Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
Permalink
INTRODUCCIÓN
Una plantación forestal es un área designada para la siembra de especies para la extracción comercial de la madera o como protección de servicios ambientales (FAO 2010). Cerca del 7 % de los bosques del mundo (290 millones has) corresponden a plantaciones forestales, aproximadamente la mitad establecidas exclusivamente para la producción comercial (FAO 2020). En el trópico, el 97 % de las plantaciones forestales se han establecido usando monocultivos de especies exóticas como Cupressus lucitanica Mill., 1768, Tectona grandis L.f. y especies de los géneros Pinus y Eucalyptus (FAO 2010, 2020). El establecimiento de estos monocultivos tiene efectos adversos en el funcionamiento de los ecosistemas como la disminución de la productividad del suelo, la interrupción del ciclo del agua, mayor riesgo de plagas y afectación de la biodiversidad (Hartley 2002, Bowyer 2006).
Por otro lado, se ha propuesto que las plantaciones forestales tienen el potencial de contribuir a la conservación de la biodiversidad, incluso de especies amenazadas, ya que pueden asemejar la complejidad estructural de un bosque nativo mejor que sistemas productivos como los agrícolas y ganaderos (Carnus et al. 2006, Sodhi y Ehrlich 2010, Ramírez y Simonetti 2011). Otros trabajos sugieren que las plantaciones pueden servir como una alternativa para la recuperación de la biodiversidad en zonas degradadas o servir de corredores a través del paisaje (Rusch et al. 2007, Bremer y Farley 2010, Paviolo et al. 2018).
De cualquier manera, se reconoce cada vez más a la biodiversidad como un elemento importante de la planificación y el manejo de las plantaciones, tanto al nivel de los lotes plantados, como a través de los paisajes de los cuales hacen parte (Brockerhoff et al. 2013, Begotti et al. 2018). La tendencia actual es establecer estrategias de manejo que disminuyan el efecto negativo de las plantaciones sobre los grupos biológicos presentes originalmente (Brockerhoff et al. 2013, CDB 2010), por lo que un primer paso para esto es entender cómo los grupos biológicos responden ecológicamente a estos sistemas productivos.
En general se ha encontrado una menor diversidad faunística asociada a las plantaciones cuando se compara con la de los bosques nativos de la misma región, especialmente cuando las plantaciones forestales están compuestas por especies exóticas (Brockerhoff et al. 2009, Bremer y Farley 2010, Iezzi et al. 2018). No obstante, cada grupo biológico responde de manera diferente al establecimiento de estas plantaciones exhibiendo patrones de riqueza, de abundancia y de ocupación particulares (Stephens y Wagner 2007, Felton et al. 2010, Paviolo et al. 2018). En los mamíferos se han encontrado diversos resultados dependiendo de los ecosistemas de referencia con los cuales se compare la diversidad presente en las plantaciones. Por ejemplo, en una revisión de 35 trabajos realizada por Stephens y Wagner (2007) se encontró que la riqueza de mamíferos disminuye en las plantaciones forestales cuando se compara con bosques nativos, pero puede ser mayor si es comparada con sistemas agropecuarios. Así mismo, Felton et al. (2010) encontraron en 36 trabajos de varias localidades del mundo, que la abundancia general de los mamíferos fue mayor en las plantaciones que en otros sistemas productivos y pasturas. Sin embargo, los mamíferos presentan diferentes formas de vida, hábitos, dietas y restricciones ecológicas (Feldhamer et al. 2015) por lo cual la respuesta al establecimiento de plantaciones puede ser diferente entre grupos como murciélagos, especies arborícolas y especies depredadoras.
Con miras a conocer como el establecimiento de plantaciones forestales influencia la diversidad local, se realizó una caracterización de la diversidad de mamíferos silvestres en cinco núcleos forestales comerciales ubicados en los Andes centrales y en la región Caribe de Colombia, donde se evaluaron: i) las diferencias en diversidad entre las plantaciones y los fragmentos de bosque nativos circundantes; ii) las diferencias en las abundancias relativas entre ambos tipos de cobertura; y iii) la presencia de especies amenazadas y endémicas en las plantaciones. Este trabajo se realizó para murciélagos, mamíferos mediano-grandes terrestres (en adelante mamíferos terrestres) y mamíferos mediano-grandes arborícolas (en adelante mamíferos arborícolas).
