











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
La cabecera de las cuencas de las quebradas Yomasa y Fucha abarca amplias zonas de pajonales y matorrales y bosques en el páramo de Cruz Verde, sobre la antigua vía Bogotá -Ubaque, la antigua vía al Llano y el Agroparque Los Soches; en su parte baja estas cuencas presentan una dinámica de urbanización legal e ilegal muy acelerada y desordenada, además de procesos sociales complejos asociados a dichas condiciones (Torres, 2016). Sin embargo, la proporción de las áreas rurales y silvestres sigue siendo mayor que la urbana, por lo que la zona tiene un gran potencial agrícola y turístico para sus comunidades y para Bogotá. Este es el caso del Agroparque Los Soches, el cual desarrolla diferentes actividades de educación ambiental orientadas al agroturismo y la conservación de la diversidad biológica y cultural (García, 2020; Melgarejo, 2009). No obstante, la presión urbana es constante y los proyectos urbanísticos del sector público ya cuentan con áreas destinadas a su desarrollo denominadas "suelo de expansión" (Melgarejo, 2009). Ambas quebradas son fuente importante de agua para los habitantes de la zona, pero una vez entran en contacto con los asentamientos humanos en áreas rurales, periurbanas y urbanas, se convierten en recolectores de aguas residuales (Chavarro & Bernal, 2016; Moreno-Murillo, 2001). Esta degradación, y la introducción de especies exóticas, son una barrera para la conservación de la biodiversidad y para el disfrute de los beneficios o servicios que brinda (fertilidad del suelo, agua y aire puros) por parte de la ciudadanía.
La flora y la vegetación actuales en la sabana de Bogotá y las montañas del Distrito Capital son producto del proceso de transformación debido a las actividades antrópicas (Cleef & Hooghiemstra, 1984; Cortés-S., et al., 1999; Cortés-Sánchez, 2011). La fragmentación y la degradación provocadas por estas actividades influyen en el cambio de las dinámicas naturales de los ecosistemas, reduciendo potencialmente los servicios ambientales que brindan (Aronson, et al., 2007) y generando un panorama desalentador en cuanto a la conservación de los recursos naturales (Moreno-Mateos, et al., 2017). Sin embargo, incluso en los ecosistemas resultantes de toda esta alteración (agroecosistemas, plantaciones y ciudades) es posible mejorar o recuperar los servicios perdidos (Augstburger, et al, 2018; Bolund & Hunhammar, 1999; Chazdon, 2008).
En este contexto, el concepto de sucesión ecológica resulta fundamental para la planificación de acciones de conservación, restauración y manejo de todos estos ecosistemas. El concepto inicial de sucesión se planteó como una secuencia lineal de reemplazo de especies que finaliza en un único clímax o estado ideal, pero actualmente se sabe que pueden existir múltiples clímax o estados estables finales que responden a diferentes condiciones ambientales dentro de una misma área (Gotelli, 2008). En conexión con este han aparecido otras propuestas, como la de estados alternativos estables (Beisner, et al., 2003) o la de vegetación natural potencial múltiple (Somodi, et al., 2017), las cuales son más apropiadas para afrontar los procesos dinámicos de los ecosistemas (Suding & Hobbs, 2009) y abordar paisajes altamente transformados donde hay un amplio rango de comunidades en vez de una única comunidad previa al disturbio (Török, et al., 2018) cuya composición histórica provee una base sólida para planificar acciones de restauración.
Los principales referentes sobre la composición y estructura de la vegetación del sector son los estudios hechos en el páramo de Cruz Verde (Lozano-Contreras & Schnetter, 1976), el páramo de Monserrate (Vargas & Zuluaga, 1985) y el páramo de Chisacá (Pedraza-Peñalosa, et al., 2004), los relacionados con los patrones de vegetación remanente de la Serranía de Chía (Cortés-S., et al., 1999) y la cuenca alta del río Bogotá (Cortés-S., 2008), así como del municipio de Soacha (Ariza, et al., 2010), con la vegetación de los cerros Orientales de Bogotá en zonas naturales (Cantillo-Higuera & Gracia, 2013) y en zonas bajo procesos de restauración ecológica (Rojas-B., 2017), y con la delimitación del complejo de páramos Sumapaz-Cruz Verde (UDFJC & IAvH, 2015).
