











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkEste artículo presenta los resultados de una investigación ictioarqueológica, la cual consistió en el análisis de una muestra de restos óseos de peces recuperada en el sitio de La Galepia, ubicado en la región de la Depresión Momposina. El objetivo de este estudio, a largo plazo, es evaluar la importancia que los peces locales de agua dulce tuvieron en los patrones de subsistencia y alimentación de la población de La Galepia durante el periodo Formativo Tardío, el cual corresponde al lapso entre los siglos XIII y XVI d. C., según la cronología regional propuesta por Langebaek y Dever (2000). Este artículo resume los resultados de la identificación anatómica y taxonómica de la muestra ictioarqueológica de La Galepia, cuyo objetivo fue determinar las especies de peces presentes en el sitio. Asimismo, se propone una interpretación de este registro material, basada en una analogía con fuentes históricas de la región. De esta forma se busca consolidar una primera mirada a las interacciones entre peces y humanos en esta ocupación ribereña del Bajo Magdalena.
La Galepia se encuentra en zona rural del corregimiento de Santa Teresita-Tierra Firme, distrito de Mompós (Departamento de Bolívar, Caribe colombiano, véase figura 1). Las investigaciones en este predio iniciaron en el 2018, por medio de una escuela de campo organizada por el Departamento de Antropología de la Universidad de los Andes (Bogotá, Colombia). Dicha actividad, a su vez, formó parte de un proyecto arqueológico presentado por los profesores Luis Gonzalo Jaramillo y Elizabeth Ramos al Instituto Colombiano de Antropología e Historia (Jaramillo y Ramos 2021, 2018).
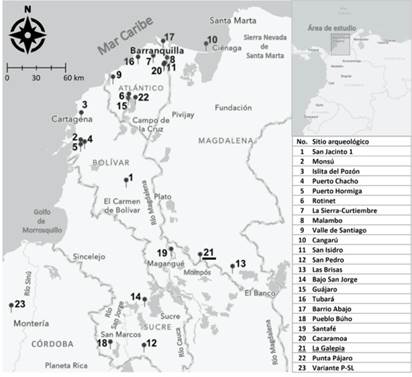
Figura 1. Ubicación del sitio de La Galepia (n.° 21) y demás sitios arqueológicos mencionados en el artículo
En el marco de ese proyecto se excavó el Corte 1 de La Galepia. En total, se demarcaron ocho niveles estratigráficos artificiales, cada uno con una profundidad de 10 cm, los cuales se ubican dentro de dos estratos naturales (Jaramillo y Ramos 2021). El material cultural, compuesto por cerámica, restos óseos, líticos, carbón y semillas, se distribuye entre los niveles 1 y 7 de esta secuencia, con una mayor concentración entre los niveles 3 y 6 (Jaramillo y Ramos 2021).
De acuerdo con las fechas de radiocarbono obtenidas en el Corte 1 de este sitio, se sabe que La Galepia fue ocupada, al menos, desde el año 1400 d. C. (siglo XV). Este depósito es producto de una acumulación de desechos (tipo basurero) asociada a una sola ocupación humana, la cual tuvo una duración relativamente corta: aproximadamente, entre el 1480 ± 30 y 1550 ± 30 d. C. (470 ± 30 a 400 ± 30 años a. p. según Jaramillo y Ramos 2021). Entre los materiales recuperados en este predio se encuentra una colección zooarqueológica compuesta por 52 931 restos óseos de fauna. Con base en investigaciones previas, se determinó que cerca del 96 % de esta colección correspondía a restos de peces (Jaramillo y Ramos 2021).
La ocupación de La Galepia es contemporánea con importantes cambios en el clima y poblamiento de la Depresión Momposina, los cuales tuvieron lugar durante el periodo Formativo Tardío. A partir del siglo XIII, esta región experimentó un periodo de baja precipitación que se prolongó hasta inicios del siglo XVI (Plazas et al. 1993). En este contexto, las poblaciones zenúes, que ocupaban la zona, la abandonaron paulatinamente y fueron reemplazadas por los grupos malibúes. Según datos etnohistóricos, estos nuevos pobladores provenían de las costas de la actual Venezuela y arribaron a la Depresión Momposina a través del río Magdalena (Bernal y Orjuela 1992; Oyuela-Caycedo 2008; Plazas et al. 1988; Rivet 1947).
Con base en análisis previos, se estableció que el sitio de La Galepia está asociado a una ocupación malibú, la cual muestra una alta correspondencia con otros yacimientos del Formativo Tardío en el Bajo Magdalena y el Bajo San Jorge. Dicha correspondencia se evidencia en los estilos cerámicos identificados en la colección del sitio, los cuales corresponden a la Tradición Cerámica Incisa Alisada, asociada al poblamiento malibú de la Región Caribe durante las etapas tardías del periodo prehispánico (Jaramillo y Ramos 2021; Martínez 2020; sobre la Tradición Cerámica Incisa Alisada véase Plazas et al. 1993; Reichel-Dolmatoff y Reichel-Dolmatoff 1991).
Por otra parte, la distribución de las diferentes clases zoológicas (peces, reptiles, mamíferos, etc.) dentro del conjunto de fauna recuperado en este sitio también muestra una correspondencia con otras ocupaciones ribereñas y lacustres de la región (Jaramillo y Ramos 2021). Por consiguiente, el análisis del material ictioarqueológico de La Galepia puede aportar información relevante sobre los usos que los humanos le dieron a la fauna y, en consecuencia, sobre las interacciones que esta población mantuvo con el entorno, en medio de los cambios ambientales y culturales que ocurrieron en la región a partir del siglo XIII. En este sentido, la presente investigación pretende aportar a las discusiones arqueológicas y antropológicas sobre el manejo del entorno en la Depresión Momposina y el Bajo Magdalena, consolidando una mirada más precisa sobre cómo los habitantes prehispánicos de La Galepia se relacionaron con el ambiente y, en particular, con la ictiofauna de los cuerpos de agua aledaños a su asentamiento.
Este artículo se divide en cinco secciones. La primera revisa las condiciones ambientales y el poblamiento humano de la Depresión Momposina. La segunda describe los métodos empleados en este estudio y la tercera presenta los resultados del análisis del material ictioarqueológico. Posteriormente, los resultados son discutidos en la cuarta sección con base en dos líneas de evidencia: el registro zooarqueológico y las fuentes históricas disponibles para la región de estudio. Por último, la quinta sección retoma los principales hallazgos de este estudio y propone algunos caminos para las investigaciones futuras.
La Depresión Momposina: entorno y poblamiento
La Depresión Momposina está conformada por una amplia llanura inundable en la que confluyen cuatro sistemas fluviales: los ríos Magdalena, Ariguaní, Cauca y San Jorge (Flórez 2018; Herrera et al. 2001). En el paisaje local predominan las planicies, múltiples colinas (de alturas menores a 100 m s. n. m.) e innumerables caños menores, humedales y ciénagas (Flórez 2018). Esta región experimenta una marcada estacionalidad climática que se refleja anualmente en dos temporadas lluviosas y dos secas, las cuales generan cambios en el nivel de los cuerpos de agua (Reichel-Dolmatoff y Reichel-Dolmatoff 1991). Los dos periodos de mayor precipitación son mayo-julio y octubre-noviembre, por lo que la Depresión se inunda entre siete y ocho meses cada año (Flórez 2018; Rojas y Montejo 2015).
