Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
Permalink1 Introducción
El dengue es una infección viral transmitida, principalmente, por la hembra del mosquito Aedes aegypti que habita en todas las regiones tropicales y subtropicales del planeta. En Cali, Colombia, a pesar de los controles que las autoridades de salud dicen estar haciendo, durante el año 2013, se reportaron más de 9.000 casos de dengue, de los cuales algunos fueron graves y otros llegaron a ser letales [1]. A principios de noviembre del 2015, se reportaron más de 18.000 casos de dengue, de los cuales 82 fueron dengue grave y 19 fueron letales [2]. Estas estadísticas evidencian altos niveles epidémicos del dengue, lo cual es una amenaza a la salud pública de la región y requiere buscar políticas eficientes que integren diferentes estrategias de control sobre la población de vectores con medidas preventivas para la población humana susceptible.
Como ya dijimos al principio, el dengue no se transmite directamente de persona a persona: para la transmisión de esta infección siempre se requiere de la hembra del mosquito Aedes aegypti como agente transmisor. En efecto, para empezar un brote epidémico, es necesaria la presencia de al menos un mosquito hembra infectado o de una persona infectada en medio de una población tanto de mosquitos como de personas susceptibles de ser infectadas. En el primer caso, el mosquito infectado, al picar una persona sana puede transmitirle la infección del dengue, la cual a su vez, al ser picada por otros mosquitos hembra, puede infectar más mosquitos y estos a su vez pueden infectar más personas. Algo similar ocurre en el segundo caso: al ser picada por un mosquito hembra, la persona infectada puede infectar un mosquito, el cual, al picar otra persona sana, la puede infectar y esta a su vez infectar más mosquitos hembra.
Por otro lado, el Aedes aegypti es un mosquito urbano, ya que vive en lugares sombreados y frescos, como el interior de las casas. Su ciclo vital tiene dos fases (o estadios): una acuática (o inmaduro) y otra aérea (o maduro). La hembra pone sus huevos en las paredes de los recipientes con agua y cuando ésta alcanza la temperatura adecuada, los huevos comienzan su desarrollo y se convierten en larvas. Luego, las larvas pasan al estado de pupas y éstas finalmente se transforman en mosquitos adultos. En la Figura 1, se observa el ciclo de vida del Aedes aegypti. Todo este proceso tiene variaciones de tiempo y de eficiencia dependiendo de la temperatura del medio ambiente [3],[4],[5],[6],[7],[8], lo cual aprovecharemos para estimar algunos parámetros entomológicos de este mosquito en la ciudad de Cali.

Con base en estas características, es posible construir modelos matemáticos para analizar cualitativa y cuantitativamente las poblaciones de mosquitos hembra total, y de mosquitos y personas infectadas, lo cual permiten una buena comprensión de la dinámica del virus, por lo que son una herramienta para el seguimiento y control de la enfermedad. Sin embargo, para que esta herramienta sea realmente útil en casos concretos, los modelos deben ser ajustados a las características particulares de la región donde se quieren utilizar, lo cual se logra hallando los valores específicos de los parámetros de los modelos, tarea que se conoce con el nombre de estimación de parámetros.
En este trabajo, para el estudio de las poblaciones de personas y mosquitos infectados y la de mosquitos en la ciudad de Cali, consideramos dos modelos. El primero es una modificación del modelo depredador-presa propuesto en [9] y estudia la dinámica de la población de mosquitos hembra en presencia de un depredador de su fase acuática. El segundo es una versión adaptada del modelo Ross-Macdonald [10] y estudia la dinámica de las poblaciones de mosquitos y personas infectadas.
Dada la dificultad para obtener los datos entomológicos de los mosquitos Aedes aegypti en la ciudad de Cali, para el primer modelo, estimamos los parámetros usando los datos hallados en [6],[7] mediante experimentos controlados sobre el ciclo vital de este mosquito para determinar su relación cuantitativa con la temperatura del medio ambiente. Para el segundo modelo, estimamos sus parámetros como la aproximación de mínimos cuadrados que se obtiene al tratar de ajustar los resultados del modelo a los datos epidemiológicos del dengue reportados por la Secretaría de Salud Municipal de Cali (SSMC) durante el 2.010.
