











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares en
SciELO
Similares en
SciELO  Similares en Google
Similares en Google 
 Permalink
Permalink
In the Tropical High Andes [3,500-4,500 meters above sea level (masl)] only a few crops and highland grasses can grow under extremely harsh environmental conditions characterized by frosts and high levels of UV radiation (Zhang et al. 2016). One such crop is Lepidium meyenii Walpers (Brassicaceae), also known as maca, traditionally grown in the Puna of central Peruvian Andes. The latter is an area characterized by plateaus and intermountain basins, as well as high spatial and temporal climatic variability (Rolando et al. 2017). Because maca hypocotyls are high in nutrients and secondary metabolites with useful pharmacological properties (Wang et al. 2007), their cultivation has been intensified in the last 20 years to meet consumer demand in both local and export markets. Climate change, though, might compromise maca production and have a negative economic impact on this predominantly family farming activity.
Among abiotic challenges associated with climate change, water restriction and high temperatures have the most negative impact on crop productivity and the availability of arable land (Fahad et al. 2017; Restrepo-Díaz et al. 2021). Climate change manifests in the Tropical High Andes through an increase in temperature (Vuille et al. 2015) and a delayed and shortened rainy season, with less frequent but more intense precipitation (Thibeault et al. 2010; Giráldez et al. 2020). Delay and shortening of the rainy season are likely to impose water restrictions on crops, either at the beginning of the growth season, if sowing takes place at the regular dates, or later, if sowing is deferred.
Seed germination and early seedling growth are the most sensitive stages to factors such as temperature and soil moisture in several crop species (Farooq et al. 2009). Interaction between temperature and water availability strongly affects these early stages of plant development, ultimately influencing crop productivity (Mahalingam 2015). Noteworthy, below ground crops of the Andes may have reduced yields when exposed to drought during early plant growth, as has already been shown in specific potato cultivars (Tourneux et al. 2003).
As for maca, aside from recent efforts to understand its tolerance to the adverse conditions under which it grow (Shi et al. 2019; Huarancca-Reyes et al. 2020; Wang et al. 2020), little is known about the effects of drought on crop cycle, in particular on early growth stages. It is possible that the water restriction present in the germination and emergence stages could be generating changes in the characteristics of the seedlings that may be influenced by other factors such as temperature. Therefore, the purpose of this study is to evaluate the effects of water restriction on germination and seedling establishment of L. meyenii in the context of warmer temperatures, as an approach to possible climate change scenarios for this crop in the currently cultivated areas. The results of this study are important because they will not only contribute to the understanding of maca’s response to water restriction but also to have a better idea of how climate change would be affecting high mountain agriculture.
MATERIALS AND METHODS
Plant material
Six seed accessions, three from Junín district (coordinates: -11.0604, -75.9739; elevation: 4,215 masl) and three from Chupaca district (coordinates -12.2011, -75.3870; elevation 3,990 masl), were obtained from local maca producers. These accessions were selected from a bigger collection due to their best performance in pilot germination trials, in addition to being representative of the districts of Junin and Chupaca, two main maca production areas. Seeds were duly cleaned, weighed, and stored at 4 °C until use.
Experimental design
An experiment was conducted in vitro in a growth chamber (BINDER KBW-400, Germany), with a three-way factorial design (seed accession, water restriction treatment and temperature). To establish regimes that compose a reasonable scenario of short-term temperature changes in the maca-producing areas of the Junín region, the temperature records of the period 1954 to 2014 were examined (SENAMHI 2020). The maximum mean temperature above 4,000 masl rose from 16 to 18 °C, whereas at lower altitudes (3,000 to 3,600 masl) it went up from 18.5 to 20 °C. As for the minimum temperature, it rose from 2.5 to 3.7 °C on average at both altitudes. Based on these observations, two temperature regimes were set concerning maximum temperatures: a "current scenario", at 15 °C; and a "warmer scenario", at 20 °C, keeping the same minimum temperature (4 °C). All experimental groups were subjected to both temperature conditions under a 14:10 hour scheme (cold:warm), corresponding also to the dark:light periods. Daylight fluorescent tubes Color 865 (Philips) provided light during the 10-hour high-temperature period.