En este contexto, es importante generar una línea de base sobre la flora y la vegetación presentes en el sector para planificar la conservación de los ecosistemas en buen estado o la restauración ecológica de los que estén degradados. Por ello el objetivo del presente estudio fue analizar la riqueza, composición y estructura de la vegetación en las cuencas de las quebradas Yomasa y Fucha con base en las siguientes preguntas de investigación: 1) ¿cuáles son las comunidades presentes y cuál su estado de desarrollo estructural? 2) ¿Cuál es el patrón de riqueza florística en dichas comunidades? 3) ¿Cómo es la composición y la estructura de las comunidades encontradas? Las respectivas hipótesis fueron: 1) hay comunidades florísticas con varios estados de desarrollo o complejidad estructural; 2) la riqueza es mayor en los bosques y arbustales donde hay menor presión antrópica, y 3) la composición es similar a la registrada en las montañas circundantes de la sabana de Bogotá, pero las comunidades identificadas pueden encontrarse en estados de baja complejidad estructural, pues en esta cuenca predomina la actividad agrícola en antiguas zonas de bosques y páramos.
Materiales y métodos
Area de estudio
Las cuencas de las quebradas Yomasa y Fucha se ubican en la localidad de Usme, principalmente en zona rural, y cubren veredas como Los Soches y El Uval en las coordenadas 4°27'53.0"N 74°06'23.2"W - 4°30'53.6"N 74°02'59.9"W (Figura 1). La temperatura fluctúa entre los 9 °C y los 17 °C y la variación altitudinal entre los 2.800 y los 3.300 m s.n.m. (Chavez & Quiroga, 2017). El clima es húmedo, con un promedio anual de humedad relativa que supera el 70 %, y la precipitación media anual es de 82,58 mm, con un régimen bimodal-tetraestacional cuyos máximos se registran en febrero (105,4 mm) y octubre (105,1 mm) (Cantillo-Higuera & Gracia, 2013). El uso del suelo en la vereda Los Soches es agrícola, ganadero, forestal y de explotación mineral (canteras) (Chavez & Quiroga, 2017; Franco, 2016; Mejía, 2013), aunque aún alberga vegetación nativa de bosque alto andino, subpáramo y páramo asociada al páramo de Cruz Verde, el cual hace parte del complejo de páramos Cruz Verde-Sumapaz (Colorado, 2018). La mayoría del área en la vereda El Uval se dedica a actividades agropecuarias y, en menor medida, a otras producto de la expansión urbana (Beltrán, et al., 2016).
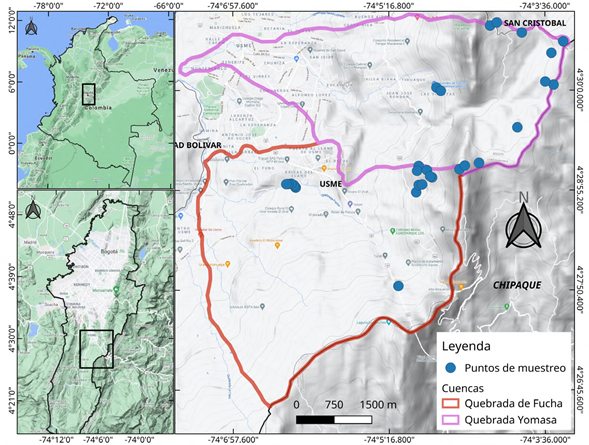
Figura 1 Localización y límites de las cuencas de las quebradas Yomasa y Fucha y ubicación de los puntos de muestreo (Google Maps, 2021). Mapa de terreno de la ciudad de Bogotá, Colombia. https://www.google.com/maps/place/Bogot%C3%A1/@4.6482837,-74.2478968,11z/data=!3m1!4b1!4m5!3m4!1s0x8e3f9bfd2da6cb29:0x239d635520a33914!8m2!3d4.7109886!4d-74.072092
Fase de campo
Los datos en campo se recolectaron a partir del 2015 en el marco del proyecto "Nodos de biodiversidad", y hasta el 2019 en el de "Caracterización ecológica de la Reserva Forestal Protectora Bosque Oriental de Bogotá", ambos desarrollados por el Jardín Botánico de Bogotá. Siguiendo las recomendaciones del proyecto de delimitación de páramos para áreas de muestreo en la alta montaña colombiana (UDFJC & IAvH, 2015), se utilizaron 17 parcelas de 25x2 m (50 m2) para la vegetación herbácea o arbustiva (menos de 5 m de altura) y dos parcelas de 50x2 m (100 m2) para la vegetación arbórea (más de 5 m de altura) (Figura 1).
