











 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
Permalink
INTRODUCCIÓN
El estudio de la estructura y función de las comunidades generalmente se ha centrado en el análisis de las interacciones negativas entre especies como depredación, parasitismo y competencia; sin embargo, las interacciones positivas como la facilitación y el mutualismo desempeñan un papel fundamental en la comprensión de los procesos y mecanismos que dan forma a esas comunidades (Hay et al. 2004). El mutualismo es una relación interespecífica donde las especies asociadas se benefician al suministrar factores nutricionales, de crecimiento, protección o transporte como en los procesos de dispersión y polinización (Boucher et al. 1982, Bronstein 1994), que son de gran importancia en el mantenimiento de la diversidad de una comunidad.
Dentro de los dispersores de semillas más importantes se encuentran los murciélagos de las subfamilias Carollinae y Stenodermatinae por su tendencia a la frugivoría, es decir, al consumo de frutos. Dentro de los murciélagos frugívoros se considera la presencia de dos categorías relacionadas con la fenología de las plantas consumidas; los frugívoros nómadas se alimentan de plantas cuya producción de frutos es masiva y de corta duración; los frugívoros sedentarios son aquellos que consumen plantas con producción continua de frutos durante todo el año (Soriano 2000).
Por otro lado, las plantas dispersadas por murciélagos presentan frutos con síndrome de quiropterocoria, es decir, con características que los atraen (Mello et al. 2005). Este fenómeno está ampliamente distribuido en plantas tropicales, entre las cuales las familias comúnmente visitadas por los murciélagos son Moraceae, Piperaceae, Solanaceae, Araceae e Hypericaceae (Calonge-Camargo 2009, Castaño Salazar 2009, Lobova et al. 2009, Estrada-Villegas et al. 2010, Ríos-Blanco y Pérez-Torres 2015, Suárez-Castro y Montenegro 2015, Aroca et al. 2016, Torres-Vásquez et al. 2016).
Dada la importancia de este tipo de interacciones, se han desarrollado métodos para su análisis como las redes de interacción, que representan la dinámica de una comunidad y la forma como interactúan las especies de dicha comunidad (Montoya et al. 2006, Bascompte 2007). Su estudio permite un mejor entendimiento de las relaciones entre complejidad y estabilidad ecológica (Dunne et al. 2002, Jordano et al. 2006, Montoya et al. 2006, Burgos et al. 2007). Así, el número de interacciones por nodo se relaciona con la estabilidad del ecosistema y la complejidad está relacionada con la heterogeneidad de las interacciones, que se ve reflejada en la presencia de una estructura anidada para el caso de las redes de interacción mutualista (Castaño Salazar 2009).
Dentro de las redes de interacción se destacan las redes mutualistas caracterizadas por presentar una especialización asimétrica denominada "estructura anidada" (Bascompte et al. 2003). Esta estructura se genera cuando las especies especialistas interactúan con un subgrupo de especies generalistas y las especies generalistas interactúan entre sí, generando un núcleo denso al que se adhiere toda la comunidad y permite la creación de rutas alternas en respuesta a perturbaciones, disminuyendo la probabilidad de que una especie desaparezca y favoreciendo la persistencia de especies raras en la comunidad. Adicionalmente, este anidamiento aumenta con la complejidad (Bascompte et al. 2003).
En Colombia las redes de interacción mutualista entre plantas y animales han sido poco estudiadas; hasta el momento se cuenta con algunos estudios enfocados en interacciones mutualistas en aves frugívoras y polinívoras en los Andes (Palacio et al. 2016, Ramírez-Burbano et al. 2017) y otros más, sobre redes de interacción entre murciélagos frugívoros y nectarívoros y plantas (Obando 2016, Zapata-Mesa et al. 2017, Mora-Beltrán y López-Arévalo 2018) , en bosques seco y húmedo tropical y bosque pre-montano en Valle del Cauca.