Germination conditions
Seeds were surface sterilized in 70% ethanol for 5 min and in 2% sodium hypochlorite with 0.05% Tween™ (Thermo Scientific) for 15 min, and finally washed three times with distilled water.
They were transferred to sterile 25x200 mm culture tubes (5 seeds/tube) containing 20 mL of solid Murashige and Skoog medium [34.3 g L-1 and Phytagel (4 g L-1)], with polyethylene glycol (PEG) 8000 added as a water retention agent (Michel 1983). Three osmotic potentials were tested: -0.25 MPa (0 g L-1 PEG as control), -0.3 MPa (150 g L-1 PEG), and -0.5 MPa (250 g L-1 PEG) (Verslues et al. 2006). These PEG levels were selected based on a pilot germination trial with PEG concentrations of up to 500 g L-1, where it was observed that at 250 g L-1 of PEG, seed germination was reduced to below 50%. At least six replicates per accession were used for each osmotic treatment. The tubes were tightly sealed with Parafilm M® to avoid water losses and incubated for 52 days (where seedlings reached the 5-leaf stage) in a growth chamber under the two temperature regimes.
Germination indexes
A visual record of seed germination was kept every 4 days using radicle extrusion (≥2 mm) as a criterion. The final germination percentage (%G) was scored and the mean germination time (MGT) was calculated as follows equation 1 (Kader 2005):
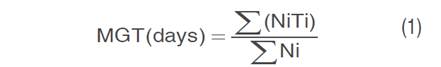
Where N is the number of seeds germinated on an ith day, and T is the number of days from the start of the experiment to the ith observation.
Seedling traits
Physiological and morphological traits were measured on day 52 after sowing as follows:
Physiological traits
Two seedlings per tube were randomly selected and used for the fluorometric quantification of Fv/Fm (OS-30P, Opti-Sciences) as an indicator of photosynthetic efficiency, and to estimate the chlorophyll content (CCM-300, Opti-Sciences).
Morphological traits
The number of leaves of two seedlings per tube was recorded. For seedling height and root length measurements, two seedlings were carefully cleaned and photographed, and their image was analyzed using ImageJ software (Schneider et al. 2012). Root:shoot ratio was calculated using the values of seedling height and root length.
Statistical analyses
A mixed-model ANOVA considering water restriction treatment with PEG (P) and temperature regime (T) as fixed effects and seed accession (A) as a random effect was used to analyze the data. When testing for the fixed factors P and T, the error term (denominator) for the F-ratio was the Mean Squares (MS) of A x P and A x T, respectively; and when testing for the P x T interaction, the error term was the MS of the A x P x T interaction. The differences between specific groups were verified by Tukey HSD tests. The analyses were performed with JMP (SAS Institute Inc.), and the graphs were created with SigmaPlot software (Systat Software, Inc.).