Se midió el diámetro del tronco y la altura total de los individuos leñosos. Según la metodología para delimitación de páramos (UDFJC & IAvH, 2015), se midió la circunferencia basal del tallo a 30 cm del suelo en pastizales y arbustales (altura<5 m), y a 1,3 m del suelo (diámetro a la altura del pecho, DAP) en vegetación arbórea como plantaciones y bosques (altura>5 m). Se utilizó el método punto-intercepto para estimar el porcentaje de cobertura de las especies y los estratos de cada parcela (Brower, et al., 1998; Elzinga, et al., 1998). El método se implementó registrando el contacto vertical de las especies en cada estrato o franja de vegetación. Para la vegetación herbácea o arbustiva esto se hizo cada 50 cm a lo largo de una línea de 25 m dentro del área de 25x2 m de la parcela para un total de 50 puntos, lo que da una precisión mínima de 2 % de cobertura. Para la vegetación arbórea el procedimiento se aplicó cada 100 cm a lo largo de una línea de 50 m dentro del área de 50x2 m de la parcela para un total de 50 puntos, lo que también da una precisión mínima de 2 % de cobertura. Los estratos según su altura fueron: a) rasante (r), de 0 a 10 cm de altura; b) herbáceo (h), de 10 a 50 cm; c) arbustivo bajo (b1), de 50 a 1,5 m; d) arbustivo alto (b2), de 1,5 a 3 m; e) subarbóreo inferior (a1), de 3 a 5 m; subarbóreo superior (a2), de 5 a 12 m; f) arbóreo inferior (s1), entre 12 y 25 m, y arbóreo superior (s2), más de 25 m de altura. Se registraron todas las especies de plantas vasculares en los levantamientos, incluso aquellas no capturadas por el método de medición de la cobertura. Se recolectaron muestras botánicas de todas las especies en estado fértil dentro y fuera de las parcelas (colecciones generales) y se depositaron en el Herbario del Jardín Botánico de Bogotá (JBB).
Análisis de la información
Se hizo un análisis de conglomerados para evaluar la composición o similitud florística entre las parcelas; en el dendrograma resultante se identificaron grupos florísticos que en adelante se denominan "comunidades". Se utilizó la suma del porcentaje de cobertura por especie como variable de comparación, el índice de Horn como medida de similitud (Brower, et al., 1998), y el método de agrupamiento de unión de vecinos (neighbor joining) (Podani, et al., 2000; Saitou & Nei, 1987); para enraizar el dendrograma se utilizó como grupo externo (outgroup) una parcela hipotética con valor de cero para todas las especies. Este análisis se hizo con ayuda del programa estadístico PAST 3.04 (Hammer, et al., 2001). Se hizo otro análisis de conglomerados para evaluar la similitud estructural entre las parcelas y en el dendrograma resultante se identificaron grupos estructurales denominados en adelante "estados de desarrollo o complejidad estructural". También se utilizó el método de neighbor joining, pero usando como variables de comparación la suma del porcentaje de cobertura por estrato y por forma de crecimiento y el porcentaje de suelo desnudo y de hojarasca; la distancia Manhattan se empleó como medida de similitud. Para enraizar el dendrograma se utilizó como grupo externo una parcela hipotética con valor de cero para todas las variables, excepto para el suelo desnudo, en donde el valor fue de 100 %. Las dos matrices de composición y estructura por parcela se unieron y se analizaron mediante un análisis dimensional no métrico (non-metric multidimensional scaling, NMDS), con el fin de evidenciar los estados de desarrollo estructural de las comunidades y los parámetros que más se relacionaban con cada uno (Anexo S1,https://www.raccefyn.co/index.php/raccefyn/article/view/1255/3080). Este análisis también se hizo con el programa PAST 3.04.
La riqueza esperada y el esfuerzo de muestreo se estimaron para las comunidades florísticas encontradas con ayuda del programa iNEXT (Chao, et al., 2016): se emplearon datos de presencia-ausencia usando los interceptos como unidad muestral y el enfoque no asintótico por interpolación y extrapolación para elaborar las curvas de acumulación correspondientes.
El resultado de los análisis de conglomerados y de ordenación se utilizó para elaborar una clasificación combinada de composición y estructura, de manera que para cada comunidad se presenta una descripción de sus estados de desarrollo o complejidad estructural, así como las especies más abundantes, los taxones con mayor riqueza y el porcentaje de cobertura total del suelo por estrato y por forma de crecimiento.