Sin embargo, se han desarrollado estudios en diferentes regiones y ecosistemas del país sobre análisis de dietas y dispersión de semillas que contienen la información base para la construcción de las redes de interacción mutualista; por ejemplo, se han encontrado frecuentemente interacciones entre murciélagos del género Carollia y plantas de los géneros Piper, Ficus (Ríos-Blanco y Pérez-Torres 2015), Vismia y Cuatresia (Estrada-Villegas et al. 2010). También, se ha visto una alta variabilidad en la dieta de Artibeus planirostris Spix, 1823 pues puede consumir especies de Piper, Cecropia, Solanum, Vismia y Ficus en proporciones similares (Suárez-Castro y Montenegro 2015). Finalmente especies del género Uroderma pueden consumir frutos de Piper, Ficus, Cecropia y Solanum (Calonge-Camargo 2009, Ríos-Blanco y Pérez-Torres 2015, Suárez-Castro y Montenegro 2015, Torres-Vásquez et al. 2016) .
La vereda Playa Güío (San José del Guaviare, Departamento del Guaviare, Colombia) se ubica en una zona estratégica, debido a la presencia de elementos de las regiones biogeográficas de la Amazonia, la Orinoquia y la Guayana, que hacen del estudio de su diversidad una oportunidad para avanzar en el conocimiento de la ecología de las especies que ahí habitan. Además, la vereda está compuesta por varias reservas privadas que hacen parte de una iniciativa local de la comunidad como alternativa al uso de cultivos ilícitos, que fueron erradicados definitivamente, abriendo paso a la implementación de actividades ecoturísticas encaminadas a la conservación de los bosques que se han ido regenerando (COOEPLAG c2021). El estudio de la estructura de las redes de interacción brinda información sobre la estabilidad de las interacciones en determinado ecosistema y su resiliencia ante posibles perturbaciones. Esta localidad en específico cuenta con un trabajo preliminar sobre las dietas de los murciélagos frugívoros, sin embargo, en el departamento del Guaviare no se han realizado estudios sobre redes de interacción mutualista entre murciélagos frugívoros y su recurso alimenticio. Por lo anterior, con este estudio se busca describir en la vereda Playa Güío, San José del Guaviare, las dietas y la estructura de las redes de interacción mutualista entre murciélagos frugívoros y plantas, además de hacer una comparación temporal de dichas redes.
MATERIALES Y MÉTODOS
Área de estudio
Este estudio se llevó a cabo en las reservas privadas Reserva Puerto Amor, Reserva la Galicia, Reserva Alta Galicia y Reserva Cedritos, localizadas en la vereda Playa Güío, en el municipio de San José del Guaviare, Colombia (Fig. 1). La Reserva Puerto Amor, cuenta con cerca de tres hectáreas, de las cuales el 76 % corresponde a bosque inundable destinado a la conservación y el 24 % restante se emplea en el establecimiento de cultivos e infraestructura para el alojamiento de turistas y propietarios. La Reserva la Galicia posee un área de 50 hectáreas aproximadamente, con un 42 % de vegetación secundaria, un 18 % de bosques de galería naturales con zonas sin alteración antrópica y zonas en proceso de regeneración de alrededor de quince años, y el 40 % restante se puede considerar como un mosaico natural-productivo. Por último, la Reserva Alta Galicia en la cual se encuentra inmersa la Reserva Cedritos, abarca 80 hectáreas aproximadamente, entre las que se encuentra una extensión importante de bosque inundable (46 %) y el 54 % restante es empleado para actividades productivas. Las Reservas están continuas y representan un área total de 133 hectáreas. Estas Reservas por gestión de la Cooperativa Ecoturística Playa Güío (COOEPLAG), actualmente se dedican a prestar servicios ecoturísticos y usos productivos.