MATERIALES Y MÉTODOS
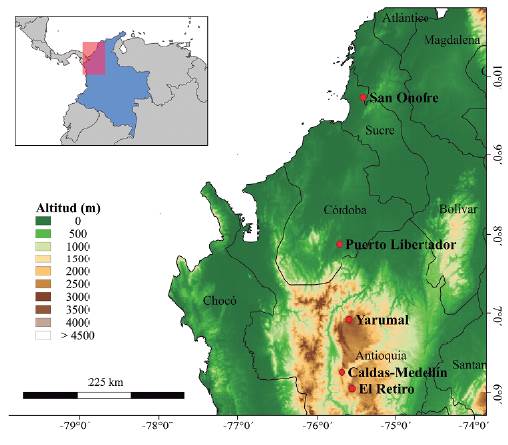
Durante los años 2013 a 2018 se visitaron cinco núcleos forestales (mosaico de lotes plantados de diferentes edades y entremezclados con fragmentos de bosque nativo y bosques ribereños) (Fig. 1). Tres núcleos correspondían a plantaciones de coníferas, compuestas principalmente de Pinus patula Schiede ex Schltdl. & Cham. y algunos lotes de Cupressus lusitanica y Pinus maximinoi H.E. More, ubicados en los Andes centrales de Antioquia (Colombia) en los municipios El Retiro, Caldas-Medellín y Yarumal entre los 1800 y 3000 m. Estos núcleos abarcaban entre 3000 y 5600 ha con el 30 % del área total en coberturas nativas. Los otros dos núcleos correspondían a plantaciones de Teca (Tectona grandis) de diferentes edades, ubicadas en Puerto Libertador (Córdoba, Colombia) con 1650 ha y 10 % de fragmentos de bosque nativo y bosques ribereños (Bosque húmedo tropical) y el otro en San Onofre (Sucre, Colombia) con 1652 ha con 40 % de fragmentos de bosque nativo (Bosque húmedo tropical), ambos entre 50 y 190 m de altitud. Cada núcleo forestal varió en su contexto paisajístico con actividades productivas diferentes en sus alrededores, y en todos los casos los lotes plantados reemplazaron potreros activos o abandonados.
Para el registro de murciélagos se seleccionaron diez a doce sitios dentro de cada núcleo forestal, cada sitio a una distancia mínima de 1 km de otro dentro del mismo tipo de cobertura vegetal (plantación o bosque nativo). La mitad de los sitios correspondieron a coberturas nativas (fragmentos y bosques ribereños) y la otra mitad a lotes plantados donde ya hubiera un dosel conformado. En cada sitio se utilizaron cinco redes de niebla de 12 m de largo, en un radio < 50m, dos de ellas elevadas en el dosel (4 a 8 m de altura), por noche durante dos noches, entre las l8:00h y las 23:00h. En total se muestrearon 85 noches en los tres núcleos forestales de coníferas (41 en plantaciones y 44 en bosques nativos) y 28 noches en los núcleos forestales de teca (quince en plantaciones y trece en bosques nativos) (Tabla 1 material suplementario). Para la identificación de las especies se siguieron como referencia a Gardner (2008) y a Díaz et al. (2016). Luego de ser identificados los animales fueron liberados cerca al sitio de captura dentro de la misma cobertura vegetal. Cada captura se asumió como un registro independiente. Para cada especie se calculó un índice de abundancia relativa IAR a partir del número de capturas dividido por el número de noches y de redes (12 m) usadas (#capturas/red-noche) (Wilson et al. 1996).
Para el registro de especies de mamíferos terrestres se utilizaron cámaras automáticas Bushnell Trophy® de 8 megapíxeles, dispuestas en caminos y carreteras forestales tanto en los diferentes lotes sembrados como en fragmentos de bosques y bosques ribereños. Las cámaras fueron ubicadas a 40-50 cm de altura desde el suelo (Díaz-Pulido y Payán Garrido 2012) y estuvieron activas entre 35 y 50 días en modo video por 24 h. Cada cámara fue instalada a una distancia mínima de 1 km una de la otra dentro del mismo tipo de cobertura vegetal (plantación o bosque nativo). Las cámaras se instalaron simultáneamente para cada tipo de cobertura en cada uno de los núcleos forestales. En total se instalaron 38 cámaras (1580 días/cámara) en los núcleos forestales de coníferas (n=17 en plantaciones, n=21 en bosques nativos) y 17 cámaras (765 días/cámara) en los núcleos forestales de teca (n=9 en plantaciones y n=8 en bosques nativos) (Tabla 1 material suplementario).