RESULTS AND DISCUSSION
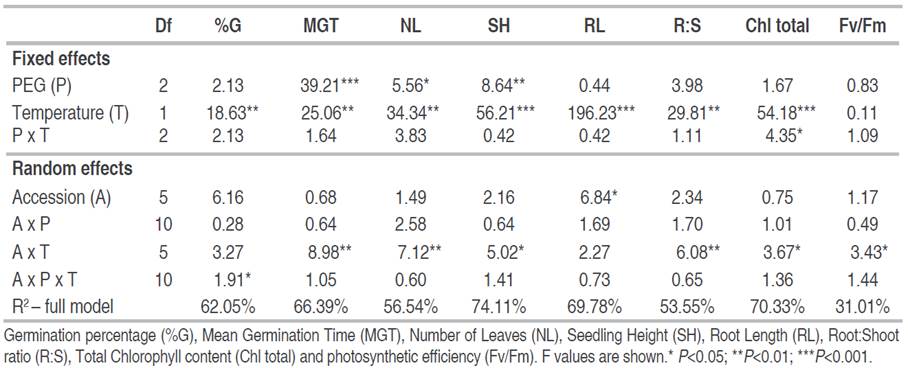
Regarding the main effects, temperature (T) affected all variables, except Fv/Fm; and water restriction treatment with PEG (P) affected the mean germination time, the number of leaves, and the seedling height. As for seed accession, it only influenced root length significantly (Table 1). In addition to the main effects, significant A x T interactions were detected for all variables except for germination percentage and root length, and significant P x T interactions for chlorophyll content. It is worth noting that PEG treatment was significant either as a main effect or when interacting with temperature (Table 1); no A x P interactions were detected among the evaluated variables, which means that water restriction affected all accessions similarly.
Table 1 Mixed-Model ANOVA for the effects of PEG treatment and temperature as fixed effects and seed accession as a random effect.
Among the water restriction effects on germination indexes and seedling traits, only the mean germination time was affected in a similar way (negatively, Figure 1B) at both temperature regimes. Maca accessions grown at 15 °C showed germination percentages below 60%, with a tendency to decrease by about 5 units with each increment in PEG concentration (Figure 1A). On the other hand, the mean germination time required to reach these percentages was more than 25 days, with a clear trend to increase at higher PEG concentrations (Figure 1B). By contrast, maca accessions that grew at 20 °C showed higher and faster germination. Their germination percentages stood above 70% regardless of PEG treatment, and despite increasing at higher PEG concentrations, the mean germination time was never longer than 22 days (Figures 1A, 1B). As for seedling traits, the water restriction effect was negative on the number of leaves at 15 °C (current scenario, Figure 1C), while negative on the seedling height and positive on the chlorophyll content at 20 °C (warmer scenario, Figure 1E, 1H). However, the effects of a higher temperature on these variables completely overcame in magnitude those caused by water restriction, with the sole exception of the photosynthetic efficiency (Fv/Fm, Figure 1C-H) that remained within optimal values (around 0.78) under all treatments. In general, the changes induced by a warmer scenario minimized the influence of water restriction and favored plant development and life cycle completion (Lin et al. 2010).
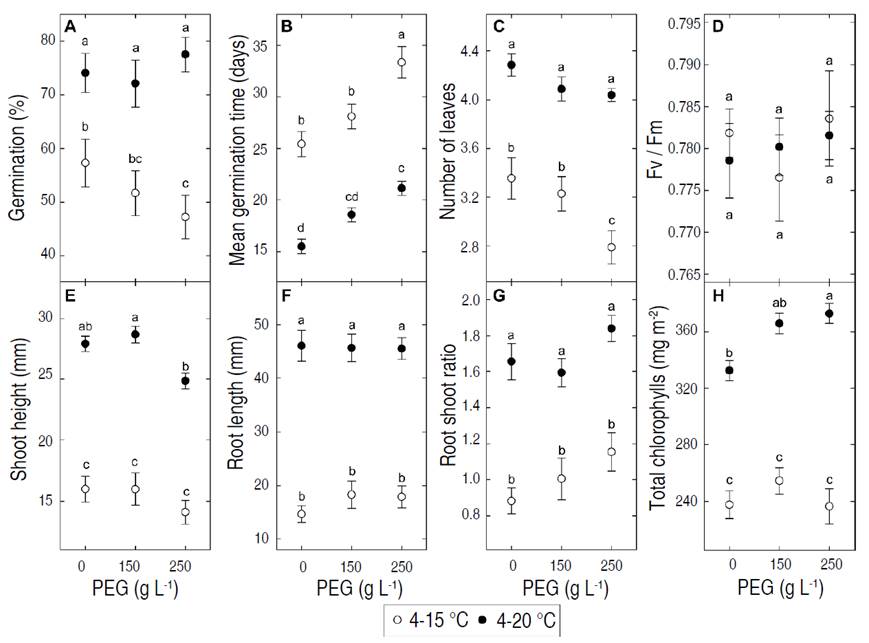
Figure 1 Effect of PEG treatments and temperature on germination percentage (%G), mean germination time (MGT), number of leaves (NL), seedling height (SH), root length (RL), root:shoot ratio (R:S), total chlorophyll content (Chl total) and photosynthetic efficiency (Fv/Fm). Black dots: 4-20 °C, white dots: 4-15 °C. Means of all accessions ± standard error are shown. Means with different letters are significantly different (P<0.05, Tukey HSD test).