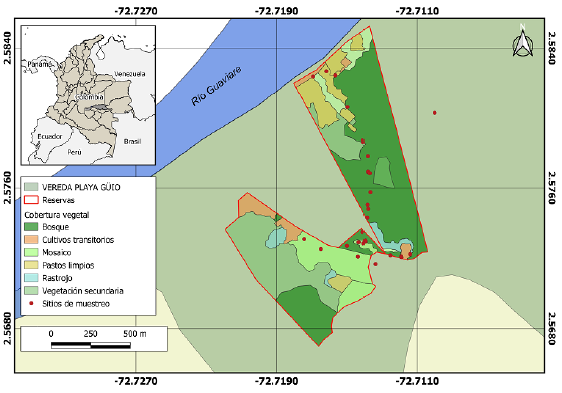
Figura 1 Mapa del área de estudio, donde se señalan los puntos de muestreo y las coberturas vegetales de la Reserva Playa Güío, municipio de San José del Guaviare, Guaviare, Colombia.
El clima de la zona es clasificado por Rudas (2009) según lo propuesto por Thornthwaite (1948) como muy húmedo con déficit de agua en el suelo por uno o dos meses, con precipitaciones promedio mensuales de 200,3 mm, precipitación anual promedio de 2628 mm y una temperatura promedio de 25,7 °C. El régimen de distribución de las lluvias es monomodal, presentándose el máximo entre los meses de abril a noviembre y el mínimo en los meses de diciembre a marzo (IDEAM 2005).
Muestreo de murciélagos
Para la obtención de los datos se realizaron dos fases de muestreo en épocas diferentes, la primera fase se realizó durante el 2014 abarcando parte de la época seca y el inicio de la época de lluvias, muestreando por cinco días en marzo, cuatro días en abril y cuatro días en mayo para un total de catorce días. La segunda fase de muestreo se realizó durante el 2017 correspondiente a la época de lluvias, muestreando nueve días en julio y nueve días en septiembre para un total de 18 días. Se instalaron entre tres y cinco redes de niebla, de doce metros de largo por tres metros de alto y con un ojo de malla de 15 mm por noche en los diferentes lugares de las reservas, ubicándolas en sitios de paso potenciales para los murciélagos; las redes se abrieron desde de las 18:00 hrs hasta las 24:00 hrs y se revisaron cada 30 minutos.
Los murciélagos capturados se retiraron de la red rápidamente y se ubicaron en bolsas de tela individuales para la posterior recolección de datos. De cada individuo se registró la hora de captura al momento de ser removidos de la red, sexo, estado reproductivo, la edad de acuerdo con la osificación de las falanges de los dedos (Mitchell-Jones y McLeish 2004, Kunz y Parsons 2009) y una identificación preliminar de la especie usando guías y claves taxonómicas (ver abajo). Adicionalmente, cada individuo se marcó en el dorso utilizando esmalte de uñas, un método de corta duración que no representa daños a largo plazo para el individuo (Mitchell-Jones y McLeish 2004, Kunz y Parsons 2009), y se liberó en el sitio de captura.
Se hizo una colección de referencia, recolectando un ejemplar por especie y aquellos que se consideren necesarios para su identificación en el laboratorio, a estos individuos se les tomaron todas las medidas estándar para murciélagos a saber: longitud total (LT), longitud de la cola (LC) longitud de la oreja (LO), longitud de la pata (LP), longitud del antebrazo (LA) y peso; estos ejemplares están depositados en la colección de mamíferos "Alberto Cadena García" en el Instituto de Ciencias de la Universidad Nacional de Colombia.
La identificación de las especies capturadas se realizó siguiendo las claves propuestas por Wilson y Reeder (2005), Gardner (2008) y Díaz et al. (2011). Para el género Platyrrhinus se utilizaron las claves propuestas por Velazco et al. (2010). Adicionalmente, el material recolectado se comparó con ejemplares de referencia de la Colección de Mamíferos "Alberto Cadena García".
Análisis de datos
Se calculó el esfuerzo de muestreo (Horas*Red) y el éxito de captura (# Individuos/Esfuerzo de muestreo) para cada salida y en total, y se construyó una curva de acumulación de especies con la información obtenida en 2014 y 2017, usando el programa Estimates (Colwell 2013), con el fin de conocer si el muestreo representaba adecuadamente la composición de la comunidad en el área. Se utilizaron los estimadores no paramétricos Jackknife de primer y segundo orden debido a que son más exactos y tienen en cuenta el número de especies en una unidad de muestreo (Moreno 2011).