Los registros consecutivos de la misma especie con un intervalo mayor a una hora se consideraron independientes (Díaz-Pulido y Payán Garrido 2012). Para cada especie se calculó el índice de abundancia relativa IAR a partir del número de registros dividido por el número de días y cámaras utilizadas X 100 (#registros/100 días-cámara) (Díaz-Pulido y Payán Garrido 2012).
Para el registro de mamíferos arborícolas se realizaron recorridos de observación diurnos y nocturnos de entre 1h y 1,5h, en diez a doce sitios seleccionados en cada núcleo. Estos se realizaron en senderos y carreteras forestales tanto dentro de las plantaciones como en los fragmentos y bosques ribereños. Se utilizaron binoculares y linternas para facilitar la detección de los animales. En total se realizaron 50 recorridos en los núcleos forestales de coníferas (n=25 en plantaciones, n=25 en bosques nativos) y 32 recorridos en los núcleos forestales de teca (n=16 en plantaciones, n=16 en bosques nativos) (Tabla 1 material suplementario). Para cada especie se calculó el índice de abundancia relativa IAR a partir del número de registros dividido el número de horas de observación X 10 h (#registros/10 h) (Wilson et al. 1996). Especies como las ardillas, zarigüeyas y tamandúas, fueron registradas tanto en cámaras como en los recorridos de manera independiente, por lo cual cada registro fue tenido en cuenta ya fuera para la categoría de terrestre o de arborícola.
Para generar las listas de especies con la taxonomía actualizada se siguió la usada en Ramírez-Chaves et al. (2017). Para identificar las categorías de amenaza de cada especie a nivel nacional se siguió la resolución 1912 de 2017 (Ministerio de Ambiente y Desarrollo Sostenible 2017) y a nivel global la lista roja de la IUCN (www.iucnredlist.org). Para la condición de endemismo se siguieron las referencias Solari et al. (2013) y Ramírez-Chaves et al. (2017).
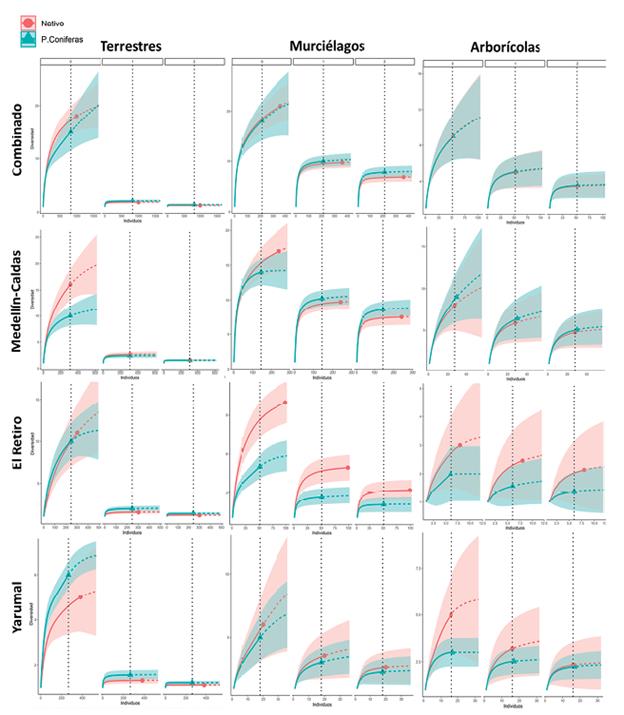
Para comparar la diversidad entre los tipos de coberturas (plantaciones y bosque nativo), estimamos la diversidad verdadera (qDα) utilizando todos los registros de cada especie. Las medidas de diversidad verdadera representan el número efectivo o equivalente de especies en una comunidad, y se calculan con base en los números de Hill (Hill 1973, Jost 2006, Moreno et al. 2011), donde los valores de "q" se conocen como el "orden" de la medida de diversidad: q=0 representa la riqueza de especies, q=1 el número de especies efectivas comunes (Diversidad de Shannon) y q=2 el número de especies efectivas dominantes en una comunidad (Diversidad de Simpson) (Jost 2006, Moreno et al. 2011). Como cada cobertura (plantaciones o bosques nativos) tuvo diferencias en el esfuerzo de muestreo, comparamos qDa mediante rarefacción al esfuerzo de la cobertura menos muestreada, y estimamos la diversidad esperada mediante extrapolación (se extrapoló al máximo posible que es dos veces el tamaño de la muestra menor) (Chao et al. 2014, Cultid-Medina y Escobar 2019) (Figs. 1 y 2 material suplementario). Comparamos los valores de qDα entre las coberturas utilizando los intervalos de confianza del 95 % en cada orden de la diversidad. Con los valores de diversidad en cada orden se construyó el perfil de diversidad para cada cobertura, representando de la estructura (riqueza y distribución de abundancias) de cada ensamblaje de especies (Moreno et al. 2011). Estos análisis se realizaron con el paquete "iNEXT" (Hsieh et al. 2016) para R.
