











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
PermalinkLivestock production is the main economic activity of the Colombian piedmont eastern plains (Vergara, 2010), which is based mainly on pastures of Brachiaria spp. In the Araucano piedmont, one of the introduced grasses of greater adoption is Brachiaria decumbens Basilisk (synonymous, Urochloa decumbens Stapf R. D. Webster) for its adaptation to acid soils and response to fertilization. This pasture is managed in rotational grazing without using specific criteria to determine its optimum defoliation frequency (Rincón et al., 2008).
In the last two decades, considering pasture growth dynamics, significant progress has been made adjusting the management of pastures regarding the optimum defoliation frequency. For Lolium perenne, the use of leaves per tiller has been widely studied and adopted by farmers (Fulkerson and Slack, 1994; Fulkerson and Donaghy, 2001; Solomon et al., 2017). This criterion was developed by the observation that, in this specie, tillers maintain a maximum of three green leaves, and senescence begins in the leaf closer to the ground when the fourth leaf emerges (Fulkerson and Slack, 1994). Therefore, L. perenne pastures should be harvested at this point to avoid the accumulation of dead material and maintain pasture quality.
The number of leaves per tiller as a criterion to determine defoliation frequency may be a better parameter to establish defoliation time than others such as days to defoliation, height to defoliation, or light interception. Days to defoliation may vary due to environmental conditions (temperature, light, rain), and a given number of days will be valid only under similar environmental conditions. Herrero et al. (2.000a, 2000b) suggested that pasture management should consider environmental conditions to determine defoliation frequency and that the number of leaves per tiller could be an applicable pasture management guideline since leaf appearance and pasture growth relates to environmental conditions. Pasture height (Gouveia et al., 2017) is a parameter that does not consider pasture quality that has a profound effect on animal performance. Finally, to defoliate pastures at 95% light interception has shown promising results (Pedreira et al., 2017a) but requires special apparatus to be measured that limits the use of this technology in practical conditions.
In tropical pastures, Gomide et al. (2007) suggested that for Panicum maximum, the number of leaves per tiller can be used to establish the resting period of a pasture. These authors found that this specie harvested with fewer leaves per tiller (2.5) was more productive than those harvested with a larger number of leaves per tiller (4.5) due to more frequent defoliation that stimulates pasture growth. In B. decumbens, the number of green leaves that a tiller can maintain varies according to management and season. Fagundes et al. (2006) observed that tillers of B. decumbens fertilized with nitrogen maintained 5.1 green leaves per tiller while unfertilized maintained 6.7. Similarly, Da Silva et al. (2012) found that unfertilized B. decumbens keep up to 6.7 green leaves while fertilized 5.2. Santos et al. (2011) observed that the number of green leaves in a tiller was lower in winter (2.9) than in summer (5.4). Santos et al. (2011) observed that this grass managed under continuous grazing and defoliated to maintain a constant (25 cm) or variable 15- to 25-cm height, tillers keep close to five green leaves during spring and summer but only 2.9 during winter. Therefore, environmental conditions may alter the number of green leaves that a tiller supports, and research should be performed under particular conditions to establish appropriate harvesting time for this species using this criterion.
There is no information to infer the appropriate number of leaves per tiller in B. decumbens to avoid senescence of leaves in Araucano piedmont; therefore, the objective of this study is to generate defoliation parameters for B. decumbens, considering the number of green leaves that a tiller can maintain.
MATERIALS AND METHODS
Two studies were conducted to determine the optimum defoliation frequency of B. decumbens pasture using the number of green leaves per tiller as a criterion.