En esta región se ha descrito una amplia diversidad biológica, representada por cerca de 800 variedades de plantas nativas y más de 400 especies de vertebrados (Franco 2015). La ictiofauna local está representada por más de 50 especies de peces de ciénaga y 60 de río que han sido clasificadas en dos conjuntos (Franco 2015). El primero está compuesto por especies migratorias que se desplazan río arriba al inicio de la temporada seca (entre diciembre y enero) para reproducirse en la cuenca alta y posteriormente (entre marzo y junio) retornar río abajo con las lluvias (Peña 2001). Por su parte, el segundo conjunto está representado por especies residentes, cuyas adaptaciones les permiten vivir en la cuenca baja todo el año, incluyendo los periodos secos, cuando suelen quedar aisladas en las ciénagas locales (Peña 2001).
Múltiples investigaciones han resaltado una estrecha relación entre los cuerpos de agua, la fauna y las poblaciones humanas de la Depresión Momposina y el Bajo Magdalena. Por ejemplo, se ha propuesto que el poblamiento de dicha área se dio en torno a los ambientes acuáticos. Distintos grupos tendieron a asentarse en los márgenes de ríos, caños y ciénagas, con el fin de favorecer el aprovechamiento de los recursos hídricos, entre ellos, la fauna silvestre (Carvajal 2019; Castro, Beltrán y Rivera 2018; Márquez 2017; Ramos y Niño 2019; Reichel-Dolmatoff 1997; Trillos 2001).
No obstante, el modelo tradicional que se planteó para entender el cambio social en esta región se basa en la idea de que el desarrollo de la agricultura (durante el periodo Formativo) llevó a que las sociedades prehispánicas del Caribe colombiano basaran su subsistencia en el cultivo intensivo de plantas. Por lo tanto, empleaban la caza y pesca como estrategias complementarias (o secundarias) para la obtención de recursos (Reichel-Dolmatoff 1997). De acuerdo con este esquema, la agricultura constituyó un motor de cambio que hizo posible la consolidación del sedentarismo y, con ello, el surgimiento de sociedades complejas, cuya subsistencia se basó, ya no en el forrajeo, sino en la producción de alimentos (Reichel-Dolmatoff 1997).
Ahora bien, los estudios recientes invitan a replantear esta noción. Las investigaciones han mostrado que, si bien hubo similitudes en los patrones de subsistencia a través de la región, estos presentan una notoria diversidad local. Por lo tanto, no pueden ser entendidos bajo modelos monolíticos ni homogéneos (Ramos 2019; Ramos y Archila 2008). Esto se debe a que los grupos humanos habitaron diferentes ambientes en la región, por lo cual desarrollaron estrategias particulares para subsistir en cada entorno (Carvajal 2019; Langebaek 1996; Langebaek y Dever 2000; Ramos 2019; Ramos y Archila 2008; Ramos y Niño 2019).
En particular, los análisis zooarqueológicos evidencian que la pesca fue una estrategia fundamental para la adaptación de diferentes poblaciones humanas al entorno de la llanura aluvial. La frecuencia de restos óseos de peces suele ser mayor en las ocupaciones ribereñas y lacustres, lo cual indica que las sociedades de la región aprovecharon de forma eficiente la fauna local y que la pesca no fue una actividad secundaria, sino, posiblemente, una práctica primordial para su supervivencia, incluso durante las etapas tardías del periodo Formativo (Carvajal 2019; Castro, Beltrán y Rivera 2018; Flórez 2018; Jaramillo y Ramos 2021; Márquez 2017; Ramos y Niño 2019).
Sin embargo, para corroborar estas tendencias es preciso realizar investigaciones en diversas escalas de análisis, con miras a forjar una comprensión más precisa sobre las interacciones entre sociedad y entorno en esta región. Por ejemplo, es fundamental explorar las variaciones del registro zooarqueológico a nivel de unidades domésticas (véase Álvarez 2022), así como en distintos tipos de sitios (cocheros, basureros, plataformas agrícolas, sitios de vivienda, etc.) y, de forma más amplia, a nivel regional (véase Carvajal 2019; Ramos 2019). De esta forma será posible construir un panorama amplio sobre los lazos entre humanos y fauna en el área de estudio. Este trabajo propone una perspectiva sincrónica que se centra en el rol de la ictiofauna local en la subsistencia de los ocupantes de La Galepia durante el Formativo Tardío. A futuro, se busca que estos resultados tengan un alcance más amplio, siendo cotejados con los análisis de otros sitios de la región, con el fin de ampliar nuestro entendimiento sobre el papel de la pesca en las estrategias adaptativas de las poblaciones prehispánicas del Caribe colombiano.
Sobre los métodos
La presente investigación se basó en dos etapas metodológicas. La primera consistió en un análisis zooarqueológico, cuyo objetivo fue realizar la identificación anatómica y taxonómica de la muestra de restos de peces de La Galepia -siguiendo la propuesta de Reitz y Wing (2008). Para tal fin, se realizó un trabajo en el Laboratorio de Zooarqueología de la Universidad de los Andes, el cual consistió en comparar la muestra con la colección de referencia disponible en dicha entidad. Este proceso permitió identificar rasgos diagnósticos en los restos óseos, con el fin de asignarlos a unidades anatómicas (elementos óseos) y/o taxones, según los lineamientos de Bonomo, Skarbun y Bastourre (2019). Simultáneamente, se elaboró una base de datos para registrar la distribución de los elementos óseos y taxones en la secuencia de ocupación de La Galepia. Con base en este registro, se calculó la frecuencia de las categorías anatómicas y taxonómicas, a partir del número de especímenes identificados por taxón (o NISP, por sus siglas en inglés, véase Bonomo, Skarbun y Bastourre 2019; Reitz y Wing 2008).
Este estudio pretende aproximarse a las estrategias de subsistencia que fueron usadas por los habitantes de La Galepia. En este contexto, el término subsistencia es entendido como la búsqueda y obtención de los recursos necesarios para el sostenimiento de una comunidad humana, entre los cuales destacan los alimentos (Bonomo, Skarbun y Bastourre 2019; Marciniak 2005). En este sentido, por medio de la identificación anatómica y taxonómica, se estableció qué tipo de peces fueron capturados y llevados al sitio, lo cual, a su vez, permitió determinar si se trató de especies locales, propias de los ambientes acuáticos aledaños al asentamiento. De esta forma fue posible consolidar una primera mirada a la subsistencia en La Galepia, explorando las especies que fueron obtenidas y los ecosistemas que fueron aprovechados por sus ocupantes durante el Formativo Tardío.
A la fecha no se han realizado estudios tafonómicos que permitan identificar y analizar alteraciones térmicas, marcas de corte u otras huellas de manipulación humana en la muestra estudiada. No obstante, diversos autores han destacado la relevancia de los análisis tafonómicos para la interpretación rigurosa de los conjuntos zooarqueológicos (véase Bonomo, Skarburn y Bastourre 2019; Lyman 1994; Reitz y Wing 2008; Wheeler y Jones 1989). Por consiguiente, se prevé que este componente sea incorporado en análisis futuros, con el fin de estudiar la formación del sitio y los usos que los humanos les dieron a los peces en ese contexto.