PEG treatments also influenced other variables under specific conditions. Although root length was not affected, a significant reduction in seedling height was observed with 250 g L-1 PEG under both temperature regimes. While statistical significance was not reached in the linear mixed model or with individual regimes, the root:shoot ratio showed a notorious trend to increase as water restriction became more severe, regardless of the temperature conditions (Figure 1G). These results imply that when maca seedlings are exposed to moderate water deficit, they prioritize maintaining their roots’ attributes over those of the aerial structures (Poorter and Nagel 2000). Similar responses have been reported in alfalfa (Zeid and Shedeed 2006; Zhang et al. 2018) and potato (Lahlou and Ledent 2005), in the latter case with positive consequences for tuber yield.
Two observations with physiological implications deserve further comment. First, the results show that all the treatments applied were mild at most, as no indication of stress was evident from the Fv/Fm measurements. No substantial variations of the photosynthetic efficiency accompanied any of the changes in the other variables, thus suggesting that metabolic adjustments occurred in the seedlings of all accessions to keep the Fv/Fm values within the optimal range. This supports the idea that the photosynthetic machinery of maca is tolerant to moderate water restriction, despite the latter having a significant effect on plant growth. Similar results have been previously reported for peanuts (Celikkol et al. 2010). Whether the increase in chlorophyll content of maca seedlings facing moderate drought challenges in a warmer scenario contributes to preserving photosynthetic capabilities remains to be established. Sample measurements of carbon dioxide assimilation in a steady-state system shall answer this question.
Second, within the maximum temperatures evaluated and with the variables measured, maca appeared to be more responsive to temperature than to water availability. Because the values of the germination indexes and seedling traits were so overwhelmingly favorable in the warmer scenario, likely, the negative effects of water restriction at these temperatures did not have the biological constraints of those at lower temperatures. Previous studies have revealed that the effects of water restriction are more severe as the temperature moves away from the optimal range for the species’ growth and development (Mokhberdoran et al. 2009). These findings are consistent with models that predict the influence of water availability and temperature on germination and initial seedling growth (Shah et al. 2021; Khan et al. 2022). Taken together, these observations strongly suggest that the temperatures at which maca is cultivated nowadays (current scenario, 15 °C) are below the optimal range for the species’ growth and development. This notion is further supported by previous results (Valqui-Peña et al. 2021) and implies that the plant must grow at sub-optimal temperatures to yield a hypocotyl with the desired attributes. Interestingly, an earlier work by González et al. (2009) has reported an analogous phenomenon in carrots, where a sub-optimal temperature produced plants with reduced general size while simultaneously favoring growth and active allocation of carbon to the storage organ.
CONCLUSION
This study showed for the first time the effects of water restriction on maca in the context of climate change. In general, water restriction significantly affects the germination of maca seeds, having a later impact on seedling development. However, an increase in the maximum temperature could have a compensatory effect by overcoming the negative effect of water restriction. The consequences of these changes for hypocotyl development remain to be investigated, but the assumption is that under a warmer scenario, it will be compromised. In the same sense, all seed accessions evaluated behaved similarly, suggesting that the development of new maca varieties that are resistant to future environmental conditions would be necessary. Finally, its findings stress the relevance of further studying the phenological responses of Andean crops to anticipate the possible impact of climate change on their cultivation and management.