El objetivo de este artículo es la estimación, para la ciudad de Cali, de los parámetros tanto entomológicos del mosquito Aedes aegypti como los epidemiológicos del dengue involucrados en el par de modelos mencionados. En la Sección 2, describimos el modelo depredador-presa y en la Sección 3, presentamos la estimación de los parámetros entomológicos del mosquito Aedes aegypti usando los datos dados en [6],[7]. Luego, en la Sección 4, describimos el modelo de Ross-Macdonald adaptado para el análisis de las poblaciones de humanos y mosquitos infectados por el virus del dengue y, en la Sección 5, presentamos la estimación de los parámetros epidemiológicos del dengue en Cali usando la solución de mínimos cuadrados en el ajuste de los resultados del modelo y los casos reportados de dengue en la ciudad de Cali según la SSMC. Conviene notar que el estudio de la estabilidad de ambos modelos y la sensibilidad de sus parámetros estan presentados en [11],[12]. En el Apéndice, hacemos un análisis comparativo de los polinomios determinados en [6],[7], y los calculados y usados en la Sección 3 para estimar los parámetros entomológicos del mosquito para la ciudad de Cali. Finalmente, presentamos la bibliografía utilizada en el desarrollo de este trabajo.
2 Modelo depredador-presa
Siguiendo la formulación planteada en [9], consideramos un modelo donde dividimos el ciclo de vida del vector Aedes aegypti en dos estadios: inmaduro, conformado por huevos, larvas y pupas, y maduro, conformado por el mosquito adulto; además, en esta dinámica poblacional, consideramos la influencia de un depredador biológico para el estadio inmaduro del mosquito.
Más precisamente, suponemos que el crecimiento del estadio inmaduro (huevos, larvas y pupas) está dado por la capacidad de carga remanente (1-mq/Kq), la tasa de ovoposición de los mosquitos hembra adultos (ε), el número de estadios maduros del mosquito hembra (ma) y, a diferencia de [9], la fracción de huevos que eclosionan (k) . El decaimiento de este estadio está relacionado con la proporción de aquellos que tienen la transición al estadio maduro (ν), la proporción de los que mueren de forma natural (δq) y aquellos que son depredados (ϕx), para los cuales suponemos la ley de acción de masas. (Primera ecuación en [1]). Similarmente, el crecimiento del estadio maduro (hembras) depende de la transición del estadio inmaduro (νm q ) y, a diferencia de (9), solo tenemos en cuenta la fracción de hembras (f) , y su decaimiento está determinado exclusivamente por los que mueren de forma natural (δ a m a ). (Segunda ecuación en [1]).
De otro lado, el depredador presenta un crecimiento logístico y, a diferencia de [9], en este trabajo, consideramos un incremento (ψm q ) en el crecimiento intrínseco del depredador asociado a la interacción con la presa, como se planteó en [11]. (Tercera ecuación en [1]).
En resumen, tenemos el siguiente modelo (Ver análisis clásico del modelo en [11])

con condiciones iniciales m q (0) = m q 0, m a (0) = m a0 , x(0) = x 0, donde (m q 0, m a0 ) ≠ (0, 0) y las variables son:
m q : número promedio de estadios inmaduros del mosquito,
m a : número promedio de estadios maduros del mosquito hembra,
x: número promedio de depredadores de estadios inmaduros, y los parámetros están dados por:
ε: tasa de ovoposición de los mosquitos hembra adultos,
ν: tasa de transición de los estadios inmaduros a maduros,
δ q : tasa de mortalidad natural de los estadios inmaduros,
δ a : tasa de mortalidad natural de los estadios maduros,
k: fracción de huevos que eclosionan a larvas,
f: fracción de mosquitos hembra en estadio inmaduro,
K q : capacidad de carga de los estadios inmaduros del mosquito,
K x : capacidad de carga del depredador,
ρ: tasa de incremento del depredador,
ϕ: tasa de depredación,
ψ: tasa de incremento del depredador por depredación.