Por otra parte, para la segunda etapa de esta investigación se realizó una revisión bibliográfica, la cual tuvo dos objetivos:
Sintetizar los datos arqueológicos e históricos sobre el uso de peces por parte de las poblaciones humanas de la Depresión Momposina y el Bajo Magdalena.
Comparar y contrastar los datos regionales con la información obtenida de la muestra ictioarqueológica de La Galepia.
Con este fin, se consultó un total de treinta documentos arqueológicos y once fuentes históricas, entre ellas, crónicas, descripciones y relaciones correspondientes al área de estudio (Gutiérrez-Ferro 2022). En el caso de las fuentes históricas, se privilegiaron los documentos que han sido citados en diferentes investigaciones sobre las tradiciones alimentarias y culinarias del Caribe colombiano o, en general, sobre los grupos malibúes (Bernal y Orjuela 1992; Carvajal 2019; Escalante 2002, 1955; Illera 2012; Langebaek 1996; Rivet 1947; Saldarriaga 2009, 2006).
En síntesis, durante la segunda etapa de la investigación, los datos de La Galepia fueron contrastados con el registro arqueológico e histórico de la Región Caribe. De esta forma se evaluó si las evidencias obtenidas en el sitio estudiado son consecuentes con los patrones identificados a nivel regional. Lo anterior permitió interpretar los posibles usos que los humanos les dieron a los peces en el contexto de esta ocupación prehispánica.
Los peces en La Galepia
Durante la primera etapa de la investigación se determinó la distribución de los restos óseos de peces sobre el total del material zooarqueológico presente en la secuencia estratigráfica de La Galepia. Como se mencionó anteriormente, en el corte 1 de este sitio se excavaron ocho niveles artificiales, siete de los cuales contenían material cultural. Los ocho niveles se excavaron sobre dos estratos naturales. Por consiguiente, se determinó que cada nivel fuera excavado en capas de aproximadamente 3 cm, hasta completar una profundidad de 10 cm, con el fin de identificar y registrar los cambios de estrato y las concentraciones de materiales arqueológicos con precisión (Jaramillo y Ramos 2021). La tabla 1 presenta la distribución de los restos de peces sobre el total de restos óseos de fauna en esta secuencia de ocupación (para un perfil del corte 1 del sitio, véase Jaramillo y Ramos 2021).
Tabla 1. Distribución de restos óseos de peces en el corte 1 de La Galepia
| Unidad estratigráfica | Total restos fauna | Restos peces | % Peces | |
|---|---|---|---|---|
| Estrato | Nivel | |||
| 1 | N1 | 91 | 58 | 63,74 |
| 1 | N2 | 355 | 197 | 55,49 |
| 1 | N3 | 1803 | 1386 | 76,87 |
| 1 | N4 | 14989 | 12027 | 80,24 |
| 1-2 | N5 | 30083 | 29536 | 98,18 |
| 2-1 | N6 | 1063 | 965 | 90,78 |
| 2 | N7 | 4547 | 4449 | 97,84 |
| Total Corte 1 | 52931 | 48618 | 91,85 | |
Fuente: elaboración propia, con base en Jaramillo y Ramos (2021).
En el transcurso de esta etapa, fue necesario revisar la clasificación que se había hecho a la muestra previamente y realizar algunos ajustes a la identificación taxonómica1. Como resultado de este proceso, fue posible identificar el 58,9 % de la muestra a nivel anatómico y el 100 % a nivel taxonómico (tabla 2). El nivel anatómico está conformado por categorías que se refieren a los elementos óseos que componen la anatomía de los peces (ej. espinas, vértebras, huesos del cráneo, etc.). Por su parte, el nivel taxonómico está conformado por la jerarquía de categorías que se usan sistemáticamente para clasificar a los seres vivos, entre ellas: clase, orden, familia, género y especie (para una breve descripción de la taxonomía de los peces, con énfasis en los grupos de mayor interés para la arqueología, véase Wheeler y Jones 1989).
Tabla 2. Porcentaje de restos identificados en la muestra ictioarqueológica de La Galepia
| Ítem | Nivel anatómico | Nivel taxonómico | ||
|---|---|---|---|---|
| NR* | % | NR* | % | |
| Identificado | 28622 | 58,87 | 48618 | 100 |
| No identificado | 19996 | 41,13 | 0 | 0 |
| Total | 48618 | 100 | 48618 | 100 |
Nota: *NR: número de restos.
Fuente: elaboración propia, con base en Gutiérrez-Ferro (2022).
Se debe tener presente que el nivel de resolución taxonómica de la muestra es variable. El 88,7 % del total de restos de peces solo pudo ser identificado a nivel de clase (Osteichthyes: peces óseos) pues se trata de restos altamente fragmentados, cuyos rasgos diagnósticos no pudieron ser distinguidos con mayor especificidad. Dentro de esta parte de la muestra también se encuentra un gran número de vértebras y espinas, las cuales, pese a estar en su mayoría completas y ser fácilmente identificadas a nivel anatómico, presentan pocas características que permitan asignarlas a categorías taxonómicas más precisas. El resto de la muestra se distribuye de la siguiente manera: 9,6 % fue identificado a nivel de especie, 1,29 % a nivel de orden y 0,48 % a nivel de género (tabla 3).
Tabla 3. Nivel de resolución taxonómica de la muestra analizada
| Nivel taxonómico | NISP* | % |
|---|---|---|
| Clase | 43111 | 88,67 |
| Orden | 627 | 1,29 |
| Género | 234 | 0,48 |
| Especie | 4646 | 9,56 |
| Total | 48618 | 100,00 |
Nota: *NISP: número de especímenes identificados por taxón.
Fuente: elaboración propia, con base en Gutiérrez-Ferro (2022).
Con base en lo anterior, se determinó que la muestra ictioarqueológica de La Galepia está representada por distintas especies de peces óseos, las cuales pertenecen a los siguientes ordenes: Siluriformes, Perciformes, Characiformes y Synbranchiformes. Todas las especies identificadas son locales y habitan los cuerpos de agua dulce de la Depresión Momposina y la cuenca baja del río Magdalena. Cinco de estas especies tienen hábitos migratorios relacionados con sus ciclos reproductivos. No obstante, también se identificaron tres especies residentes, las cuales cuentan con adaptaciones que les permiten permanecer en la región durante las estaciones secas (tabla 4).
Tabla 4. Ictiofauna identificada en el corte 1 de La Galepia
Fuente: elaboración propia, la información sobre el hábitat y comportamiento migratorio de las especies fue tomada de Ramos y Niño (2019) y Peña (2001).
De acuerdo con el número de especímenes identificados por taxón (NISP) la especie más frecuente en la muestra es Hoplosternum magdalenae (chipe) (tabla 5). Esto puede deberse a que cada ejemplar de esta especie posee un gran número de placas dérmicas que recubren su organismo, lo cual puede producir una sobrerrepresentación del taxón en las muestras arqueológicas -tendencia reportada por Ramos y Niño (2019). En el contexto de esta investigación, la mayor parte del material identificado para esta especie corresponde a dichas placas dérmicas (figura 2).