Study 1
Ten cattle farms were selected from the Araucano piedmont (municipality of Tame in the department of Arauca) to determine the moment when the first leaf that emerged after grazing begins its senescence. All the farms have B. decumbens cv. Basilisk pastures. Climatic conditions were similar among farms. The average temperature was 25.3 oC, a dry season of four months (December to March), a rainy season from April to November with total accumulated rainfall of 2,047 mm in 2015 (Figure 1). Soils of the region are strongly acid (<4.5), with a high aluminum saturation (2.4 meq 100g-1), low cation exchange capacity (<9 meq 100g-1), low content of organic carbon(<1.3 %), and P (<20 mg kg-1) (IGAC, 2017). Most farms (80%) did not use fertilization; pastures were rotationally grazed and rainfed. Stocking rates vary among farms due to differences in the primary productivity of pastures. From September 3 to October 10, 2015, 350 tillers were collected in each farm; seventy tillers per each maturity stage (2, 3, 4, 5, or 6 leaves per tiller).

Figure 1 Rainfall (mm) in the Araucano piedmont during 2015 (Based on historical data collected in the region, IDEAM, 2020)
The number of leaves per tiller was established, counting the number of leaves from the bottom of the tiller to its top. Data were recorded considering expanding, expanded, senescent, and dead leaves: a) Expanding leaf: the blade of the leaf was not fully exposed, b) expanded leaf: the ligule was visible, c) senescent: less than 50% of the leaf blade showed signs of senescence. d) Dead: more than 50% of the leaf blade showed signs of senescence.
A pooled sample by farm of leaves per tiller (2, 3, 4, 5, or 6), were dried at 65 °C for 48 hours and ground in a Wiley type blade mill to determine de nutritional quality of the tiller with a different number of leaves. Samples were analyzed for dry matter (DM), ash, crude protein (CP) (AOAC, 2012), neutral detergent fiber (NDF), acid detergent fiber (ADF) (Van Soest et al., 1991) and in vitro digestibility of dry matter (IVDDM) (Tilley and Terry, 1963).
Study 2
This experiment was conducted in a farm located in the municipality of Tame (6°52’36.55” N, 71°68’96.23” W), in an established pasture of B. decumbens cv. Basilisk. The study began on September 15 and ended on December 15, 2016. Rainfall was 256 mm during the experimental period, and the average temperature was 26.7 oC. However, the rain was not evenly distributed and did not rain in the last 20 days of the experiment. According to soil composition (Table 1), dolomite lime (300 kg ha-1), diammonium phosphate (250 kg ha-1), urea (11 kg ha-1), and sulfur (18 kg ha-1) were applied in an area of 250 m2. The selected pasture area was defoliated before fertilizing at 25 cm height on September 27, 2016.

The treatments consisted of three defoliation frequencies of the pasture, taking as a criterion the number of leaves per tiller (3, 4, or 5 leaves). Nine plots of 5×4 m were established as experimental units assigning one of the treatments randomly to each plot (3 repetitions per treatment). The methodology proposed by Fulkerson et al. (2010) was used to determine the number of leaves per tiller. Every 2 to 3 days, 15 tillers were counted in each plot, considering the time of harvest when at least 70% of them had the number of leaves of the treatment. The plots were defoliated in two consecutive periods at 25 cm height, as recommended by Rincon et al. (2008). The two periods for each treatment were evaluated without interruption.
Morphogenic characteristics were measured by selecting 15 tillers in each plot. These tillers were tagged and followed throughout the experiment every 2 or 3 days. In each of the 15 tillers, data were recorded regarding a) Days of the appearance of leaves, b) length of each leaf from its appearance until ligule exposition, c) alive and dead leaves. According to Curcelli (2009), recorded data were used to estimate leaf appearance rate (LAR), phyllochron, leaf elongation rate (LER), the final length of the leaf (mean length of all leaves in a tiller), and the number of dead leaves (DL)
Pasture undisturbed height was estimated by placing a transparent sheet on top of the pasture and measuring with a ruler the distance from the soil to the sheet. Ten observations per plot were taken every 7-8 days until harvest.