Resultados
Comunidades florísticas y desarrollo estructural
El análisis de similitud florística arrojó seis comunidades ordenadas con respecto al grupo externo según su composición de especies, y el primer grupo (con una sola comunidad: C1) se denominó "sabana de Bogotá" (por encontrarse en la parte baja de la cuenca y concentrar la mayor extensión de cultivos y zonas de pastoreo, es decir, la matriz agropecuaria), en tanto que el segundo grupo se denominó "alta montaña" (por ocupar la parte alta de la cuenca). Con excepción de una plantación forestal (C2) que alberga especies silvestres de alta montaña en el sotobosque, en este grupo se distinguieron una comunidad de bosque (C3) y varias comunidades de páramo (C4-C6) que comparten las especies Gaultheria anastomosans, Bucquetia glutinosa, Hesperomeles obtusifolia y Calamagrostis effusa (Figura 2A). El análisis de similitud estructural estableció siete estados de desarrollo o complejidad estructural, que en orden de mayor a menor complejidad se denominaron: E1) bosque alto, E2) bosque bajo, E3) arbustal denso, E4) pastizal arbolado, E5) matorral denso, E6) matorral abierto, y E7) pastizal. Los estados E1 y E2 constituyen la vegetación boscosa, y de E3 a E7 conforman la vegetación abierta (Figura 2B).

Figura 2 Clasificación de la vegetación en la cuenca de la quebrada Fucha. A) Dendrograma de similitud florística. B) Dendrograma de similitud estructural. C) Ordenación florística y estructural de la vegetación en la cuenca de la quebrada Fucha. En color se diferencian las comunidades florísticas: C1: bosque de la sabana de Bogotá (naranja), C2: plantación forestal (gris), C3: bosque altoandino (verde), C4: matorral mixto de páramo (morado), C5: arbustal de páramo (azul), C6: pajonal de páramo (cian). La variación de tonalidad del mismo color indica diferencias estructurales al interior de la comunidad. E1: bosque alto, E2: bosque bajo, E3: arbustal denso, E4: pastizal arbolado, E5: matorral denso, E6: matorral abierto, E7: pastizal
Teniendo en cuenta la afinidad florística de cada parcela (Figura 2A), mediante la representación por colores de las diferentes comunidades en el análisis de afinidad estructural (Figura 2B) y en el de ordenación (Figura 2C) se pudo apreciar que algunas comunidades presentaban más de un estado de desarrollo estructural (representado en la variación del color principal): por ejemplo, las comunidades C1 y C3 mostraban tres estados (tres tonalidades de naranja y verde, respectivamente), y la comunidad C5, dos estados (dos tonalidades de azul). Según el análisis NMDS, el primer eje agrupó el 70 % de la variación y en términos estructurales separó claramente hacia el extremo negativo las parcelas con vegetación boscosa, mientras que las parcelas con vegetación abierta se ubicaron hacia el centro y el extremo positivo del eje (Figura 2C). El extremo negativo del primer eje evidenció un mayor peso de las variables de cobertura de árboles, arbustos y bambusoides, de hojarasca, y de cobertura en los estratos subarbóreo (alto y bajo) y arbustivo alto; el extremo positivo tuvo un mayor peso de las variables de cobertura de los estratos rasante y herbáceo y de herbáceas, musgos, hepáticas y epífitas vasculares (Figura 2C). Por el otro lado, el segundo eje agrupó el 14 % de la variación, dando resolución a la separación entre los estados E1 (bosque alto) y E2 (bosque bajo), y entre E3 y E4 (arbustal denso y pastizal arbolado) y E5-6 (matorral denso y abierto) y E7 (pastizal). Este eje presentó en su extremo positivo un mayor peso de las variables de cobertura del estrato subarbóreo bajo, de epífitas, trepadoras y porcentaje de suelo desnudo; en el extremo negativo presentó mayores valores de las variables de cobertura del estrato arbustivo bajo y de arbustos, líquenes, hepáticas y musgos (Figura 2C).