Análisis de dietas
Cada murciélago se dejó en la bolsa de tela entre una y tres horas para que defecara, se tomaron las muestras fecales de dichas bolsas y se depositaron en tubos Eppendorf con etanol al 70 % para posterior identificación y conteo de semillas en el laboratorio. Las muestras fecales se limpiaron mediante lavados con etanol al 70 % y se dejaron secar a temperatura ambiente por una noche.
Para la identificación de las semillas de los géneros de Cecropia se siguió la clave propuesta por Linares y Moreno-Mosquera (2010); las demás especies se identificaron con una colección de referencia de plantas y semillas realizada en el sitio de estudio y con la información disponible en la Carpoteca del Herbario Nacional Colombiano, del Instituto de Ciencias Naturales de la Universidad Nacional de Colombia. Adicionalmente, de cada murciélago recolectado se preservó el contenido estomacal con el fin de contar con la mayor cantidad de semillas posible.
Teniendo en cuenta que los datos de la comunidad estudiada representan el uso de un recurso, se analizaron también índices de amplitud y traslape de nicho. Para conocer la amplitud de nicho de cada especie se calculó el índice de Levins Estandarizado (Krebs 2013), cuyos valores van de cero a uno, donde cero indica que la población usa solo un recurso y uno que la población utiliza los diferentes recursos en igual proporción. Se calculó por medio de la siguiente ecuación:
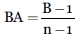
Donde, BA: Índice de Levins estandarizado,
B: Amplitud de nicho trófico B= 1/ Σpj 2
n: Número de ítems presa
pj2: Frecuencia de cada ítem en la dieta de los individuos muestreados
El porcentaje de solapamiento entre las dietas de las diferentes especies de murciélagos se calculó mediante el Índice de Morisita, cuyos valores van de cero a uno, donde cero es la ausencia de traslape y uno indica un traslape completo entre las dietas de las especies. Valores mayores a 0,6 son indicadores de un traslape significativo mientras que los menores a 0,3 son valores no significativos (Krebs 2013):
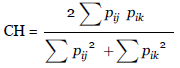
CH: Índice de Morisita
p ij : Proporción del recurso i del total de recursos utilizados por la especie j
pik: Proporción del recurso i del total de recursos utilizados por la especie k
Análisis de redes tróficas
Se elaboraron las matrices y redes en las que se evidencia la interacción del murciélago y su respectivo recurso alimenticio para 2014 y 2017, y en total de los dos años. Se estimó el tamaño de la red, es decir, el número de especies en la red en determinado momento, la modularidad donde valores de 1,0 indican una modularidad completa, mientras que valores de 0,0 indican que la red no es modular, para su cálculo se utilizó el programa MODULAR (Marquitti et al. 2014); la robustez se estimó para evaluar el efecto de la extracción aleatoria de especies de cualquiera de los grupos, valores de 1,0 indican un grado de robustez alto, mientras que valores de 0,0 indican robustez baja donde el sistema colapsa al eliminar pocas especies (Burgos et al.2007); finalmente se calculó el algoritmo de anidamiento NODF propuesto por Almeida-Neto et al. (2008), con el fin de observar el grado de acoplamiento y estabilidad de la red; este algoritmo varía de cero a 100, siendo cero cuando la red no está anidada y 100 cuando está completamente anidada. Estos últimos dos estimadores se calcularon utilizando el paquete Bipartite 2.02 (Dormann et al. 2008) del programa estadístico R en la plataforma RStudio (RStudio Team c2020) A partir de ello, se realizaron las comparaciones correspondientes a cada año.