Pasture growth and dry matter yield were determined by harvesting in each plot an area of 0.25 m2 every 7 to 8 days at a 25 cm height. A sample of the cut pasture of each plot was dried in an oven (60 °C for 48 hours) to estimate dry matter and nutritional quality by wet chemistry as described above (Study 1). Dry matter and nutrient yield were estimated considering the area of the plot and the nutrient composition of the pasture.
Pasture components were measured at each defoliation frequency by harvesting a pasture area (25×25 cm2) at ground level in two points of each plot. The harvested biomass was separated into three components: green leaves, green sheaths and stems, and dead material. Then, dried in an oven (60 °C for 48 hours) to estimate dry matter and nutritional quality (CP, NDF, ADF, and ash) by near-infrared reflectance spectroscopy (NIRS), employing a DS2500 FOSS equipment calibrated against wet chemistry (Ariza-Nieto et al., 2017). The IVDDM was determined by the Tilley and Terry (1963) method.
Statistical analysis
Study 1
A complete randomized block design was applied using the GLM procedure of SAS®. Sources of variance were the number of leaves per tiller (2, 3 4, 5, 6), farms (Blocks), and the experimental error. Statistical assumptions for linear models were achieved. An analysis of Variance (ANOVA) was performed at a significant level of 5% (P<0.05), and multiple comparisons among means were performed using the Tukey test (Martinez et al., 2011a).
Study 2
A complete randomized design was used. Dry matter and nutrient yields, nutritional quality, morphogenic characteristics, and structural components were analyzed using the GLM procedure of SAS®, performing an analysis of variance with the level of significance of 5% (P<0.05). Source of variance were the number of leaves per tiller (3, 4, 5), defoliation (1, 2), and experimental error. Tukey test was used to determine differences between mean treatments (Martinez et al., 2011a).
Dry matter accumulation rate (kgDM ha-1 d-1) of each treatment (leaves per tiller) was determined by simple linear regression (Growth days vs. DM yield) and slopes of each treatment compared using the student T-test. The assumptions of the models were homogeneous experimental material and the experimental error as an independent random variable with normal distribution, mean 0, and common variance. Shapiro-Wilk test was computed to determine the normality (Martinez et al., 2011b).
RESULTS AND DISCUSSION
Study 1
In the first study, the evaluation of a large number of tillers (3,500) in ten cattle farms suggests a similar behavior of leaves senescence in a tiller in the pastures of B. decumbens regardless of management and environmental conditions among farms (Figure 2). It also suggests that the number of leaves per tiller is a valid parameter to give recommendations of the optimum defoliation frequency for the Araucano piedmont region. In each tiller, the senescence started in the leaf that appeared first (leaf closest to the ground), and leaves of a tiller remained green until the third leaf. From leaf 4, some senescent leaves were seen in the tillers, although their proportion was low (2.3%). From the fifth leaf, dead leaves appear, but also in a low proportion of tillers (3.3%). From leaf 6, most of the tillers had senescent or dead leaves. Therefore, data suggested that the senescence of the first leaf began in most tillers when these reached the fifth leaf since, at a stage of six leaves, most tillers had at least one senescent or dead leaf (Figure 2). Therefore, defoliate B. decumbens at 6 leaves per tiller would seem inappropriate. At this point, the additional leaf would imply the senescence of the first leaf, which results in a constant number of green leaves (5) in a tiller and a decrease in the nutritional quality of the pasture. From leaf 6, any additional leaf will result in an accumulation of senescent and dead leaves from there onwards.

Figure 2 Proportion of tillers with green, senescent, and dead leaves in 10 farms of the Araucano piedmont. Error bars represent the standard deviation (SD) of the mean.