3 Estimación de los parámetros entomológicos
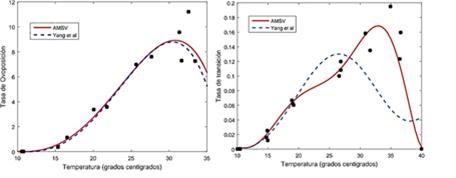
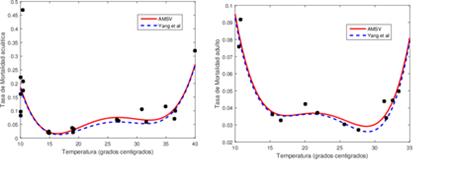
Como el dengue es una enfermedad transmitida por el mosquito Aedes aegypti y su incidencia está fuertemente afectada por la temperatura y la humedad del medio ambiente donde se desarrollan, las cuales varían estacionalmente, en [6],[7], se realizaron experimentos sobre el ciclo de vida del mosquito con control de la temperatura ambiental. Los datos así recolectados les permitió establecer relaciones funcionales de los principales parámetros entomológicos del mosquito (tasa de ovoposición, tasa de transición de estadios, tasa de mortalidad inmaduros, tasa de mortalidad maduros) y la temperatura del medio ambiente donde se desarrolla el mosquito. Los mosquitos usados en estos experimentos fueron atrapados en Marilia, ciudad situada a 350 Km al Noroeste de Sao Paulo, Brasil, razón por la cual los periodos de luz-oscuridad fueron diseñados para simular el día y la noche de esta ciudad según la temperatura (bajas invierno y altas verano). Se obtuvieron datos diarios del número de huevos, de larvas, de pupas y de mosquitos para temperaturas promedio de la cámara de reproducción entre 10 y 40 grados centígrados durante casi cuatro meses, para la fertilidad (huevos) y la edad adulta (mosquitos), y seis meses, para la fase acuática (larvas y pupas), con tres y dos observaciones para cada temperatura, respectivamente.
Con base en los anteriores datos, se estimaron los parámetros para cada temperatura, y luego, cada parámetro se ajustó a una función de la temperatura (polinomio de grado entre 3 y 8), usando mínimos cuadrados lineales, como aparece en el anexo, al final de este trabajo.
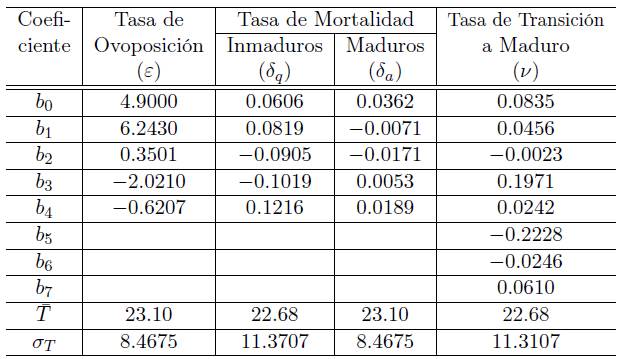
Con base en una observación planteada en [13], sobre un posible error en los coeficientes de los polinomios estimados en [6], especialmente para el polinomio correspondiente a la tasa de transición a maduro, usando las Tablas 4, 5, 8 y 10 del Apéndice de [6], para cada parámetro (tasa de mortalidad maduros, tasa de oviposición, tasa de mortalidad inmaduros y tasa de transición a maduro), se calculó el polinomio de grado m, centrado y re-escalado de la siguiente forma

que minimizara el error E dado por

donde m es el grado del polinomio determinado en [6], T¯y σ T son el promedio y la desviación estándar de las temperaturas usadas para las medidas del parámetro, N es el número de observaciones (temperaturas) hechas para el parámetro, φ j es el parámetro estimado para la temperatura j y σ j es la desviación estándar correspondiente. Los coeficientes b i , los promedios T¯ y las desviación estándar σ T de estos polinomios están dados en la Tabla 1.