Riqueza florística
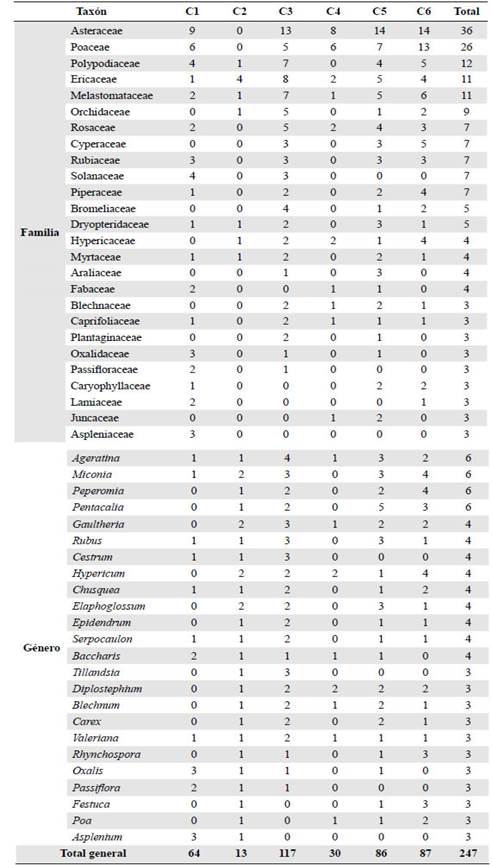
En el muestreo total, incluidas las colecciones generales, se registraron 247 especies, 151 géneros y 64 familias (Tabla 1yAnexo S2, https://www.raccefyn.co/index.php/raccefyn/article/view/1255/3081); los taxones con mayor riqueza fueron: Asteraceae, Poaceae, Polypodiaceae, Melastomataceae, Ericaceae, Orchidaceae, Cyperaceae, Piperaceae, Rosaceae, Rubiaceae, Solanaceae, Miconia, Ageratina, Pentacalia, Peperomia, Hypericum, Gaultheria, Chusquea, Rubus, Elaphoglossum y Epidendrum (Tabla 2). Las comunidades con mayor riqueza estimada (q0) fueron la C3 (bosque altoandino), C5 (arbustal de páramo) y C6 (pajonal de páramo), en tanto que la riqueza más baja se observó en C2 (plantación de pino) y C4 (matorral mixto de páramo); según el estimador de la cobertura muestral (C.hat), la representatividad del muestreo superó el 90 % en todas las comunidades identificadas (Figura 3ytabla 1).
Tabla 1 Riqueza florística y esfuerzo de muestreo por comunidad vegetal en las cuencas de las quebradas Yomasa y Fucha

T: número de unidades de muestreo observadas en la muestra de referencia (interceptos); U: frecuencia de aparición en la muestra de referencia; S.obs: número de especies observadas; qD: diversidad estimada de orden q0 (riqueza) para una muestra de 500 interceptos; qD.LCL, qD.UCL: límites de confianza inferior y superior de bootstrap para la diversidad de orden q0 en el nivel 0,95; C.hat: estimador de la cobertura muestral sugerido por Chao & Jost (2012); C1: bosque de la sabana de Bogotá; C2: plantación forestal; C3: bosque altoandino; C4: matorral mixto de páramo; C5: arbustal de páramo; C6: pajonal de páramo
Tabla 2 Familias y géneros con mayor riqueza de especies por comunidad florística en las cuencas de las quebradas Yomasa y Fucha
C1: bosque de la sabana de Bogotá, C2: plantación forestal, C3: bosque altoandino, C4: matorral mixto de páramo, C5: arbustal de páramo, C6: pajonal de páramo

Figura 3 Curvas de acumulación de especies de las comunidades florísticas en las cuencas de las quebradas Yomasa y Fucha. Diversidad estimada de orden q0 (riqueza). C1: bosque de la sabana de Bogotá; C2: plantación forestal; C3: bosque altoandino; C4: matorral mixto de páramo; C5: arbustal de páramo; C6: pajonal de páramo
Composición y estructura de las comunidades
A partir de los análisis de similitud y de ordenación, se obtuvo la clasificación combinada que muestra la variación en la composición y la estructura de cada comunidad en cada uno de sus estados de desarrollo (Figura 4yfigura 5): Sabana de Bogotá

Figura 4 Clasificación estructural de la vegetación para cada comunidad florística en las cuencas quebrada Yomasa y quebrada Fucha.

Figura 5 Comunidades florísticas en las cuencas de las quebradas Yomasa y Fucha. A) Bosque de la sabana de Bogotá (C1). B) Plantación forestal (C2). C) Bosque altoandino (C3). D) Matorral mixto de páramo (C4). E) Arbustal de páramo (C5). F) Pajonal de páramo (C6).
C1) Fisionomía dominante: bosque. Especies más abundantes: Varronia cylin-drostachya, Vallea stipularis, Chusquea scandens y Cenchrus clandestinus. Taxones con mayor riqueza: Asteraceae, Poaceae, Polypodiaceae, Solanaceae, Asplenium y Oxalis. Presenta tres estados de desarrollo estructural, los valores más altos de cobertura se concentran en: E1) subarbóreo bajo y arbustivo alto; E3) arbustivo alto y bajo, y herbáceo; E4) rasante y herbáceo. Formas de crecimiento más abundantes: hierba postrada (E4), árbol (E4 y E3), trepadora leñosa (E1), arbusto (E1) y bambusoide (E1).