Additionally, the presence of dead leaves in the pasture would be undesirable since bovines prefer green leaves over dead leaves (Rattray et al., 2007), which are located in the lower layer of the canopy, which difficult their intake. Additionally, these leaves have a lower nutritional quality than green leaves (Gomide et al., 2001). Defoliating pastures with a greater number of leaves per tiller (>5 leaves) would have other disadvantages. Research on tropical pastures has found that increasing the rest period (number of leaves per tiller) in a pasture results in pastures with greater height (Maranhão et al., 2010) which causes elongation of stems due to the elevation of the apical meristem in search of light and increases the shade of leaves near to the ground inducing their senescence (Lara and Pedreira, 2014).
It may be advisable to defoliate the pasture with fewer leaves per tiller (<5), considering the nutritional quality. In this study, maturity measured as the number of leaves per tiller was related to a linear manner with most of the nutritional parameters of the tillers. In particular, the variations in IVDDM (r2=0.40) and ADF (r2=0.55) were more closely associated with the change in the number of leaves per tiller. In vitro digestibility of dry matter (IVDDM) of tillers decreased with the increase in the number of leaves per tiller, being similar between tillers with 2 or 3 leaves and tillers with 4, 5, or 6 leaves. Crude protein decreased 2.9% from 2 to 6 leaves per tiller. NDF and ADF increased with the number of leaves per tiller but were similar among tillers with 2 or 3 leaves and 4, 5, or 6 leaves (Table 2).
Table 2 Nutritional quality of Brachiaria decumbens tillers defoliated with 2, 3, 4, 5, or 6 leaves in pastures of ten cattle farms of the Araucano piedmont.

The changes in nutritional quality were slow, and quality was similar between two consecutive leaves (Table 2). A larger proportion of senescent and dead leaves in the tillers (tiller with 6 leaves) did not accelerate the loss of nutritional quality, probably due to their relatively low proportion compared to the total green leaves in the tillers (Figure 3).
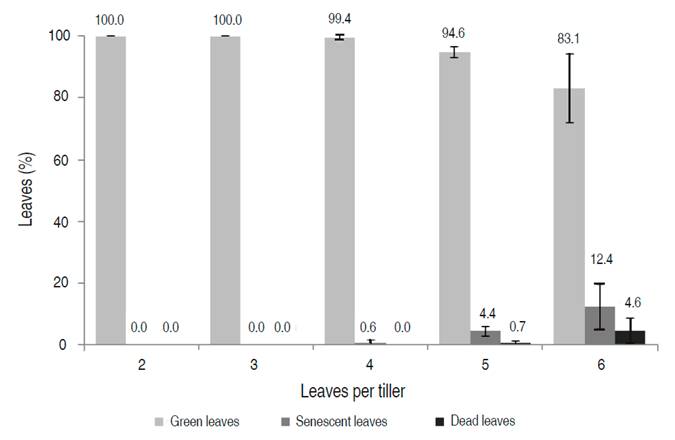
Figure 3 Percentage of green, senescent, and dead leaves from total leaves in tillers of B. decumbens in 10 cattle farms of the Araucano piedmont. Error bars represent the standard deviation (SD) of the mean.
The effect of increasing leaves per tiller on the reduction of the nutritional quality of the pastures has been documented in both temperates (Solomon et al., 2017) and tropical grasses (Agnusdei et al., 2011). In Chlorys guayana, Agnusdei et al. (2011) observed that increasing the number of leaves in a tiller and leaf stage reduced the in vitro digestibility of dry matter that was explained mainly by a reduction in the digestibility of leaf neutral detergent fiber.
In the present study, the highest nutritional quality was observed when tillers had two leaves. Defoliating the plants at two leaves per tiller increases defoliating frequency, and some authors point out that increasing defoliation frequency could compromise pasture recovery and that pastures may be depleted over time (Chilibroste et al., 2000). However, studies with tropical pastures suggest that higher defoliation frequencies (Rizato et al., 2019), or a lower pasture height (Afonso et al., 2018) generate a higher pasture quality without sacrificing production. Consequently, higher yields of meat or milk per hectare at the higher nutritional quality of the pasture may be expected (Congio et al., 2018). Therefore, optimum defoliation frequency for these pastures may be achieved with a lower number of leaves per tiller (higher quality) as long as the pasture nutrients yield is not compromised.