Tabla 2 Temperaturas medias para Cali dadas por el IDEAM en el año 2010.
| Sitio | Temperatura | ||
| Media Mínima | Media | Media Máxima | |
| Universidad del Valle | 19.0°C | 23.9°C | 29.8°C |
| Aeropuerto A.B.A. | 18.7°C | 23.5°C | 29.6°C |
| Base Aérea M.F.S. | - | 24.6°C | - |
Para hacer el ajuste de los parámetros entomológicos del mosquito Aedes aegypti para Cali, consideramos las temperaturas media, media mínima y media máxima registradas por el Instituto de Hidrología, Meteorología y Estudios Ambientales (IDEAM) para la ciudad de Cali, en el año 2010 [14], las cuales se miden en tres sitios de la ciudad: Universidad del Valle, Aeropuerto Alfonso Bonilla Aragón y la Base Aérea Marco Fidel Suárez, y aparecen en la Tabla 2.
En la Tabla 3, se presentan los valores de cada uno de los parámetros entomológicos del mosquito calculados usando los polinomios dados en la Tabla 1 y para las temperaturas de Cali registradas en la Tabla 2.
Tabla 3 Parámetros entomológicos del modelo presa-depredador para Cali.
| Sitio | Temp. | Parámetros | |||
| ε (σε) | ν (σν) | δ q (σ δ q) | δ a (σ δ a) | ||
| UniValle | 19.0°C | 2.2 (2.8) | 0.0628 (0.0004) | 0.03 (0.01) | 0.036 (0.003) |
| 23.9°C | 5.5 (6.7) | 0.089 (0.001) | 0.07 (0.01) | 0.035 (0.004) | |
| 29.8°C | 8.8 (7.9) | 0.143 (0.003) | 0.07 (0.02) | 0.030 (0.003) | |
| Aeropuerto | 18.7°C | 2.0 (3.9) | 0.0602 (0.0007) | 0.03 (0.01) | 0.036 (0.004) |
| 23.5°C | 5.2 (6.4) | 0.087 (0.001) | 0.07 (0.01) | 0.036 (0.005) | |
| 29.6°C | 8.8 (7.9) | 0.140 (0.003) | 0.07 (0.02) | 0.030 (0.003) | |
| Base Aérea | 24.6°C | 6.0 (7,1) | 0.092 (0.001) | 0.07 (0.01) | 0.034 (0.004) |
Según estos resultados [Tabla 3], podemos decir que, en Cali,
cada mosquito hembra pone entre 2 y 9 huevos diarios (tasa de ovoposición), siendo esta tasa creciente con la temperatura.
del 6 al 14.3% de los estadios inmaduros pasa al estadio maduro, diariamente (tasa de transición), siendo esta tasa también creciente con la temperatura.
entre el 3 y 7 % de los estadios inmaduros mueren, diariamente (periodo de vida entre 14 y 34 días). Y
entre el 3.0 al 3.6 % de los mosquitos adultos mueren en forma natural, diariamente (periodo de vida entre 28 y 34 dias).
En la Tabla 3, también podemos observar que, para el rango de temperaturas y los diferentes sitios de Cali, la variación de la transición del estadio inmaduro a maduro (ν), y las mortalidades de los estadios inmaduro y maduro (δ q y δ a ) es relativamente baja (sus valores están entre 0.06 y 0.14, 0.03 y 0.07, y 0.030 y 0.036, respectivamente), mientras que la ovoposición tiene una alta variación (sus valores están entre 2.2 y 8.8). En cuanto a la desviación estándar (σ.), es baja para ν y δ a , moderada para δ q y alta para s. Lo anterior plantea, para los entomólogos, el reto de diseñar mecanismos para estimar la tasa de ovoposición con mayor precisión.
También es interesante notar que, en Cali, mientras las temperaturas altas favorecen (aumentan) la ovoposición, la transición de estadios inmaduros a maduros y la mortalidad de estadios inmaduros, disminuyen la mortalidad del mosquito adulto.
Para terminar esta parte, debemos anotar, primero, que los valores de los parámetros aquí estimados son válidos no solo para el modelo [1] sino tambien para cualquier otro modelo donde tengan el mismo significado; y segundo, que los otros dos parámetros del modelo (1), las capacidades de carga tanto de los estadios inmaduros como del depredador, pueden tomarse en forma relativa la una con la otra, según la situación que se desee modelar (Por ejemplo, ver Sección 6 en (11)).