Alta montaña
C2) Fisionomía dominante: bosque. Especies más abundantes: Pinus patula, Miconia ligustrina y Macleania rupestris. Taxones con mayor riqueza: Ericaceae, Miconia, Elaphoglossum, Gaultheria e Hypericum. Estado de desarrollo E1 (bosque alto). Mayor porcentaje de cobertura: estratos subarbóreo bajo y alto, y en el arbustivo alto. Formas de crecimiento más abundantes: {arbol, arbusto y musgo.
C3) Fisionomía dominante: bosque. Especies más abundantes: Weinmannia tomentosa, Vallea stipularis, Chusquea spencei y Chusquea scandens. Taxones con mayor riqueza: Asteraceae, Ericaceae, Polypodiaceae, Melastomataceae, Ageratina, Miconia, Cestrum, Gaultheria, Rubus y Tillandsia. Presenta tres estados de desarrollo estructural, los valores más altos de cobertura se encuentran en: E1) subarbóreo alto y bajo; E2) subarbóreo bajo y arbustivo; E3) herbáceo y rasante. Formas de crecimiento más abundantes: arbusto (E6), áÁrbol (E1), bambusoide (E2), musgo (E2) y liquen (E6).
C4) Fisionomía dominante: matorral. Especies más abundantes: Cortaderia columbiana, Pernettya prostrata, Ulex europaeus e Hypericum myricariifolium. Taxones con mayor riqueza: Asteraceae, Poaceae, Ericaceae, Diplostephium, Hypericum y Agrostis. Estado de desarrollo E6 (matorral abierto). Mayor porcentaje de cobertura: estratos herbáceo y rasante. Formas de crecimiento más abundantes: arbusto, macolla, musgo, hierba erecta y liquen.
C5) Fisionomía dominante: matorral. Especies más abundantes: Gaultheria anasto-mosans, Tibouchina grossa, Hypericum goyanesii y Blechnum schomburgkii. Taxones con mayor riqueza: Asteraceae, Poaceae, Ericaceae, Melastomataceae, Pentacalia, Ageratina, Miconia, Rubus y Elaphoglossum. Presenta dos estados de desarrollo estructural E5 y E6, en los valores más altos de cobertura ambos estados se encuentran en los estratos herbáceo y rasante, pero en E5 hay más cobertura en arbustivo bajo. Formas de crecimiento más abundantes: arbusto (E5-E6), musgo (E5), Caulirrósula (E5), hierba postrada (E5) y macolla (E6).
C6) Fisionomía dominante: pajonal. Especies más abundantes: Calamagrostis effusa, Festuca pilar-franceii, Espeletia grandiflora y Gaultheria anastomosans. Taxones con mayor riqueza: Asteraceae, Poaceae, Melastomataceae, Polypodiaceae, Cyperaceae, Miconia, Peperomia, Hypericum, Pentacalia, Rhynchospora y Festuca. Estado de desarrollo E7 (Pastizal). Mayor porcentaje de cobertura: estratos rasante y herbáceo. Formas de crecimiento más abundantes: macolla, arbusto, musgo, caulirrósula, y hierba postrada.
Discusión
Comunidades florísticas y desarrollo estructural
Los análisis confirmaron la hipótesis sobre la existencia de comunidades florísticas con varios estados de desarrollo estructural. Estas comunidades no son más que grupos ad hoc, delimitados artificialmente dentro de un conjunto reducido de datos, pero su existencia como entidades discretas o como estados estables se propone como hipótesis para ser corroborada posteriormente con un mayor volumen de datos; los estados de desarrollo presentados también son hipotéticos, pero están inspirados en propuestas como la de la base de datos Corine Land Cover, la cual establece umbrales de atributos estructurales entre un tipo discreto de cobertura vegetal y otra para una unidad mínima susceptible de cartografiarse (IDEAM, 2010). Con los análisis realizados se pretende mostrar la posibilidad de que estas comunidades o ensamblajes florísticos presenten estados de desarrollo estructural dentro de unos límites estables, fuera de los cuales la comunidad (o el estado) se transforma en una totalmente diferente (Beisner, et al., 2003). En este caso, si cada comunidad representa un estado alternativo estable, los límites o umbrales estarían definidos por una composición y una abundancia de especies específicas independientes de los atributos estructurales: por ejemplo, tres comunidades distintas, como C1, C2 y C3, pueden presentar la misma estructura de bosque alto, u otras tres, como C3, C4 y C5, pueden presentar la misma estructura de matorral abierto, en tanto que C3 tiene una identidad florística independiente del tamaño de los árboles y arbustos que la conforman. Esta aproximación puede ser similar al concepto de comunidades seriales (Huschle & Hironaka, 1980), que en sinfitosociología representan la evolución hacia el óptimo estable, clímax o comunidad permanente (Martínez, 1979), un concepto derivado del paradigma de la facilitación y la sucesión lineal de Clements (Gotelli, 2008). Sin embargo, dado que no se trabajó bajo un esquema sinfitosociológico, los estados de desarrollo estructural observados en esta cuenca no se asumieron como comunidades de una sinasociación (Martínez, 1979), sino como parte de la variación de una misma comunidad. La comprobación de que efectivamente estas comunidades sean estados estables requerirá un mayor muestreo en un área más amplia, pero los datos que se presentan aquí pueden ser el insumo para abordar este enfoque que no se ha trabajado detalladamente en Colombia, y que puede representar una mejor aproximación para la planificación de acciones de manejo, conservación y restauración (Beisner, et al., 2003; Suding & Hobbs, 2009).