Study 2
In study 2, each additional leaf in a tiller required 8 to 9 days to appear. Therefore, the treatment of 3 leaves per tiller requires 16 to 18 days less than 5 leaves to be ready for defoliation (Table 3). Although pasture DM yield increased as the number of leaves in tillers increased from 3 to 5 leaves, results suggested that pasture may be defoliated at 4 or 5 leaves per tiller without sacrificing DM yield. Similar to that observed in the first study, IVDDM and CP decreased as the number of leaves per tiller increased, but the magnitude of these changes depends on defoliation (interaction). Fiber components (NDF, ADF, and lignin) increased as the number of leaves increased, but changes per each additional leaf were small (1 to 2%) (Table 3). Therefore, defoliating pastures of B. decumbens with 4 leaves per tiller resulted in an increase of the yield of digestible nutrients and CP as compared with 3 leaves per tiller and had the largest yield of NDF and ADF (Table 3). The pastures defoliated at 4 leaves per tiller had the largest DM accumulation rate (143.5 kgDM ha-1 d-1) while those harvested at 3 (114.4 kgDM ha-1 d-1) or 5 (92.6 kgDM ha-1 d-1) leaves per tiller were similar to each other (P<0.05). Fernández et al. (2001) reported that growth and accumulation rates might be reduced with the increase of age in tropical pastures. As pastures mature, the net rate of photosynthesis is reduced (Lopes et al., 2013), probably due to shading that occurs when the number of leaves per plant increases, reducing the assimilation of CO2 (Hernández et al., 2000).
Table 3 Days to harvest, dry matter yield, nutritional quality, and nutrient yield of pastures of Brachiaria decumbens defoliated at 3, 4, or 5 leaves per tiller (LT) in two consecutive defoliations (DF).

The plant adapts to lower penetration of light, generating an increase in the length of stems by the action of the phytochrome, providing a greater height in the pasture. This method of avoidance of shading occurs by raising the apical meristem of tillers that seeks light to ensure their production and survival (Smith, 1982). This would explain the lower performance of pastures harvested at 5 leaves, but not the lower performance of pastures harvested at 3 leaves when compared with pastures harvested at 4 leaves per tiller. However, the observations of the morphogenic characteristics would seem to favor the treatment of 3 leaves per tiller that in the second defoliation had a shorter phyllochrone, a faster leaf appearance rate, and a shorter height. This treatment also had a faster leaf elongation in the first defoliation (Table 4).
Table 4 Pasture height, leaf growth characteristics, and structure of Brachiaria decumbens pastures defoliated at 3, 4, or 5 leaves per tiller (LT) in two consecutive defoliations (DF).

Skinner and Nelson (1995) suggested that when the size of the tiller is short (less height), the leaves make a shorter route for its emergence at the end of the sheath. In the second study, the height was linearly and negatively related to the leaf appearance rate and to the phyllochron, which suggests that the longer the stem, the greater the length of the sheath, the appearance of new leaves becomes slower. Menezes et al. (2019) reported that the higher defoliation frequency increases the leaf expansion rate in Brachiaria brizantha; therefore, the tiller with fewer leaves has a higher leaf expansion rate. This behavior can be attributed to a greater renewal of tissues. More frequent defoliations facilitate a greater elimination of tissues and allow greater production of leaves, possibly by better penetration of light (Marcelino et al., 2006). Such behavior was observed in the first defoliation where the faster leaf elongation rate was observed in tillers with a lower number of leaves, but not in the second one, where no differences among treatments were observed. However, in the second defoliation, leaf appearance was larger for tiller with fewer leaves (Table 4).