Tabla 5. Abundancia taxonómica de la muestra ictioarqueológica de La Galepia
| Taxón | NISP* | % |
|---|---|---|
| H. magdalenae | 3410 | 61,92 |
| Siluriformes | 546 | 9,91 |
| Pr. magdalenae | 372 | 6,76 |
| C. kraussii | 370 | 6,72 |
| P. fasciatum | 242 | 4,39 |
| Pimelodus sp. | 234 | 4,25 |
| T. insignis | 122 | 2,22 |
| Pl. magdalenae | 75 | 1,36 |
| Characiformes | 56 | 1,02 |
| S. marmoratus | 50 | 0,91 |
| Perciformes | 25 | 0,45 |
| P. blochii | 5 | 0,09 |
| Total | 5507 | 100,00 |
Nota: *NISP: número de especímenes identificados por taxón.
Fuente: elaboración propia, con base en Gutiérrez-Ferro (2022).
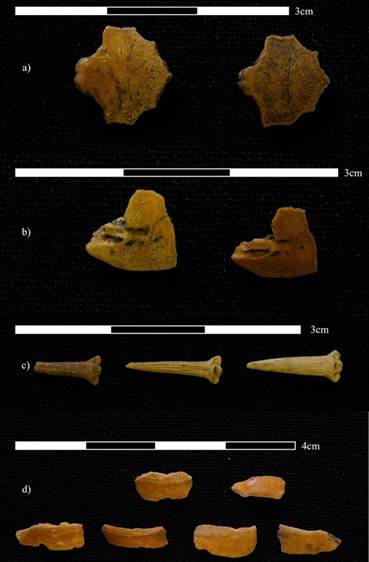
Fuente: fotografía cortesía de Mauricio Salinas Rozo, Laboratorio de Antropología Visual, Universidad de los Andes, Colombia, 2023.
Figura 2. Material arqueológico identificado para la especie Hoplosternum magdalenae (chipe)
Las siguientes especies con mayor frecuencia en la muestra son Prochilodus magdalenae (bocachico, véase figura 3) y Caquetaia kraussii (mojarra amarilla, véase figura 4), seguidas por Pseudoplatystoma fasciatum (bagre rayado) y el género Pimelodus (barbudos). También se identificaron, en porcentajes menores, las especies Trachelyopterus insignis (doncella), Plagioscion magdalenae (curvinata), Synbranchus marmoratus (anguila) y Pimelodus blochii (nicuro).

Nota: a) opérculo, b) frontal, c) espinas dorsales. Fuente: fotografía cortesía de Mauricio Salinas Rozo, Laboratorio de Antropología Visual, Universidad de los Andes, Colombia, 2023.
Figura 3. Material arqueológico identificado para la especie Prochilodus magdalenae (bocachico)

Fuente: fotografía cortesía de Mauricio Salinas Rozo, Laboratorio de Antropología Visual, Universidad de los Andes, Colombia, 2023.
Figura 4. Material arqueológico identificado para la especie Caquetaia kraussii (mojarra amarilla)
Por otro lado, el 9,9 % del material identificado a un nivel de resolución mayor al de clase está compuesto por restos correspondientes al orden Siluriformes; 1,02 % a los Characiformes; y 0,45 % a los Perciformes (tabla 5). Lo anterior se debe a que existen grandes similitudes en la estructura ósea de las especies que conforman estos órdenes, lo cual impidió identificar la totalidad de los restos a nivel de especie. A futuro, se busca que la consulta de una colección de referencia especializada permita refinar el nivel de resolución taxonómica de esta porción de la muestra.
Por otra parte, con base en la identificación anatómica se determinó que los elementos óseos más frecuentes son las vértebras y espinas (tabla 6). Esto puede deberse a que cada organismo posee un gran número de estos huesos, por lo que suele darse una sobrerrepresentación de estas unidades anatómicas en las muestras ictioarqueológicas. Los siguientes elementos con mayor representación son los huesos del cráneo y las placas dérmicas (estas últimas, correspondientes a la especie H. magdalenae). El resto del material identificado está conformado por diferentes huesos de la cara (articular, cleitron, dentario, maxilar, entre otros) cuyos porcentajes son menores (tabla 6).
Tabla 6. Abundancia anatómica de la muestra ictioarqueológica de La Galepia
| Elemento | Total muestra | Total identificado | ||
|---|---|---|---|---|
| NR* | Frecuencia absoluta (%) | NR* | Frecuencia relativa (%) | |
| Articular | 62 | 0,13 | 62 | 0,22 |
| Cleitron | 272 | 0,56 | 272 | 0,95 |
| Cráneo | 5560 | 11,44 | 5560 | 19,43 |
| Cuadrado | 55 | 0,11 | 55 | 0,19 |
| Dentario | 151 | 0,31 | 151 | 0,53 |
| Dientes premaxilar | 103 | 0,21 | 103 | 0,36 |
| Espinas | 8271 | 17,01 | 8271 | 28,90 |
| Hyomandibular | 5 | 0,01 | 5 | 0,02 |
| Maxilar | 53 | 0,11 | 53 | 0,19 |
| Placas dérmicas | 3368 | 6,93 | 3368 | 11,77 |
| Premaxilar | 45 | 0,09 | 45 | 0,16 |
| Vértebras | 10677 | 21,96 | 10677 | 37,30 |
| No identificado | 19996 | 41,13 | n.a. | n. a. |
| Total | 48618 | 100 | 28622 | 100 |
Nota: *NR: Número de restos.
Fuente: elaboración propia, con base en Gutiérrez-Ferro (2022).
Por último, vale la pena mencionar que una porción pequeña de la muestra (compuesta por cerca de 200 restos, relativamente completos y bien preservados) fue identificada a nivel anatómico, asociando dichos restos con los siguientes elementos: cuadrado, dentario y premaxilar. No obstante, a nivel taxonómico, no mostraron correspondencia con ninguno de los especímenes de la colección de referencia consultada en la Universidad de los Andes. Por consiguiente, solo pudieron ser identificados a nivel de clase.
La Galepia en el contexto regional
Una vez completado el análisis zooarqueológico, el siguiente paso consistió en cotejar los datos de La Galepia con los patrones de uso de los peces que se han identificado a nivel regional. A continuación, se presenta una discusión de los resultados basada en un diálogo con el registro arqueológico e histórico del área de estudio.
El registro arqueológico
Los restos óseos de peces han sido reportados en veintitrés sitios arqueológicos de la Región Caribe de Colombia (véase tabla 7; Gutiérrez-Ferro 2022; Ramos s. f.; Ramos y Gutiérrez-Ferro s. f.). Estas ocupaciones cubren un margen de tiempo muy amplio que abarca desde el periodo Arcaico, entre 6000 y 5000 años a. p. en el caso de San Jacinto 1 (Stahl y Oyuela-Caycedo 2007) hasta el siglo XVI d. C. (± 450 años a. p.) al final de las ocupaciones de sitios como Barrio Abajo (Rivera-Sandoval 2018) y Tubará (Ramos y Archila 2008). No obstante, el nivel de resolución taxonómica varía considerablemente entre estos sitios y solo en doce casos se han realizado cuantificaciones que permitan conocer la distribución de los taxones dentro de los conjuntos de fauna (véase tabla 7; Ramos s. f.; Ramos y Gutiérrez-Ferro s. f.).
