. So, it can be inferred that C-25 variety is carrier of the lethal gene ‘luteus-Pa’ in heterozygote condition (LPlp) and the lethal seedlings are of genotype recessive homozygote (lp lp).
= 0.853 value was inferior to Xt
2
(0.05, 1df) = 3.841 value and as falls out of the region of acceptation of Ho, it can be concluded that observed segregation ratio in the resulting progeny from selfing C-25 variety, it fits the pattern of expected Mendelian segregation 3:1. Likewise, similar = 0.853 values were obtained in the replicate 2. So, it can be inferred that C-25 variety is carrier of the lethal gene ‘luteus-Pa’ in heterozygote condition (LPlp) and the lethal seedlings are of genotype recessive homozygote (lp lp).
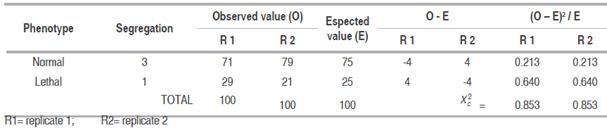
Similar results of segregation of normal and lethal seedlings (“chlorophyll deficient mutants”) in a phenotypic 3:1 ratio from selfing have been reported for cacao varieties from Parinari series (Yamada,1982; Bartley et al., 1983), corroborating the monogenic inheritance of lethal gene ‘luteus-Pa’ and identifying to the C-25 variety as a carrier genotype of this lethal gene. We believe that C-25 may be inherited the ‘luteus-Pa’ gene from cultivar Pa-169 already identified as carrier and this one might have participated in different crosses in cacao breeding programs in Brazil and whose hybrid seed mixed with others hybrids it would arrived in Peru in the late 80s or earlier decades from Brazil, being found in some farms of Huánuco, San Martin and Rio Apurimac-Ene valley. Lethal seedlings (“mutants deficient in chlorophyll”) die shortly after emergence because chlorophyll is essential for photosynthesis processes and also in the absorbance and transference light energy to the reaction centers within of the chloroplasts (Ning et al., 2013). His acute deficiency causes stunted growth, senescence and subsequent death. Although the genetic mechanism is not known, it has been reported that the yellow-pale (lethal) phenotype could be due to a problem that arises in the membranes of thylakoid which is essential for the photosynthetic process (Reed et al., 2014).
Chi-square test (X 2 ) for the progenies of cross-fertilization
The Table 3 show the results of the segregation of cacao seedlings from Pa-150 x C-25 cross. We can observe there the absence of segregation of normal and lethal seedlings in a phenotypic ratio 3:1 and being all (100%) of normal green colour and 0% abnormal (yellowish). The = 33.333 value was higher to Xt
2
(0.05, 1df) = 3.841 and as falls out of the Ho acceptance region, we can conclude which this obtained progeny by cross-fertilization does not adjust to expected Mendelian segregation 3:1 (normal: lethal), rejecting the Ho ( 3:1ratio). These results allow us to infer that Pa-150 variety (Parinari series) does not carry the lethal gene ‘luteus-Pa’ but the C-25 variety does; otherwise, they would have appeared as lethal seedlings in a 25% (expected frequency). Reports from Brazil admit that not all varieties from Parinari series are carriers of this lethal gene ‘luteus-Pa’, with exception of the Pa-169 variety (Yamada, 1982; Bartley et al., 1983). Generally recessive lethal alleles have variable frequencies in the populations and they can persist over a long period of time. Natural selection eliminates copies of allele when they occur in homozygote genotypes; however, recessive lethal alleles are suppressed by dominant alleles in heterozygote genotypes, and they evade natural selection; hence heterozygote carriers of recessive lethal gene have a selective advantage in the natural selection process (Ryder et al., 1999).
Physiogenetics of evolution of the chlorosis from lethal seedlings
Chlorophyll deficiency or albinism is perhaps the most easily mutant phenotype detectable in higher plants and in Arabidopsis thaliana. Among the various mutants of normal green leaves, e.g. Chlorina mutant (green-yellowing), the physiology and genetic mechanism is different from others mutants (Jarvis et al., 2000). The effects of mutations on chloroplast development, photosynthesis, accumulation of chlorophyll b, and levels of uptake of light by chlorophyll a/b associated proteins, have been examined by (Kosuge et al. 2011). Conditional lethal plants have been obtained by inactivation of the ribulose-1,5-bifosfatocarboxilasa-oxygenase enzyme catalyzing both oxidation and carboxylation of ribulose-1,5-bisphosphate resulting in uptake of O2, CO2 release (photorespiration) and NH4 (Migge and Becker, 2000). In the Figure 1, the evolution of lethal chlorosis in seedlings from selfing C-25 variety carrying the lethal gene ‘luteus-Pa’ is showed. There we can see the chlorotic leaf yellowing followed by necrosis, senescence and death of the seedlings in a period which range among12 to 28 days after emergence. According to (Rehem et al. 2011), the leaf yellowing is a symptom of senescence associated with chlorophyll degradation and catabolized by polyphenol oxidase, among others.When occur the accumulation of anthocianins in the senescents leaves, this is preceded by chlorophill degradation (Díaz et al., 2006).The persistence of the chlorophyll when the photosynthetic capacity is affected, increases susceptibility to oxidative stress induced by light in the leaf cells. A protective role of anthocyanins as sunscreens and as neutralizers for reactive oxygen species (ROS) in young expanded leaves and senescent leaves susceptible to damage by light, it has been suggested by Baker and Hardwick (1972). On the other hand, the ‘shared pathway’ approach to lethals gene identification, the complexity and redundancy of metabolic pathways in plants may limit the strategy overall. The assumption here is that disrupting any portion of the complex proteic will result in lethality (Meinke et al., 2008). Studies with molecular markers found that the level of polymorphism between Pa-30 and Pa-169, based on RAPD and SSR markers, was compatible with the average values usually found in genetic diversity studies of T. cacao (Rehem et al., 2010). Data from ESTs identified genes associated with Rubisco, peroxidases, and other proteins and enzymes related to carbon assimilation, respiration, and photosystem 2. Mutant seedlings were characterized by synthesizing defective PsbO and PsbA proteins, which were overexpressed from 15 to 20 days after seedling emergence (Rehem et al., 2016). A better understanding on the photobiological origin, the mapping with molecular markers and functional genomics of chlorophyll deficient mutants, can help to identify and understand the factors involved in the photosynthetic mechanism.

CONCLUSIONS
In the progenies from selfing, only C-25 variety showed a Mendelian segregation 3:1, identifying it as a heterozygote carrier of lethal gene ‘luteus-Pa’.
The progeny from the crossing Pa-150 x C-25 does not gave none lethal seedling failing in adjust to Mendelian segregation 3:1, therefore we can to infer that the Pa-150 variety, is not carrier of lethal gene ‘luteus-Pa’.
The physiogenetics basis of the chlorophyll deficient mutants are not yet very clear to understand the factors involved in the genetic control, the photosynthetic mechanism and its photobiological origin.