4 Modelo Ross-Macdonald
El modelo Ross-Macdonald es un modelo compartimental basado en ecuaciones diferenciales ordinarias, que describe las interacciones entre la población de mosquitos y humanos en la dinámica de una enfermedad de transmisión indirecta. El modelo divide cada población, la de humanos y la de mosquitos, en dos compartimientos: susceptibles e infectados, y describe la interacción dinámica de los infectados. Los supuestos del modelo son:
Las poblaciones de humanos y de mosquitos se mantienen esencialmente invariables en el tiempo (son poblaciones cerradas).
Las poblaciones de humanos y de mosquitos son homogéneas en cuanto susceptibilidad y exposición.
Se ignoran los tiempos de incubación dentro de los humanos y los mosquitos.
Se ignora la adquisición gradual de inmunidad en los humanos (en Cali, circulan hasta cuatro serotipos del dengue, simultáneamente).
Se ignora la natalidad y la mortalidad en los humanos (el horizonte de tiempo es relativamente corto).
Los mosquitos no se recuperan.
No se considera muerte inducida por la enfermedad en los humanos ni en los mosquitos.
Sólo se infectan los susceptibles, teniendo en cuenta que los recuperados pasan a ser susceptibles.
El modelo está descrito por el siguiente sistema de ecuaciones diferenciales:
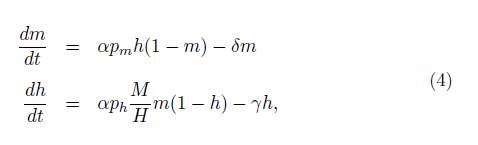
con condiciones iniciales m(0) = m 0, h(0) = h 0, donde (m 0 , h 0) = (0, 0) y las variables son:
m: proporción de mosquitos infectados
h: proporción de humanos infectados
y los parámetros son:
M/H: número de mosquitos hembra por persona
α: número de picadas sobre los humanos de un mosquito por día
p h : probabilidad de infección de un humano susceptible al ser picado por un mosquito infectado
p m : probabilidad de infección de un mosquito susceptible al picar un humano infectado
γ: tasa a la cual los humanos se recuperan de la infección
δ: tasa per capita de muerte de los mosquitos
En resumen, en el Modelo [4], suponemos que el crecimiento de los mosquitos infectados está dado por la proporción (αp m ) de contactos entre humanos infectados (h) y mosquitos sanos (1 m) y su decaimiento está dado por las muertes naturales de mosquitos (δm). Similarmente, el crecimiento de los humanos infectados está dado por la proporción (αp h M ) de contactos entre mosquitos infectados (m) y humanos sanos (1 h) y su decaimiento está dado por la recuperación de los humanos infectados (γh).
5 Estimación de los parámetros epidemiológicos
Para la estimación de los parámetros β = (α, M , p h , p m , γ, δ) T del modelo matemático descrito por el sistema de ecuaciones diferenciales [4], usamos la solución de mínimos cuadrados que se obtiene al ajustar los humanos infectados dados por el modelo y los datos recolectados por el Programa de Vigilancia Epidemiológica del Dengue de la Secretaría Municipal de Salud de Cali, durante el brote epidémico ocurrido en el 2010.
Más exactamente, dado y = (yi) , donde y i es el dato correspondiente al tiempo t i que se desea ajustar, los parámetros los estimaremos como el minimizador de la función

siendo β el vector de parámetros y (m(β), h(β)) T la solución del sistema [4] para los tiempos t i , i = 1, 2,…, N, cuando los parámetros toman los valores dados por el vector β.
Matemáticamente, el problema de mínimos cuadrados (no lineal) que deseamos resolver lo podemos plantear de la siguiente forma.
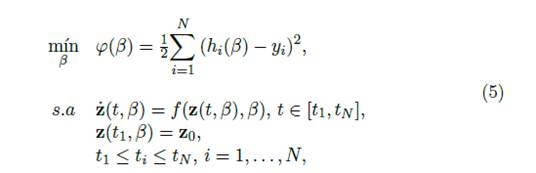
donde z = (m, h) T , β ϵ R6 y f: R2 x R6 →R2 está dada por la expresión del lado derecho que define la derivada de cada una de las variables de estado, humanos infectados y mosquitos infectados, que describen el modelo de Ross-Macdonald [4]. Esto es:

donde el vector de parámetros está representado por β = (α, p m , p h , M/H , δ, γ) T .