Para comparar la distribución de las abundancias de las especies entre plantaciones y bosque nativo, se realizó una prueba Chi2 de homogeneidad (Zar 2010) entre el conjunto de abundancias relativas (IAR), para cada uno de los tres grupos de mamíferos. Para correr la prueba se escalaron los IAR a frecuencias absolutas (# de registros proporcionales de cada especie) teniendo en cuenta que los IAR ya incluían una estandarización por el esfuerzo de muestreo para cada tipo de cobertura, y se obtuvo el valor P mediante 1000 simulaciones de Montecarlo. Esto debe aplicarse cuando puede haber frecuencias esperadas bajas (< 5) (Bilder y Loughin 2014). Estos análisis se realizaron en la plataforma RStudio para R (R Core Team 2020).
RESULTADOS
Dentro de los núcleos forestales de coníferas se registraron 46 especies de mamíferos silvestres, de los cuales 36 (78 %) (murciélagos 17 (81 %), terrestres 15 (75 %) y arborícolas ocho (89 %) fueron registrados al interior de los lotes forestales plantados (Tabla 2 material suplementario). De estas, A. lemurinus y L. tigrinus están en alguna categoría de amenaza (Carretero et al. c2020, Ramírez-Chaves et al. 2016) y N. pucheranni es endémica del país (Solari et al. 2013). Del total de especies, 18 tuvieron mayores IAR al interior de los lotes plantados y 24 tuvieron IAR mayores en bosque nativo (Tabla 2 material suplementario).
La diversidad observada de los murciélagos en ambas coberturas fue similar (Fig. 2a), aunque la riqueza esperada de murciélagos de las plantaciones de coníferas fue de 23 especies y para bosques nativos de 26 especies. Cuando se compararon los conjuntos de IAR de murciélagos para ambas coberturas, se encontraron diferencias significativas (Chi2 = 88,7, P < 0,01). La diversidad observada fue mayor para mamíferos terrestres en bosques nativos (Fig. 2a) y la riqueza esperada de especies terrestres en plantaciones de coníferas fue de 21 especies y para bosques nativos de 31 especies. Cuando se compararon los conjuntos de IAR de especies terrestres en ambas coberturas, se encontraron diferencias significativas (Chi2 = 73,2, P < 0,01). La diversidad observada para especies arborícolas fue similar en ambas coberturas (Fig. 2a) y la riqueza esperada de especies arborícolas en plantaciones de coníferas y para bosques nativos fue igual con doce especies. Cuando se compararon los conjuntos de IAR de especies arborícolas para ambas coberturas (Tabla 2 material suplementario), se encontraron diferencias significativas (Chi2 = 83,5, P < 0,01).
Para los núcleos forestales de teca se encontraron 49 especies de mamíferos silvestres, de los cuales 35 (71 %) (murciélagos 17 (74 %), terrestres doce (75 %) y arborícolas siete (58 %)) fueron registrados al interior de los lotes forestales (Tabla 3 material suplementario). De estas, C. capucinus, A. griseimembra, S. oedipus y O. cf cariacou están en alguna categoría de amenaza (Ramírez-Chaves et al. 2016, Link et al. c2019, Mittermeier et al. c2020, Rodríguez et al. c2020), además, S. oedipus es endémica de la región Caribe (Solari et al. 2013). Del total de especies, trece tuvieron mayores IAR al interior de los lotes plantados y 36 tuvieron IAR mayores en bosque nativo (Tabla 3 material suplementario).