Tabla 7. Distribución de tres clases zoológicas (peces, reptiles y mamíferos) en sitios arqueológicos del Caribe colombiano
| Sitio | Cronología | Ubicación (véase figura 1) | % Taxón | Referencias | |||
|---|---|---|---|---|---|---|---|
| Peces | Reptiles | Mamíferos | n. d. | ||||
| San Jacinto 1a | 5940 ± 60 a. p. - 5190 ± 40 a .p. | Sabanas de Bolívar | 4,37 | 31,45 | 16,64 | 47,18 | Stahl y Oyuela-Caycedo (2007) |
| Monsúb | 5300 ± 80 a. p. - 2800 ± 80 a. p. | Canal del Dique | 61,18 | 4,27 | 1,84 | 31,78 | Diaz-Chauvigne (2016); Carvajal (2019) |
| La Sierra-Curtiembre | 2000 a. C. - 1540 ± 70 d. C. (varias ocupaciones) | Llanuras del Bajo Magdalena | 10,0 | 19,0 | 24,0 | 38,0 | Langebaek y Dever (2000) |
| Malambo | 3070 ± 200 a. p. - 1890 ± 2000 a. p. - 500 ± 30 a. p. | Ciénaga de Malambo, Bajo Magdalena | 18,5 | 49,3 | 22,7 | n. a. | Angulo (1991); Archila (1993); Rivera-Sandoval (2018) |
| San Pedro | 680 ± 120 d. C. | Bajo San Jorge, Depresión Momposina | 45,7 | 19,9 | 22,2 | n. a. | Flórez (2018); Rojas y Montejo (2015) |
| San Isidro | 640 d. C. - 690 d. C. | Ciénaga el Convento, Bajo Magdalena | 22,6 | 42,82 | 3,56 | 30,7 | Ramos (2019) |
| Las Brisas | Siglos VIII - XVI d. C. (cronología relativa) | Ciénaga el Piñal, Bajo Magdalena | 99,2 | 0,5 | 0,3 | n. a. | Rodríguez y Peña (2001) |
| Tubará | 850 ± 50 d. C. - 1080 ± 80 d. C. - 1570 ± 70 d. C. | Serranía de Piojó | Corte 3 | Corte 3 | Corte 3 | Corte 3 | Ramos y Archila (2008); Ramos (2019) |
| 8,65 | 36,5 | 11,31 | 41,85 | ||||
| Corte 4 | Corte 4 | Corte 4 | Corte 4 | ||||
| 44,94 | 7,83 | 11,93 | 30,82 | ||||
| Barrio Abajoc | 770 ± 30 a. p. - 650 ± 30 a. p. | Bajo Magdalena | 81,5 | 14,6 | 3,2 | n. a. | Castro et al. (2018); Márquez (2017); Rivera-Sandoval (2018) |
| Cacaramoa | 1430 d. C. - 1480 d. C. | Ciénaga el Convento, Bajo Magdalena | Total | Total | Total | Total | Ramos (2019); Ramos y Niño (2019) |
| 79,03 | 6,77 | 0,14 | 13,92 | ||||
| Pozo 24 | Pozo 24 | Pozo 24 | Pozo 24 | ||||
| 95,09 | 3,83 | 0,08 | 0,86 | ||||
| Santaféd | 600 ± 30 - 640 ± 30 a. p. | Depresión Momposina | 97,9 | 0,8 | 0,8 | n. a. | Álvarez (2022) |
| Punta de Pájaro | s. f. | Ciénaga de Guájaro, Bajo Magdalena | 61,0 | 24,0 | 15,0 | n. a. | Lozano (2014) |
Nota: a) para el caso de San Jacinto 1, se calcularon los porcentajes con base en el total de especímenes identificado para cada clase zoológica, reportado por Stahl y Oyuela-Caycedo (2007, 335); b) para el sitio de Monsú, los porcentajes corresponden a la muestra de vertebrados analizada por Diaz-Chauvigne (2016); c) en el sitio de Barrio Abajo se recuperó material correspondiente a varios periodos. Esta tabla sólo incluye los datos correspondientes al periodo Prehispánico; d) para el caso de Santafé, los porcentajes fueron calculados con base en el mínimo número de individuos (MNI) determinado por Álvarez (2022, 90).
Fuente: elaboración propia, con base en Gutiérrez-Ferro (2022); Ramos (s. f.).
Cinco de estos sitios, conocidos como Las Brisas, Barrio Abajo, Tubará, Cacaramoa y Santafé, están asociados al poblamiento malibú del periodo Formativo Tardío. En general, estas ocupaciones muestran un comportamiento similar: los peces suelen ser la clase con mayor representación, seguida por los reptiles y mamíferos (Ramos s. f.; Ramos y Gutiérrez-Ferro s. f.). Muestra de ello es que los peces son la clase predominante en cuatro de los cinco sitios malibúes, al igual que en dos de los sitios más tempranos (Monsú y San Pedro) asociados con otras ocupaciones del periodo Formativo (tabla 7).
No obstante, existen dos factores que dificultan la comparación entre estos sitios. Por un lado, en las últimas décadas se han implementado cambios en los procedimientos de excavación y recuperación de restos faunísticos, en particular en el caso de los peces, a raíz de múltiples sesgos metodológicos que inducían a errores en la interpretación. Por ejemplo, el uso de mallas más finas para el cernido de capas de suelo ha hecho posible que abundantes restos ictioarqueológicos, que antes eran ignorados, ahora sean incorporados en los análisis de conjuntos óseos (Morales-Muniz y Llorente-Rodriguez 2020; Ramos 2014). Por otro lado, diferencias en los protocolos de identificación y clasificación de los restos óseos en el laboratorio también pueden producir sesgos en los resultados (Lyman 2019). Por lo tanto, los datos presentados en la tabla 7 deben leerse con precaución, pues las variaciones entre sitios pueden deberse no solo a cambios en las estrategias de subsistencia a través del tiempo y el espacio, sino a distintas aproximaciones teórico-metodológicas en la arqueología de la Región Caribe.
Teniendo en cuenta lo anterior, se emplearon dos criterios para la interpretación del registro zooarqueológico. En primer lugar, la distribución de las clases zoológicas en los conjuntos faunísticos fue tomada como un indicio general sobre los ambientes aprovechados por los grupos humanos para la obtención del alimento, es decir, para su subsistencia. Por esta razón, en segundo lugar, se tuvo en cuenta la relación entre dicha distribución taxonómica y la ubicación de los sitios, con miras a identificar tendencias a nivel regional. Sin embargo, se entiende que estos porcentajes no constituyen la reconstrucción de una dieta, entendida como patrón de consumo de alimentos a lo largo del tiempo (en términos de Bonomo, Skarburn y Bastourre 2019) ya que la naturaleza desigual de los datos impide llegar a ese nivel de especificidad en la interpretación.
Ahora bien, pese a las limitaciones de la información arqueológica disponible, fue posible identificar ciertos patrones en el área de estudio. Tal como se esperaría, la frecuencia de restos de peces tiende a ser mayor en los sitios del litoral y aquellos que se ubican en inmediaciones de ríos, caños y ciénagas. Por otra parte, los reptiles y mamíferos suelen tener una mayor representación en los yacimientos situados en serranías, zonas de sabana o bosque seco tropical (ej. Tubará, La Sierra y Curtiembre) (Ramos s. f.; Ramos y Gutiérrez-Ferro s. f.). Estas variaciones podrían deberse a que los sitios se ubicaron en ecosistemas distintos y, por lo tanto, sus habitantes emplearon estrategias diferentes para obtener el alimento, aunque también podrían haber influido factores culturales y cronológicos (Ramos 2019).