RESULTADOS
Se capturaron en total 485 individuos (185 en 2014 y 300 en 2017) pertenecientes a las familias Phyllostomidae, Vespertilionidae y Emballonuridae, y a 34 especies de las cuales 18 son frugívoras. La familia mejor representada es Phyllostomidae con 25 de las 34 especies, seguida de Em-ballonuridae con tres especies y Vespertilionidae con una especie (Anexo 1, material suplementario).
Se obtuvo un alto porcentaje de especies frugívoras (52,94 %). Se encontraron, aunque en menor medida, murciélagos insectívoros (29,41 %), omnívoros (8,82 %), hematófagos (2,94 %), melívoros o nectarívoros (2,94 %) y carnívoros (2,94 %) (Anexo 1, material suplementario).
La curva de acumulación de especies proyecta un mayor número de especies para la zona (Fig. 2); pero la representatividad del muestreo estuvo entre 83,78 % según Jac-kknife 1 y 90,14 % de acuerdo con Jackknife 2 del total de especies estimado.
Análisis de dietas
Las familias de plantas típicamente consumidas por los murciélagos durante este muestreo fueron Piperaceae, Moraceae, Urticaceae, Hypericaceae, Solanaceae y muestras que no pudieron asignarse a una familia específica (Anexo 2, material suplementario). De estas familias se reconocieron 24 especies de las cuales se identificaron a nivel de especie Cecropia membranácea Trécul, Vismia baccifera (L.) Triana & Planch, Vismia guianensis (Aubl.) Pers., Piper peltatum L. y Solanum jamaicense Mil, y a nivel de género Piper (ocho morfotipos), Ficus (seis morfotipos), Solanum (dos morfotipos), Cecropia (un morfotipo) y un morfotipo no identificado.
En cuanto a la amplitud de nicho de Levins (Tabla 1) los mayores valores se encuentran en las especies Carollia castanea (0,2671), A. planirostris (0,1840), Platyrrhinus brachycephalus (0,1739) y C. perspicillata (0,1440), indicando que sus dietas tienden a ser generalistas pues consumen frutos de varias especies, sin embargo, el valor no se encuentra muy alejado del cero pues se alimentan de uno de los recursos en mayor proporción.
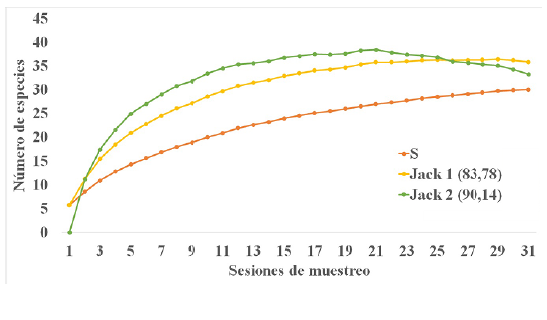
Figura 2 Curva de acumulación de especies de los murciélagos capturados en los dos periodos de muestreo 2014 y 2017, en Playa Güío, San José del Guaviare. Las líneas amarilla y verde representan los valores estimados por Jackknife 1 y 2 respectivamente, cuya representatividad se encuentra en paréntesis. La línea naranja representa los valores obtenidos durante los muéstreos realizados.
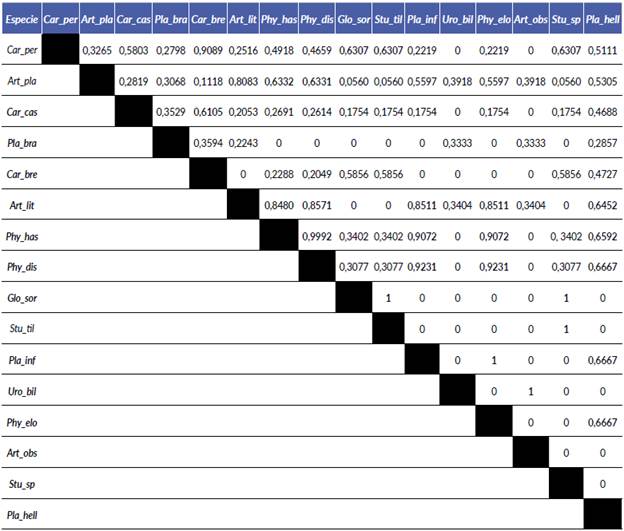
El mayor valor del índice de Morisita se presenta en especies de frugívoros sedentarios donde se encuentran los géneros Carollia, Sturnira, Phyllostomus y Glossophaga, y entre los géneros Artibeus, Platyrrhinus y Uroderma que se encuentran dentro de los frugívoros nómadas, para los dos casos, los valores son mayores a 0,6 lo que indica un traslape significativo y en el caso de Sturnira con Glos-sophaga (sedentarios) y Artibeus con Uroderma (nómadas), el traslape es completo (Tabla 2).