La diversidad observada de los murciélagos en ambas coberturas fue similar (Fig. 2b), aunque la riqueza esperada de murciélagos para las plantaciones de teca fue de 20 especies y para bosques nativos de 22 especies. Cuando se compararon los conjuntos de IAR de murciélagos para ambas coberturas, se encontraron diferencias significativas (Chi2 = 110,5, P < 0,01). La diversidad observada de especies terrestres fue similar en ambas coberturas (Fig. 2b), pero la riqueza esperada de especies terrestres en plantaciones de teca fue de catorce especies y para bosques nativos de 18 especies. Cuando se compararon los conjuntos de IAR de especies terrestres para ambas coberturas, se encontraron diferencias significativas (Chi2 = 50,8, P < 0,01). La diversidad observada para especies arborícolas fue similar en ambas coberturas (Fig. 2b), aunque la riqueza esperada para especies arborícolas en plantaciones de teca fue de nueve especies mientras que para bosques nativos no se esperaban especies adicionales a las siete registradas. Cuando se compararon los conjuntos de IAR de especies arborícolas para ambas coberturas, no se encontraron diferencias significativas (Chi2 = 37,6, P < 0,99).

Figura 2 Perfiles de diversidad para los diferentes grupos de mamíferos en los diferentes tipos de núcleos forestales obtenidos a partir de la rarefacción. Las barras verticales representan los intervalos de confianza del 95 %. a. Plantaciones de coníferas vs. Bosques nativos. b. Plantaciones de Teca vs. Bosques nativos
DISCUSIÓN
La mayoría de las especies de mamíferos registradas tanto para los núcleos muestreados en la zona Andina como en la región Caribe (71 y 78 % respectivamente) fueron encontradas dentro de los monocultivos forestales. Adicionalmente, la diversidad observada (q=0, q=1, q=2) fue similar entre las plantaciones y los bosques nativos, y tanto en plantaciones de coníferas como de teca se registraron especies amenazadas de extinción. Esto sugiere que estos sistemas productivos tienen el potencial de ser diversos, es decir que son usados por un número considerable de especies y pueden presentar una diversidad aproximada a la observada para las coberturas nativas de referencia, y aunque la riqueza esperada fue mayor en los bosques nativos, las plantaciones distan de ser percibidos como "desiertos verdes" (Potton 1994, Hartley 2002, Bremer y Farley 2010). Patrones similares de riqueza (q=0) han sido observados para aves en los mismos sitios de estudio, donde se han registrado hasta el 76 % de las especies totales dentro de los lotes plantados (Saravia Ruiz y Sánchez-Londoño 2016, Sánchez-Londoño obs pers). No obstante, es posible que patrones de diversidad diferentes se presenten para mamíferos y otros grupos biológicos en otras regiones geográficas y con otros esquemas de manejo forestal (Iezzi et al. 2018).
No existen muchos trabajos donde se haya evaluado como responde cada ensamblaje de mamíferos al establecimiento de plantaciones (Loyn et al. 2007, Felton et al. 2010) lo cual consideramos debe ser la aproximación más adecuada debido a la amplia variedad de formas de vida y hábitos que muestran los mamíferos en los ecosistemas tropicales (Fergnani y Ruggiero 2015, González-Maya et al. 2016). Nuestros resultados mostraron que los diferentes grupos de mamíferos analizados respondieron de manera diferente ya que los perfiles de diversidad para los murciélagos fueron muy similares entre las plantaciones forestales y los bosques nativos tanto en coníferas como en teca, mientras que para los mamíferos arborícolas y los mamíferos terrestres los perfiles de diversidad fueron más heterogéneos entre los tipos de coberturas.