Es factible que debido a su ubicación geográfica las poblaciones de las sabanas, serranías y bosques hayan privilegiado la obtención de fauna terrestre (en especial, mamíferos y reptiles). Por su parte, en el conjunto de sitios de las llanuras aluviales, la evidencia indica que sus habitantes privilegiaron el uso de animales acuáticos y semiacuáticos, propios de los ríos y ciénagas (Carvajal 2019; Ramos y Niño 2019). Esto se refleja, no solo en la predominancia de los peces, sino también en la presencia de reptiles semiacuáticos, como la tortuga hicotea (Trachemys callirostris), babillas (Caiman crocodilus) y caimanes (Crocodylus acutus) en las muestras zooarqueológicas recuperadas en esta área (Archila 1993; Castro, Beltrán y Rivera 2018; Flórez 2018; Márquez 2017; Ramos 2019; Ramos y Niño 2019).
Los peces que integraron la oferta alimentaria de las poblaciones prehispánicas de la Región Caribe fueron, en su mayoría, especies locales de agua dulce. Por ejemplo, el bocachico (Pr. Magdalenae) y el bagre rayado (Pseudoplatystoma sp.) son las dos especies con mayor ubicuidad en el registro arqueológico de la región. El primero ha sido identificado en doce sitios arqueológicos y el segundo en once, sobre un total de veintidós sitios que reportan restos óseos de peces (tabla 8). Otras especies, como el nicuro (P. blochii), la doncella (T. insignis) y la anguila del lodo (S. marmoratus) también presentan una alta ubicuidad, seguidas por la dorada (Brycon sp.), el moncholo (Hoplias malabaricus) y la curvinata (Plagioscion sp.). Por su parte, los loricáridos (Loricariidae), las mojarras (Caquetaia sp.) y otros taxones se encuentran menos representados a nivel regional (véase tabla 8; Ramos s. f.; Ramos y Gutiérrez-Ferro s. f.).
Tabla 8. Ubicuidad de ictiofauna en sitios arqueológicos del área de estudio (parte 1)
| Sitio | Taxón (ver abreviaturas y notas al final del cuadro) | Referencia | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pro mag | Pse sp.* | Syn mar | Pim blo | Tra ins | Pla sp. | Bry sp. | Caq sp. | Hop mal | Tri mag | Sil | Sor lim | Lep muy | Ari | Lor | Sor cus | Hop mag | Pim gro | ||
| San Jacinto 1 | X | Stahl y Oyuela-Caycedo (2007) | |||||||||||||||||
| Monsú | X | X | Carvajal (2019); Diaz-Chauvigne (2016) | ||||||||||||||||
| Puerto Chacho | X | X | X | X | Álvares y Maldonado (2009); Archila (1993) | ||||||||||||||
| Islita del Pozón | X | Carvajal (2019) | |||||||||||||||||
| Puerto Hormiga | X | X | X | Archila (1993); Carvajal (2019) | |||||||||||||||
| Rotinet | X | X | X | X | X | X | Angulo (1995; 1988); Archila (1993) | ||||||||||||
| Malambo | X | X | X | Angulo (1981); Archila (1993) | |||||||||||||||
| Bajo San Jorge | X | X | X | X | X | Plazas y Falchetti (1981); Plazas et al. (1993) | |||||||||||||
| Valle de Santiago | X | Angulo (1983); Archila (1993) | |||||||||||||||||
| San Pedro | X | Flórez (2018) | |||||||||||||||||
| Guájaro | X | X | X | X | X | X | X | X | X | X | X | Peña (2001) | |||||||
| Tubará | X | X | X | Ramos y Archila (2008); Ramos (2019) | |||||||||||||||
| Cangarú | X | Archila (1993) | |||||||||||||||||
| Pueblo Búho | X | X | X | X | X | X | X | X | Rojas y Montejo (2006) | ||||||||||
| Las Brisas | X | X | X | X | X | X | X | X | X | X | X | X | Rodríguez y Peña (2001) | ||||||
| Barrio Abajo | X | X | X | X | X | X | X | X | X | X | X | X | X | X | X | Castro et al. (2018); Márquez (2017) | |||
| Cacaramoa | X | X | X | X | X | X | X | X | X | X | X | Ramos y Niño (2019) | |||||||
| Santafé | X | X | X | X | X | X | X | Álvarez (2022) | |||||||||||
| Punta de Pájaro | X | X | X | X | Lozano (2014) | ||||||||||||||
| Variante PSL | X | X | X | X | X | X | X | Sánchez (2002) | |||||||||||
| Total registros | 12 | 11 | 9 | 8 | 8 | 7 | 6 | 5 | 5 | 5 | 5 | 4 | 4 | 4 | 4 | 3 | 2 | 2 | - |
| Abreviaturas | |||||
|---|---|---|---|---|---|
| Marcador | Taxón | Marcador | Taxón | Marcador | Taxón |
| Pro mag | Prochilodus magdalenae | Bry sp. | Brycon sp. | Lep muy | Leporinus muyscorum |
| Pse sp.* | Pseudoplatystoma sp. | Caq sp. | Caquetaia sp. | Ari | Ariidae |
| Syn mar | Synbranchus marmoratus | Hop mal | Hoplias malabaricus | Lor | Loriicariidae |
| Pim blo | Pimelodus blochii | Tri mag | Triportheus magdalenae | Sor cus | Sorubim cuspicaudus |
| Tra ins | Trachelyopterus insignis | Sil | Siluriformes | Hop mag | Hoplosternum magdalenae |
| Pla sp. | Plagioscion sp. | Sor lim | Sorubim lima | Pim gro | Pimelodus grosskopfii |
Nota: en la región de estudio se han reportado dos especies del génenro Pseudoplatystoma (P. fasciatum y P. magdalenae). Para propóstiso de síntesis, los datos referentes a estas dos especies fueron agrupados en una sola columna, la cual corresponde a Pseudolpatystoma sp. (abreviatura: Pse sp.)
Fuente: elaboración propia, con base en Gutiérrez-Ferro (2022); Ramos (s. f.).
No obstante, se debe tener presente que para la mayoría de los sitios únicamente se dispone de listados (o inventarios) de fauna identificada. Por lo tanto, ante la ausencia de cálculos de abundancia taxonómica, se desconoce la distribución real de estos taxones dentro de los conjuntos zooarqueológicos. Sin embargo, el hecho de que estos hayan sido reportados en numerosos sitios a través de la región evidencia que, dentro de la amplia diversidad de la fauna acuática local, las poblaciones humanas han privilegiado la obtención y consumo de un rango limitado de especies (Ramos s. f.).
Con base en lo anterior, es posible establecer dos patrones generales para la región de estudio, los cuales han sido señalados previamente por investigadoras como Archila (1993), Carvajal (2019) y Ramos (s. f.):
Los peces son la clase más frecuente en las ocupaciones ribereñas y lacustres, mientras que los reptiles y mamíferos tienen mayor representación en los sitios ubicados en ambientes de sabana, serranía y bosque seco.
La ictiofauna identificada en los sitios arqueológicos de la Región Caribe corresponde a ambientes diversos. No obstante, en los yacimientos aledaños a ríos, caños y ciénagas predominan las especies locales de agua dulce.