Análisis de redes tróficas
Durante la temporada seca en 2014 se encontró que la red está constituida por cinco especies de murciélagos que se alimentan de seis especies de plantas. En la temporada de lluvias, se adicionan a la red once especies de murciélagos y catorce especies de plantas (Fig. 3). Para ambas temporadas los frutos pertenecientes a los géneros Piper y Vismia estuvieron entre los más consumidos por los murciélagos, especialmente por las especies Carollia perspicillata y Artibeus planirostris (Figs. 3 y 4).
Tabla 1 Valores de amplitud de nicho de Levins (B) y Levins estandarizado (BA) para las especies de murciélagos en la comunidad de la vereda Playa Güío.
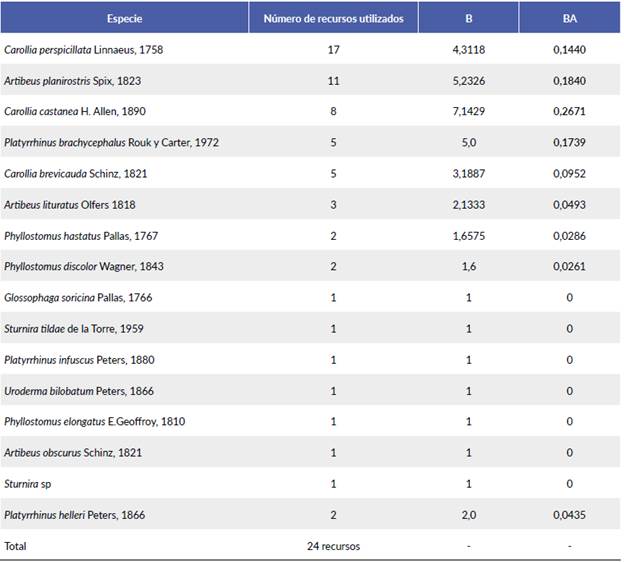
Tabla 2 Valores de traslape de nicho del índice de Morisita simplificado (CH) para las especies de murciélagos en la comunidad de la vereda Playa Güío. Las abreviaciones corresponden a los nombres de las especies Carper (Carollia perspicillata), Art_pla (Artibeus planirostris), Carcas (Carollia castanea), Pla_bra (Platyrrhinus brachycephalus), Carbre (Carollia brevicauda), Art_lit (Artibeus lituratus), Phy_has (Phyllostomus hastatus), Phydis (Phyllostomus discolor), Glo_sor (Glossophaga soricina), Stutil (Sturniratildae), Pla_inf (Platyrrhinus infuscus), Uro_bil (Uroderma bilobatum), Phyelo (Phyllostomus elongatus), Art_obs (Artibeus obscurus), Stu_sp (Sturnira sp) y Pla_hell (Platyrrhinus helleri).
La red de interacción gráfica de los datos registrados en 2014 da como resultado seis especies de plantas utilizadas por cinco especies de murciélagos para un tamaño total de once especies. Entre las primeras, la más consumida por los murciélagos es V. baccifera con cinco conexiones, seguida de dos especies del género Piper con cuatro y tres conexiones respectivamente (Fig. 3); en cuanto a los murciélagos, A. planirostris es la especie que consume más recursos (cuatro). La modularidad fue de 0,28, la robustez fue de 0,68 y el anidamiento fue 54,67, este valor indica que la red se encuentra bien anidada.