La riqueza observada de especies efectivas (q=0) solo mostró diferencias significativas en algunos de los grupos como las especies terrestres y los murciélagos en teca, y el número de especies efectivas comunes (q=1) y dominantes (q =2) solo mostró diferencias para los mamíferos terrestres en los núcleos forestales de coníferas, mientras que para todos los demás mamíferos fue el mismo entre bosques nativos y plantaciones para los dos núcleos forestales. Esa similitud en la diversidad entre ambas coberturas puede deberse a la conformación de paisajes homogéneamente usados por la gran mayoría de especies (ver más adelante), y también a una posible pérdida de algunas especies especialistas o raras en las áreas como Panthera onca (L., 1758) en zonas de teca o Mazama rufina Bourcier & Pucheran, 1852 en zonas de coníferas, que ya había ocurrido debido a la transformación y fragmentación histórica sobre los bosques nativos previo al establecimiento de los proyectos forestales (Debinski y Holdt 2000, Michalski y Peres 2005).
Es claro que para la mayoría de las especies de los tres grupos de mamíferos muestreados hubo IAR mayores en las coberturas nativas respecto a las plantaciones (Tablas 2 y 3 del material suplementario) y de ahí consideramos que provienen las diferencias halladas. Iezzi et al. (2018) y Ramírez y Simonetti (2011) encontraron en bosques subtropicales, que tanto mamíferos como aves muestran abundancias considerablemente más bajas al interior de plantaciones forestales, lo cual parece ser un patrón general. Los bosques plantados son sistemas simplificados en términos de su estructura y composición y por lo tanto se esperaría encontrar ensamblajes de especies más empobrecidos puesto que ofrecen hábitat de menor calidad (Pawson et al. 2010, Simonetti et al. 2013, Begotti et al. 2018, Iezzi et al. 2018). No obstante, algunas especies mostraron lo contrario, por ejemplo, especies de murciélagos como A. lituratus, S. erythromos, S. lilium y Glossophaga soricina, así como algunas de las especies terrestres como C. thous, y L. tigrinus, y la especie semi-arborícola D. pernigra, fueron más frecuentes al interior de las plantaciones. Esto sugiere que el hábitat ofrecido por las plantaciones favorece bien sea su abundancia o su frecuencia de uso, teniendo en cuenta que estas especies tienen capacidad de adaptarse a ambientes intervenidos (Emmons y Feer 1999, Wainwright 2002). Los murciélagos podrían encontrar menos depredadores (serpientes, búhos, etc.) y gastar menos energía en su desplazamiento ya que el dosel y subdosel son menos complejos en las plantaciones, lo cual ya ha sido evaluado en otras localidades (Heer et al. 2010). Para el caso de algunos depredadores y omnívoros medianos terrestres, en las plantaciones disminuyen sus competidores y depredadores potenciales, como P. concolor Linneo, 1771 y L. pardalis Linneo, 1758, lo cual favorece su permanencia o uso en esos espacios (Sánchez-Londoño 2017, Cruz et al. 2018, Paviolo et al. 2018).
La diversidad registrada al interior de las plantaciones forestales puede ser influenciada por factores internos y externos. La proporción de plantaciones y coberturas nativas, la presencia de sotobosque al interior de los rodales plantados, la edad de los rodales, su periodo de rotación (tiempo de cultivo y cosecha), y el contexto paisajístico son aspectos que se han identificado afectan en mayor medida a la riqueza y abundancia de las especies al interior de las plantaciones (Ogden et al. 1997, Brockerhoff et al. 2003, Durán y Kattan 2005, Aubin et al. 2008). Por ejemplo, Simonetti et al. (2013) encontraron que la presencia del sotobosque, que en ocasiones depende de la edad y del manejo de los lotes plantados, afecta positivamente la abundancia de especies de mamíferos terrestres. De tal manera que especies de murciélagos frugívoros y nectarívoros (p.e. Artibeus sp, Glossophaga sp) así como especies terrestres y arbóreas herbívoras y frugívoras (Sylvilagus sp., Mazama sp., Aotus sp.) que fueron frecuentes en las plantaciones estudiadas por nosotros, seguramente se benefician de los recursos ofrecidos por el sotobosque al interior de los lotes, especialmente a los que ya se han hecho entresaca y los claros formados son colonizados por especies nativas de géneros como Cecropia, Piper, Vismia, Solanum, Ficus y Miconia entre otras (Obs. pers.).