La muestra ictioarqueológica de La Galepia indica una alta correspondencia con estas tendencias regionales. La ictiofauna del sitio está compuesta por especies locales de agua dulce, las cuales se encuentran dentro del rango de taxones que predominan en las muestras zooarqueológicas del área de estudio. Por ejemplo, el taxón más frecuente de La Galepia (H. magdalenae) ha sido identificado en otros dos yacimientos de ocupación malibú: Barrio Abajo y Cacaramoa (Márquez 2017; Ramos y Niño 2019). Otras especies identificadas (Pr. magdalenae, P. fasciatum, T. insignis, S. marmoratus y P. blochii) también han sido reportadas en varios sitios del Medio y Bajo San Jorge y del Bajo Magdalena, tales como Guájaro, Tubará, Las Brisas, Barrio Abajo y Cacaramoa (véase tabla 8). Por último, algunas especies representadas en la muestra (C. kraussi y Pl. magdalenae) son menos frecuentes a nivel regional. No obstante, los géneros a los que pertenecen (Caquetaia y Plagioscion, respectivamente) comparten una distribución similar en el área de estudio (tabla 8).
Con base en esto, se puede plantear un comportamiento similar al de otras ocupaciones malibúes, como Barrio Abajo. Según Castro y sus colegas (2018, sin paginación) en este sitio “el área de captación” de los recursos faunísticos no fue muy amplia, sino que se ubicó en cercanías de los sitios de vivienda. En otras palabras, ciertos grupos malibúes aprovecharon la ubicación estratégica de sus asentamientos, en cercanías del río Magdalena y las ciénagas para obtener y consumir peces locales. Esto concuerda con el patrón regional que se ha descrito para el periodo Formativo, según el cual varias poblaciones de la llanura inundable desarrollaron un modo de vida sedentario, caracterizado por “una marcada orientación ribereña” (Reichel-Dolmatoff 1997, 64-65). Por esta razón, las estrategias de subsistencia de varias poblaciones locales tuvieron una alta dependencia de los recursos acuáticos (Márquez 2017; Plazas et al. 1993; Ramos s. f.; Reichel-Dolmatoff y Reichel-Dolmatoff 1991).
En suma, los datos ictioarqueológicos de La Galepia reflejan el “modo de vida lacustre” de los grupos malibúes, “caracterizado por el establecimiento sobre espacios acuáticos o cercanos a ellos, con una economía basada en el aprovechamiento de recursos de ciénaga”, entre ellos, los peces locales (Márquez 2017, 35). El análisis de la muestra del sitio indica que los cuerpos de agua dulce fueron fuentes primarias de recursos para los habitantes de La Galepia, tendencia similar a la descrita para la población malibú de Barrio Abajo (Márquez 2017).
No obstante, para corroborar estos patrones es necesario que en el futuro los resultados de esta investigación sean analizados en conjunto con otras líneas de evidencia, tales como el conjunto general de fauna de La Galepia, el material arqueobotánico, industria lítica y cerámica. De esta forma, será posible forjar una comprensión integral sobre las dinámicas de manejo del entorno en este sitio. Por otra parte, una mirada al registro histórico de la Región Caribe puede ayudar a interpretar los usos que se les dieron a los peces en esta ocupación y, de esta forma, estimar el papel que estos animales desempeñaron en los patrones alimentarios de su población prehispánica.
El registro histórico
De acuerdo con las fuentes del siglo XVI, uno de los rasgos que captó la atención de los observadores europeos y funcionarios coloniales fue la amplia oferta de fauna acuática y semiacuática presente en las llanuras del Caribe. Por ejemplo, las relaciones geográficas y crónicas de este periodo hacen referencia a ciénagas “muy fértiles” y “ríos con abundancia de pescados” en la Cuenca Baja de los ríos Magdalena y Sinú (Paleólogo et al. 1983, 203; Castellanos 2015, 139, 141)2. Algunos funcionarios coloniales incluso se preocuparon por nombrar los “muchos géneros de pescados” presentes en las ciénagas, caños y ríos de la región, resaltando entre ellos a las “doncellas”, “corvinatas, bagres y boquichicos” (Briones et al. 1983, 170).
Este contexto de abundancia y diversidad permitió a las poblaciones ribereñas y del litoral obtener grandes cantidades de peces, con las cuales aseguraban su sustento y generaban excedentes para el intercambio con otros grupos. A finales del siglo XVI, Girolamo Benzoni describió a los indígenas costeros de la provincia de Cartagena en los siguientes términos: “Esta gente tiene abundancia de frutas, pescado y todas las demás cosas requeridas para mantenerse”. Además, afirmó que “[s]us principales productos son la sal, el pescado y el pimiento; y los cargan tierra adentro, en donde hay escasez, trocándolos por otras cosas” (Benzoni 2010, 108, traducción propia).
Es factible que, para esta época, la obtención y consumo de fauna acuática tuviera igual o mayor importancia que el cultivo intensivo de plantas, al menos para los grupos que habitaban ambientes lacustres, ribereños y costeros. Esto pudo deberse a que “las condiciones imperantes” en dichas zonas resultaron “propicias para que algunas comunidades prefirieran la recolección de moluscos y la pesca a la siembra” (Langebaek 1996, 125). El cronista Pascual de Andagoya da indicios sobre este patrón en su descripción de la provincia de Cartagena a inicios del siglo XVI:
Había pocos indios que pudieran cultivar la tierra; porque el país es estéril e insalubre, una tierra baja y pantanosa, con pocos ríos y poca agua dulce. La gente, tanto hombres como mujeres, van desnudos; y tienen pocas provisiones, pero las pesquerías son abundantes. (Andagoya 2010, 80, énfasis añadido, traducción propia)
A partir de este fragmento puede deducirse que en zonas donde el potencial para la agricultura era reducido, mientras que los animales acuáticos eran abundantes y accesibles, las poblaciones tendieron a privilegiar estrategias como la recolección de moluscos, la captura de tortugas y, sobre todo, la pesca (tendencia descrita por Langebaek 1996). Tal comportamiento generó el rechazo de ciertos observadores españoles. Por ejemplo, Tomás López Medel definió a los pueblos pescadores como primitivos, perezosos y faltos de industria en su obra De los Tres Elementos (ca. 1570):
Hay gentes tan bárbaras en algunas partes de aquel Nuevo Mundo que, por su poca industria o por haberse apartado y estado de otras gentes más políticas, o no han recibido y entendido la sementera y labranza del maíz o de haraganes y flojos no quieren usar de ello ni de otro alguno, y en tanto que su comida y cuotidiano mantenimiento es pescado y éste es su pan y su vianda y todo su sustento; de manera que ni tienen otra granjería ni otro sembrar, ni otro trigo, ni otro maíz, ni cazabi, más de la pesquería que les sirve de todo. (López 1990, 154-55)
Por consiguiente, estas fuentes presentan una limitación importante, ya que se fundamentan en un discurso de diferenciación, el cual, a su vez está basado en las prácticas alimentarias (Earle 2012). En este contexto, los “indios” y otros grupos fueron marginalizados, pues sus hábitos contradecían las convenciones españolas sobre qué era comestible y cómo debía ser consumido (Saldarriaga 2009). Tal como se evidencia en el discurso de López Medel, una alimentación basada en pescado, que no incorporaba harinas producidas a partir de cultivos, fue concebida por los españoles como un signo de atraso, ignorancia, pereza y otros supuestos vicios de las gentes del Nuevo Mundo (Saldarriaga 2009). Sin embargo, estas dinámicas de diferenciación permiten aproximarse a la diversidad en las estrategias de subsistencia y hábitos de consumo de las poblaciones de la Región Caribe, la cual también se manifiesta en el registro arqueológico.