En la Figura 2, aparecen los casos nuevos de dengue reportados diariamente por el Programa de Vigilancia Epidemiológica del Dengue en Cali. En ella, se observan varios brotes epidémicos, destacándose por su mayor pico el de 2010. Por esta razón, para la estimación de los parámetros del modelo Ross-Macdonald, escogimos los datos de este periodo, el cual corresponde a los 98 dias comprendidos entre el día 29 y el día 126 de 2010.
Al tratar de ajustar la salida del modelo [4] a los datos mencionados, usando el criterio de mínimos cuadrados descrito previamente, encontramos que el estimativo de γ, la tasa de recuperación de los humanos infectados, oscila entre 0.083 y 0.100, lo cual corresponde a un periodo de infección en los humanos entre 10 y 12 días, pero con unos niveles de la función objetivo no muy pequeños. Por esta razón, y teniendo en cuenta los datos presentados en [15] y [16], decidimos asumir este parámetro como conocido e igual a 0.100 (10 días de enfermedad). Bajo este supuesto, convertimos

Figura 2 Incidencia diaria de dengue en Cali, 2001-2011 (Secretaría de Salud Municipal) los datos de incidencia diaria reportados por el Programa de Vigilancia Epidemiológica del Dengue en Cali en datos de prevalencia diaria para el periodo escogido.
Tabla 4 Valores iniciales (β0) y rango de los parámetros.
| Parámetro (β) | Valor inicial (β0) | Rango |
| M/H | 1 | (1, 10) |
| α | 1 | (0, 5) |
| p m | 0.5 | (0, 1) |
| p h | 0.5 | (0, 1) |
| δ | 0.035 | (1/30, 1/15) |
Para minimizar la función ', usamos la función lsqcurvefit del Toolbox de Optimización de MatLab de MathWorks Inc., tomando como punto inicial, β0, y usando los intervalos de búsqueda dados en la Tabla 4, los cuales asumimos como razonables según [17],[18] y [19].
Como para evaluar la función φ, necesitamos los valores de h(β) para los tiempos en el periodo escogido, usamos la función ode45 de MatLab para resolver el sistema de ecuaciones diferenciales [4], con condición inicial z(0, β) = z 0 = (y 1 , y 1) T (Proporción de mosquitos igual a la proporción de humanos infectados en el punto inicial del período escogido).
Tabla 5 Datos y curva de ajuste (x 10-3) y parámetros estimados γ=1/10
| Parámetros (β) | Valores Estimados (βˆ) |
| M/H | 1.4727 |
| α | 0.3352 |
| pm | 0.0834 |
| ph | 0.2499 |
| δ | 0.0333 |
Residuos = φ (βˆ) , 0.0652x10 −3
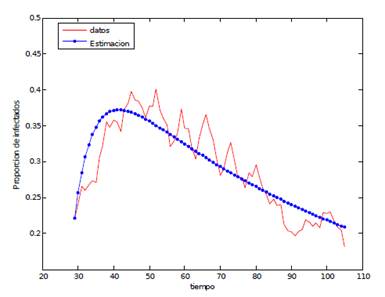
En la Tabla 5, se observa el ajuste obtenido y la tabla que registra los valores estimados de los parámetros. Según estos resultados, podemos decir que, en Cali, durante periodos epidémicos, hay 3 mosquitos por cada dos personas (M/H); que la vida media de los mosquitos es de 30 días (1/δ); que cada mosquito pica a una de cada tres personas (α), y que solo el 8 % de estas picadas infecta a los mosquitos susceptibles (pm), mientras que el 24 % de ellas infecta a las personas sanas (ph).
Con el fin de verificar la sensibilidad de los valores estimados (solución del Problema 5) respecto al punto inicial de búsqueda (β0), encontramos el minimizador de este problema variando, en cada caso, el punto inicial en uno de los parámetros con los valores dados en la Tabla 6, y encontramos las estadísticas de los valores estimados de los parámetros que aparecen en la Tabla 7.