Por otro lado, aunque el contexto paisajístico afecta la diversidad de mamíferos al interior de los núcleos forestales, esto dependerá de los requerimientos de cada especie, por ejemplo, del tamaño mínimo de área de hábitat (Loyn et al. 2007). Para plantaciones de palma de aceite Pardo et al. (2018) encontraron que la diversidad de mamíferos medianos y grandes como agutíes, zorros, venados, se ve afectada por un umbral en las proporciones del área ocupada por los remanentes nativos y el área plantada, mientras que Paviolo et al. (2018) encontraron que especies medianas y pequeñas como agutíes, zarigüeyas y pecarís, fueron afectadas negativamente en su abundancia a mayor proporción de plantaciones de coníferas en relación al bosque nativo. Especies más grandes como el puma no mostraron un efecto. Campos et al. (2018) encontraron como patrón general que las especies medianas y grandes usan la heterogeneidad del paisaje conformada en los núcleos forestales como un todo, por lo que no registraron diferencias en la diversidad de las coberturas vegetales. Si bien en nuestro trabajo tuvo un enfoque a escala de fragmentos, aproximaciones como estas a mayor escala y especie-específicas como modelos de ocupación, podrían explicar el efecto de la proporción de área cubierta por cada tipo de vegetación y la disposición de los lotes plantados sobre diferentes especies focales, y así tener un diseño más apropiado (mayor diversidad asociada) en la conformación de cada núcleo forestal.
Dos de los núcleos muestreados, ("Yarumal" de coníferas y "Puerto Libertador" de teca) se encontraban embebidos en una matriz de potreros para ganadería, lo cual los convertía en la única cobertura boscosa continua (> 3000 ha) en el paisaje. Tanto al interior de los lotes plantados como de los fragmentos nativos y bosques ribereños se registraron especies como P. concolor, L. tigrinus, B. neblina, O. cf. cariacou, y S. oedipus, entre otras, lo que sugiere que estas poblaciones dependen localmente, al menos de manera temporal, de la presencia de estos núcleos forestales. Por el alto grado de eliminación de cobertura boscosa regional las plantaciones forestales pueden actuar como refugio para muchas especies donde el contexto paisajístico así lo facilita, pueden ser sitios de paso entre grandes áreas de hábitat natural, o pueden proveer hábitat temporal (Wijesinghe y de Silva 2012). Este es un atributo que se debe considerar en los ejercicios de planeación del territorio y de conservación a escala del paisaje ya que existe el potencial de ofrecer hábitat o conectividad a determinadas especies (Paviolo et al. 2018), especialmente en regiones altamente transformadas como las regiones Andina y Caribe de Colombia (Morales y Armenteras 2013, Castaño-Uribe et al. 2013).
Cuatro de los cinco núcleos forestales estudiados (dos de coníferas en la zona Andina: "Caldas-Medellín" y "Yarumal" y los dos de teca en la zona del Caribe) se encontraban bajo el esquema de certificación FSC© (Forest Stewardship Council™) al momento de realizar este trabajo. Estos certificados forestales se crearon en respuesta a la preocupación general sobre cómo estos agroecosistemas pudieran ser uno de los impulsores de la pérdida de biodiversidad (van Kujik et al. 2009). En estos esquemas se acreditan los diferentes núcleos por adherirse a medidas de manejo más estrictas que intentan tener un mayor beneficio sobre la biodiversidad (Gullison 2003, van Kujik et al. 2009). En estos esquemas se solicita tener identificados dentro de cada núcleo forestal, altos valores objeto de conservación (AVOCs) dentro de los planes de manejo, en los cuales pueden estar las especies amenazadas o sus hábitats específicos (Stewart 2010).
Independientemente de la certificación forestal, proyectos forestales de este tipo cuentan con algunos atributos que los convierten en candidatos para ser aliados en la investigación y conservación de los mamíferos a nivel local o regional. Esto debido a que en algunos casos los núcleos forestales se han establecido en zonas previamente transformadas como los potreros, se suelen controlar la cacería y los incendios en sus predios, poseen infraestructura que facilita los desplazamientos y suelen contar con trabajadores conocedores de la diversidad local (obs. pers.). No obstante, como son sistemas productivos mediados por intereses económicos, es necesario incluir en sus planes de manejo medidas que faciliten su diseño espacial y su conectividad en el paisaje; las medidas de enriquecimiento ambiental de los lotes plantados pueden facilitar la presencia de especies silvestres, y las medidas de cosecha pueden tener menor impacto para la fauna.