Riqueza florística
Tal como se planteó inicialmente, la riqueza fue mayor en el bosque altoandino (C3) y los arbustales de páramo (C5) e, incluso, el pajonal de páramo mismo (C6) tuvo más especies que otras comunidades boscosas a menor altitud en esta cuenca; probablemente la riqueza total registrada en el pajonal de páramo sea ligeramente superior a la de los arbustales, pues en estos hubo menos recolecciones generales. Este patrón es el mismo que se observa en otras zonas de alta montaña en la sabana de Bogotá, como la cuenca alta del río Subachoque (Fernández-Alonso & Hernández-Schmidt, 2007), la flora de Soacha (Ariza, et al., 2010) o la de los cerros Orientales (Cantillo-Higuera & Gracia, 2013), o la caracterización del complejo de páramos Sumapaz-Cruz Verde (UDFJC & IAvH, 2015). La riqueza decreciente con el incremento en la elevación y la disminución de la precipitación también coincide con el patrón de montaña del trópico (Gentry, 1988), ya que las comunidades de arbustal y pajonal de páramo ocupan las partes más altas de la cuenca.
Asimismo, el patrón de riqueza de las familias y géneros con más especies coincidió con la registrada en los anteriores estudios (Ariza, etal., 2010; Cantillo-Higuera & Gracia, 2013; Fernández-Alonso & Hernández-Schmidt, 2007; Pedraza-Peñalosa, etal., 2004; UDFJC & IAvH, 2015). Lo interesante es que estas comunidades aún conservan un patrón de riqueza de especies representativo de la región, a pesar de la vocación agropecuaria y forestal de este sector del Distrito Capital (Chávez & Quiroga, 2017), probablemente porque la parte baja de la cuenca (más relacionada con la vegetación de la parte plana de la sabana de Bogotá) ha sido la más transformada por las actividades humanas desde la época precolombina hasta la actualidad (Cleef & Hooghiemstra, 1984; Cortés-S., 2008).
Composición y estructura de las comunidades
La composición fue similar a la registrada en las montañas circundantes de la sabana de Bogotá, como ya se señaló, pero algunas comunidades identificadas mostraron estados de alta complejidad estructural, incluso en la parte baja con mayor presión agrícola. En términos de atributos florísticos y estructurales se destacan los siguientes aspectos: 1) las comunidades de pajonal (C6) y arbustal (C5) de páramo coinciden con los resultados de comunidades similares en los páramos de Soacha, Monserrate, Cruz Verde y Sumapaz (Ariza, et al., 2010; Lozano-Contreras & Schnetter, 1976; Sturm & Abouchaar, 1981; UDFJC & IAvH, 2015); no obstante, la comunidad C4 no parece guardar relación con la vegetación descrita en estos trabajos, a pesar de que alberga especies comunes, lo que tal vez se deba al hecho de que se desarrolla sobre zonas alteradas, como cunetas en el borde de la carretera, y a la presencia de la especie exótica Ulex europeaus. Aparentemente esta especie no tiene un alto potencial invasor a gran altitud (Ocampo-Zuleta & Solorza-Bejarano, 2017) y solo se distribuye hasta los 3.180 m s.n.m. (Ruiz, et al., 2020), pero este registro confirma que ya se ha naturalizado en zonas de páramo por encima de los 3.400 msnm. 2) En el caso de los bosques, también hay coincidencia en cuanto a la composición y estructura de la comunidad C3 con el bosque altoandino de Soacha (Ariza, et al., 2010), los cerros occidentales de Chía (Cortés-S., 2003), la cuenca alta del río Bogotá (Cortés-S., 2008), los cerros Orientales (Cantillo-Higuera & Gracia, 2013) y Sumapaz (UDFJC & IAvH, 2015). En contraste, la comunidad C1 no compartió muchas especies con la comunidad C3 ni con los bosques altoandinos mencionados, pero sí mostró ciertas similitudes con el bosque andino bajo de los cerros occidentales de Chía (Cortés-S., 2003) y de la cuenca alta del río Bogotá (Cortés-S., 2008), es decir, sí estuvo más relacionada con la flora de la parte plana de la sabana de Bogotá. Por el otro lado, la plantación de Pinus patula (C2) parece tener una composición única dada la altitud a la que se encuentra (3.226 m s.n.m.), pues las pocas especies que tiene son típicas de arbustales y pajonales de páramo, frente a las especies de bosque establecidas a menor altitud (Bohórquez, 2013; Rojas-B., 2017). Estos resultados muestran que no solo están presentes en el área de estudio las especies y taxones esperados, sino que aparecen en las configuraciones esperadas de composición y estructura (a veces con menor abundancia), todo ello a pesar de las actividades económicas que allí se desarrollan. Por ello es necesario evaluar a una escala regional si la vegetación silvestre y las actividades económicas efectivamente pueden coexistir manteniendo altos niveles de diversidad y funciones ecosistémicas.