Tabla 6 Variación de los parámetros en β 0
| Parámetro ((β0)) | Valores |
| M/H | 1, 3, 5, 7, 9 |
| α | 1, 2, 3, 4, 5 |
| p m | 0.1, 0.2, 0.4, 0.5, 0.6, 0.8 |
| p h | 0.1, 0.3, 0.5, 0.7, 0.9 |
| δ | 0.04, 0.05, 0.06, 0.07 |
Observamos que la variación no es mucha (coeficiente de variación entre 16.9 y 21.6 %) para α, p h , p m y M/H y prácticamente nula para δ y los residuos. Es de anotar que, en general, la mayor variación en los parámetros estimados fue debida a la variación en el valor inicial de α, número de picadas sobre los humanos de un mosquito por día, mientras que la menor variación en la estimación la produjo la variación en δ, la tasa de mortalidad del mosquito.
Tabla 7 Estadísticas de los parámetros estimados al variar β 0
| Estadístico | α | p h | p m | δ | M/H | Residuos |
| Promedio | 0.3606 | 0.2268 | 0.0805 | 0.0333 | 1.5969 | 0.0652 |
| Desv. estándar | 0.0777 | 0.0482 | 0.0152 | 6.7E-10 | 0.2712 | 9.0E-10 |
| Coef. variación | 0.2154 | 0.2127 | 0.1894 | 2.0E-08 | 0.1698 | 1.3E-08 |
Es de anotar que la variación en el valor de la función objetivo en el óptimo (Residuos), al variar β 0, es prácticamente nula y que la diferencia entre los valores estimados [Tabla 5] y los promedios obtenidos al variar β 0 [Tabla 7] están a menos de media desviación estándar de los estimativos obtenidos al hacer dichas variaciones; en efecto, dichas diferencias están entre 0.19 y 0.48 desviaciones estándar.
De la misma forma, al analizar la sensibilidad de los estimativos al valor de γ asumido, encontramos las estadísticas que aparecen en la Tabla 8
Tabla 8 Estadísticas de los parámetros estimados al variar γ
| Estadístico | α | ph | pm | δ | M/H | Residuos |
| Promedio | 0.3423 | 0.2556 | 0.0833 | 0.0333 | 1.4170 | 0. 0860 |
| Desv. estándar | 0.0394 | 0.0171 | 0.0083 | 1,4E-11 | 0.0938 | 0.0723 |
| Coef. variación | 0.1152 | 0.0671 | 0.1006 | 4.4E-10 | 0.0662 | 0.8402 |
Observemos que la variación en los valores estimados debida a la variación de γ, es baja (coeficiente de variación inferior al 11.6 %), lo cual valida los valores estimados bajo el supuesto hecho sobre el periodo de recuperación de 10 dias (γ = 1 ).
Es de anotar que el estimativo para δ, la tasa de mortalidad del mosquito adulto, coincide con los estimativos encontrados para este mismo parámetro (δa) cuando se usa la temperatura media de Cali en el Modelo Depredador-Presa [Tabla 3, el cual es 0.0333, que corresponde a un periodo de vida del mosquito de 30 dias.
6 Conclusiones
En este trabajo, además de presentar dos modelos sencillos para el estudio y análisis de la dinámica de la infección del dengue y la dinámica poblacional de su principal vector de transmisión en presencia de un depredador de sus estadios acuáticos, hemos adaptado ambos modelo a la ciudad de Cali, estimando sus parámetros de tal forma que uno de ellos reprodujera lo mejor posible uno de los brotes de los últimos años y que el otro tuviera en cuenta las condiciones de temperatura de la ciudad de Cali.
Una vez estimados los parámetros de ambos modelos, los podemos usar como base para la toma de decisiones y elaborar políticas de control del mosquito que transmite el dengue en la ciudad de Cali, como en efecto hacemos en [11] y [12], donde presentamos un análisis más detallado y la correspondiente interpretación de los valores de los parámetros aquí estimados, junto con el análisis de sensibilidad de sus características respecto de los parámetros. Otra ventaja de tener estos modelos adaptados a la ciudad de Cali es la posibilidad de usar el enfoque de la teoría de control óptimo con el mismo propósito de ayudar en la toma de decisiones y elaboración de políticas, lo cual constituye nuestro proximo trabajo.