Some literature favors a lower number of leaves per tiller as the criterion to defoliate the pasture. Martuscello et al. (2017) mention in their research that light determines leaf appearance rate, the tillering, and the leaf expansion rate. As the leaf area index increases, the uptake of light radiation by the plants’ decreases, resulting in delayed emergence of tillers and leaves and therefore decrease in the photosynthetic rate (Brown, 1984). However, a higher frequency may have limits since high frequencies may deplete storage carbohydrates. Fulkerson and Donaghy (2001) showed that in ryegrass requires at least two leaves per tiller to have an adequate subsequent regrowth. On the other hand, Pedreira et al. (2017b) reported that more frequent defoliation (base in a 95% light interception) accumulates more underground carbohydrates than less frequent defoliation in B. brizantha.
In the present study, the final length of leaves did not change due to defoliation treatment (Table 4). In tropical grasses, Da Silva et al. (2012) associated the final length of the leaves with nitrogen fertilization and Pontes et al. (2003) with residual height, aspects that were uniform among treatments. However, differences in final length were found between defoliation periods where the first defoliation presented longer leaves (Table 4), suggesting that changes in environmental conditions may affect leaf growth. Santos et al. (2011) observed that during winter, leaves were shorter. In the present study, temperatures were slightly lower for the second defoliation (0.5 to 1.0 °C) that may explain shorter leaves for the second defoliation.
The proportion of components (leaves, stems, dead material) of the pasture did not change due to defoliation treatment (number of leaves per tiller). Green leaves and green stems represented close to three-quarters of dry matter while dead material (leaves and stems) one fourth. Green stems were the largest proportion of dry matter (Table 4). Gouveua et al. (2017) suggested that more frequent defoliation reduces the proportion of stems and dead material. In the present study, the proportion of stems and dead material was similar among treatments. Since avoiding the senescence of leaves was one of the criteria used in the selection of defoliation treatments (number of leaves per tiller), it could be expected that the proportion of dead material would be similar among defoliation treatments. This result would indicate that within the range studied (3 to 5 leaves per tiller), there would be no significant differences in the proportion of pasture components, which in practical terms would imply that the pasture could be managed among these ranges of the number of leaves per tiller without changing these parameters.
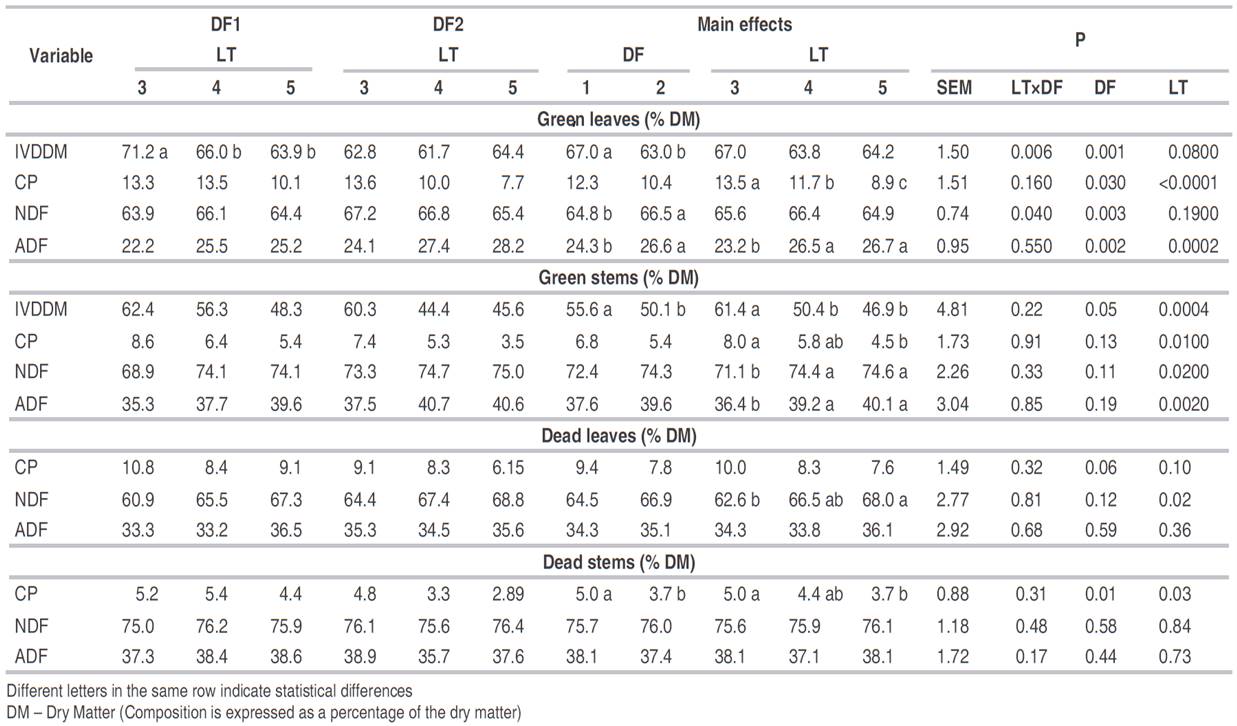