Vale la pena resaltar que en la mayoría de los estudios mencionados se describen las comunidades bajo una premisa florística independiente de las variaciones estructurales que se puedan encontrar en su interior o, al contrario, asumiendo como una comunidad diferente cada uno de los estados de desarrollo estructural encontrados independientemente de su composición. Esto podría dificultar la planificación de acciones de restauración basadas en este tipo de información. Aunque recientemente se ha argumentado que la estructura de la vegetación es el aspecto más relevante (McAlpine, et al., 2016), es fundamental partir de un conocimiento de la flora local para seleccionar aquellas especies que sean claves y así poder emprender el restablecimiento de la estructura de la vegetación (Török, et al., 2018). En este sentido, solamente en los estudios fito-sociológicos de Cortés-S. (2003; 2008) y Cantillo-Higuera & García (2013) se utilizó una aproximación similar a la del presente estudio, con una jerarquía de comunidades anidadas en un marco florístico, cada una con sus propios atributos de composición y estructura; pero incluso así, en los tres estudios habría que unificar la sintaxonomía propuesta y enmarcarla en un esquema sinfotosociológico que relacione las diferentes asociaciones con un proceso sucesional. Por lo pronto, la propuesta metodológica aquí empleada puede resultar útil para la misma tarea, evidenciando de forma más sencilla patrones generales de composición y estructura ampliamente documentados de las comunidades vegetales y permitiendo profundizar en detalles de la variación de su estructura cuando se ven sometidas a perturbaciones (como especies dominantes, cobertura de estratos o epifitismo), los cuales pueden ser claves para adoptar decisiones sobre los diferentes aspectos de su conservación.
Conclusiones
Se confirmó que las comunidades florísticas pueden presentar varios estados de desarrollo estructural, probablemente debido a las perturbaciones de origen agrícola; dichos estados son más evidentes en aquellas comunidades boscosas que representan el mayor nivel de complejidad estructural. Asimismo, se encontró que los parches de vegetación todavía exhibían un patrón de riqueza representativo de la región a pesar de las transformaciones sufridas, pues la mayor riqueza se observó en el bosque altoandino y decreció en los arbustales y pajonales de páramo a medida que se incrementaba la altitud, tal como predicen los modelos para las montañas tropicales. También se confirmó la similitud en la composición y la estructura con respecto a la registrada en las montañas circundantes de la sabana de Bogotá, y se presentó información sobre posibles variantes en presencia de especies exóticas como Pinus patula o Ulex europaeus. Se encontraron comunidades con estados de baja complejidad estructural debido a las perturbaciones que provocan las actividades agropecuarias de la cuenca, aunque en esas mismas también se encontraron estados de alta complejidad, lo que sugiere que los ecosistemas silvestres podrían llegar a coexistir en un contexto rural de uso de la tierra. La propuesta metodológica presentada puede usarse para evidenciar de forma sencilla patrones generales de riqueza y composición de la vegetación, así como profundizar en detalles de su variación estructural en situaciones de perturbación que son claves para tomar decisiones sobre su conservación.
Se recomienda incrementar el esfuerzo de muestreo en la cuenca o en sus zonas aledañas para abarcar una mayor variación florística y estructural de las comunidades identificadas. En este sentido es importante documentar con mayor detalle las comunidades con presencia de especies exóticas para hacer un seguimiento de su posible comportamiento invasivo. También deben incluirse variables detalladas del suelo y la topografía, y hacer mediciones a lo largo del tiempo para corroborar las comunidades y los estados de complejidad propuestos.