Los datos históricos indican que los malibúes emplearon una economía mixta, basada en la pesca y el cultivo intensivo de plantas, como el maíz, la yuca, entre otras (leguminosas, hortalizas y frutales) (Bernal y Orjuela 1992; Plazas et al. 1993, 1988; Ramos y Gutiérrez-Ferro s. f.; Reichel-Dolmatoff 1951). Según Bartolomé Briones de Pedraza y el cabildo de Tenerife (población del Bajo Magdalena) “Los malibúes no comen carne de ninguna forma, sino todo pescado y frutas e legumbres” (Briones et al. 1983, 168). La pesca parece haber sido la principal estrategia para la obtención del alimento entre estos grupos, pues según los funcionarios coloniales “el pescado del río y ciénagas” constituía “todo su comer” (Briones et al. 1983, 163).
Nuevamente, estas afirmaciones no deben leerse de forma literal. La información arqueológica sobre los malibúes da indicios de unas estrategias de subsistencia variadas (Bernal y Orjuela 1992; Márquez 2017). Si bien los peces tuvieron un papel importante, existe un “desencuentro” entre las líneas de evidencia, pues la idea de que el “pescado” era “todo el comer” de los malibúes no encuentra soporte en el registro arqueológico (Ramos y Gutiérrez-Ferro s. f.). El énfasis en la exclusividad del pescado entre los malibúes fue más una estrategia discursiva, empleada por los observadores coloniales para construir la imagen de un “otro” inferior (en términos de Todorov 2014) antes que una descripción rigurosa de los hábitos de estos grupos.
Los documentos disponibles para el área de estudio atestiguan este esfuerzo por construir una distinción entre malibúes y españoles, los cuales introdujeron animales y plantas traídos de Europa para su consumo. De acuerdo con las fuentes, los indígenas que eran socializados entre españoles (pues estaban sujetos a encomiendas o servidumbre) desarrollaron hábitos distintos a los indios “bozales”, sobre los cuáles no existía un dominio efectivo por parte del orden colonial (Ramos y Gutiérrez-Ferro s. f.). Por ejemplo, el cabildo de Tamalameque informó en 1579 que “carne no la comen todos [los indios], sino sólo aquellos que están criados entre españoles” (Rodríguez et al. 1983, 182). De forma similar, en cercanías de la Villa de Tenerife, “[l]os [indios] que son ladinos comen carne y todas las cosas; [mientras que] los demás bozales no la comen sino pescado y yerbas, bledos -que hay muchos-, uyamas, batatas, yuca, melones y pepinos y todas verduras” (Briones et al. 1983, 168).
En este sentido, deben tenerse ciertas reservas frente a la información que proveen los documentos. No obstante, al revisarlos con precaución, estos ayudan a contextualizar los datos arqueológicos. Por ejemplo, estos textos evidencian que los peces fueron incorporados en las prácticas de subsistencia y alimentación de ciertas poblaciones locales. Asimismo, en medio de su afán por resaltar diferencias entre grupos humanos, las fuentes indican cierta especialización ecológica entre las poblaciones indígenas de la región. Mientras los malibúes ribereños tendieron a enfocarse en la agricultura y obtención de recursos acuáticos, otros grupos privilegiaron la caza de mamíferos en las sabanas y bosques tropicales (Briones et al. 1983; Ramos y Gutiérrez-Ferro s. f.). Tal fue el caso de los caribes, grupos posiblemente relacionados con poblaciones del norte de Venezuela y las Antillas Menores (Trillos 2001). Según el Cabildo de Tenerife, los caribes acostumbraban “comer carne de monte” y se alimentaban de mamíferos que obtenían en los “arcabucos” (zonas de bosque seco tropical). Entre ellos, destacan los “puercos”, “venados”, “guartinajas”, “armadillos”, “monos”, “zorras y otras muchas sabandijas” (Briones et al. 1983, 168).
Al igual que el registro arqueológico, estos datos permiten inferir que las poblaciones de la región se enfocaron en el aprovechamiento de diferentes ambientes para obtener el alimento. En medio de esta variabilidad, ciertos grupos desarrollaron un modo de vida ribereño o lacustre, basado en la explotación de entornos acuáticos, en conjunto con la producción agrícola (Ramos y Gutiérrez-Ferro s. f.). Además, los documentos muestran que en el área de estudio los peces fueron capturados, principalmente, para ser consumidos como alimento o intercambiados entre grupos, lo cual permitió a las comunidades abastecerse de distintos recursos (Bernal y Orjuela 1992; Langebaek 1996).
De acuerdo con el registro histórico, es altamente probable que los ocupantes de La Galepia usaran los peces como alimento. Estas fuentes confirman que los patrones de consumo de los grupos malibúes privilegiaron el “pescado del río y ciénaga” como principal (más no única) fuente de alimento, pues también hicieron uso de diversas plantas cultivadas. Esta tendencia, además, es consecuente con el hecho de que los “dominios malibú”, situados en el Bajo Magdalena, han sido descritos como uno de los principales “centros pesqueros” de la Región Caribe durante el siglo XVI (Langebaek 1996, 103).
Conclusiones y perspectivas de investigación
Por medio de este estudio fue posible consolidar una primera aproximación al conjunto de restos de peces recuperado en el sitio de La Galepia y, consecuentemente, situar los resultados de este análisis en el contexto regional de la Depresión Momposina y el Bajo Magdalena. Con base en esto, fue posible concluir que los peces locales de agua dulce desempeñaron un papel fundamental en los patrones de subsistencia y alimentación de los habitantes de La Galepia durante el periodo Formativo Tardío.
A futuro, los resultados de este trabajo deben complementarse con estudios más minuciosos que permitan, por un lado, avanzar en el nivel de resolución taxonómica de este conjunto ictioarqueológico y, por el otro, realizar análisis cuantitativos más detallados, con el fin de consolidar un corpus de datos de mayor precisión sobre la distribución de las especies de peces a lo largo de la secuencia de ocupación. Entre ellos, se planea realizar otros cálculos de abundancia taxonómica y anatómica, tales como el mínimo número de individuos (MNI), mínimo número de elementos (MNE) e índices de biomasa, al igual que estudios tafonómicos y/o actualísticos.
Por medio de análisis más detallados será posible evaluar los modelos de aprovechamiento del ambiente que se han propuesto para otros sitios de la región (ej. Langebaek 1996; Langebaek y Dever 2000; Márquez 2017). Por ejemplo, se puede considerar el modelo de “explotación horizontal” propuesto por Langebaek (1996) que tiene en cuenta cierta especialización ecológica entre las poblaciones locales, sobre todo, entre aquellas que habitaron en cercanías de ambientes acuáticos. En general, este afirma que los grupos humanos tendieron a aprovechar diversas ecologías, con el fin de maximizar la obtención de recursos y asegurar el autoabastecimiento.
Este trabajo ha permitido dilucidar algunos aspectos relacionados con la subsistencia en el sitio de La Galepia. No obstante, los resultados de esta y otras investigaciones en el área de estudio deben ser cotejados, con el fin de ampliar nuestro entendimiento sobre las múltiples y complejas relaciones entre humanos, animales y entorno que tuvieron lugar en las llanuras del Caribe colombiano durante el periodo prehispánico.