In the second study, defoliating the pasture of B. decumbens at a lower number of leaves per tiller improved its nutritional quality, similar to the results observed in the first study. Crude protein was increased in green leaves, green stems, and dead stems as the number of leaves decreased at defoliation. IVDDM increased as the number of leaves decreased in the tiller for the stems but varied among defoliations for green leaves. The concentration of NDF in the leaves was similar among treatments but increased in the green stems, and dead leaves as the number of leaves increased in a tiller (Table 5). Pedreira et al. (2017a) observed that more frequent defoliation resulted in a higher CP and in vitro organic matter digestibility in this grass in most conditions. However, stubble height and season may modulate the response of the pasture. Merlo-Maydana et al. (2017) also reported a positive effect of harvesting Brachiaria pastures at younger ages on the nutritional quality. In the present study, IVDDM decreased more rapidly in stems than in leaves because of defoliating the pasture with a greater number of leaves per tiller. Stems had an acceptable digestibility (61.4%) when harvested at earlier stages, while leaves had an IVDDM almost constant (Table 5).
Table 5 Nutritional quality of green leaves, green stems, dead leaves, and dead stems of Brachiaria decumbens pastures defoliated at 3, 4, or 5 leaves per tiller (LT) in two consecutive defoliations (DF).
The results of the present study suggest that harvesting pastures with fewer leaves per tiller would improve nutritional quality in both leaves and stems. Although cattle are highly selective and prefer leaves over stems (Santos et al., 2013), harvesting pastures at a younger age with stems of better quality could have significant impacts on pasture digestibility and intake, as these represented an important part of the plant in B. decumbens. Additionally, the results of leaf expansion and appearance rate would also favor harvesting the pasture at a lower number of leaves per tiller. However, the best performance in DM accumulation rates and nutrient yield was observed when pastures were defoliated at 4 leaves per tiller. The interactions observed in some variables between the first and the second defoliation, would suggest that it is recommended to repeat experiments of this nature with longer experimental periods where the structural changes in the plant caused by defoliation frequency are consolidated.
CONCLUSIONS
In pastures of B. decumbens at the Araucano piedmont, senescence of leaves does not start uniformly in all tillers. However, most of the tillers (65%) present senescent or dead leaves when reaching the sixth leaf, and their nutritional quality decreases. Green stems represent a large portion (43.9%) of the pasture and losses its nutritional quality faster than leaves. Defoliating B. decumbens with fewer leaves per tiller, with stems of better quality, improves pasture DM digestibility. However, defoliating the pasture with fewer leaves per tiller (3 leaves) reduces nutrient yield. Therefore, the evidence from this study suggests that B. decumbens pastures should be harvested at 4 leaves per tiller.