Por otra parte, la red de interacción del muestreo en 2017 muestra 20 especies de plantas que son consumidas por 16 especies de murciélagos, en total el tamaño de esta red fue de 36 especies. De las plantas más consumidas por los murciélagos se destacan C. membranacea con nueve conexiones, Piper sp5 con nueve conexiones, Ficus sp4 con cinco conexiones y finalmente Vismia baccifera y Cecropia engleriana con cuatro y tres conexiones respectivamente (Fig. 3). En cuanto a los murciélagos, Carollia perspicillata se alimenta de la mayor cantidad de recursos (16 especies de plantas), seguida de Artibeus planirostris y Carollia castanea que utilizan siete recursos cada una. Para esta red de interacción, se destaca la importancia de C. perspicillata al representar la mayor cantidad de conexiones de la comunidad de murciélagos y Cecropia membranacea por ser consumida por la mayor cantidad de murciélagos. En cuanto a las medidas calculadas; la modularidad y la robustez fueron de 0,41 y 0,65, respectivamente; el anidamiento corresponde a 50,71, menor que el valor alcanzado por la red de 2014; este valor indica que la red tiene una estructura anidada.
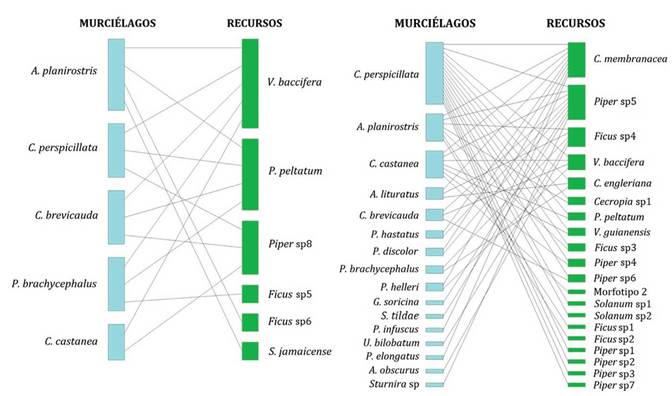
Figura 3 Redes de interacción mutualista entre murciélagos frugívoros y su recurso alimenticio, para los periodos 2014 (izquierda) y 2017 (derecha), en la vereda Playa Güío, San José del Guaviare.
Por último, la red total para la vereda Playa Güío (Fig. 4), adiciona cuatro especies de plantas y se mantiene la misma cantidad de especies de murciélagos (16), para un tamaño total de 40 especies. Los valores de modularidad y robustez corresponden a 0,4160 y 0,6488, respectivamente, y el valor de anidamiento (NODF) para este caso es 46,63; en este caso se agregan nuevas especies con una conexión, pero no se aumenta el número de conexiones.
DISCUSIÓN
La comunidad estudiada presenta una estructura cuyo patrón coincide con el de los ensambles de murciélagos en el Neotrópico, en los que la familia Phyllostomidae es la más abundante y la mayoría de las especies que la componen se incluyen dentro del gremio de los frugívoros (Fleming et al. 1972, Bolaños-Arrieta 2013, Mendoza-Saenz et al. 2017), sin embargo, esta situación se debe también al efecto del tipo de muestreo.
La red de interacción de la vereda Playa Güío comprende un total de 16 especies de murciélagos y 24 de plantas, el número de especies frugívoras en esta interacción solo es superado por la información obtenida en Barro Colorado (22 especies) y es superior a otras ocho localidades de Brasil, Costa Rica y Perú analizadas por Mello et al. (2011) a partir de estudios de dietas de murciélagos frugívoros Neotropicales. Así como para frugívoros de Bosque Seco y Bosque Húmedo Neotropical, estudiados por Zapata-Mesa et al. (2017). En cuanto al número de especies de plantas nuestros valores son intermedios entre los estudios anteriormente reportados, los cuales fluctúan entre seis y 47.
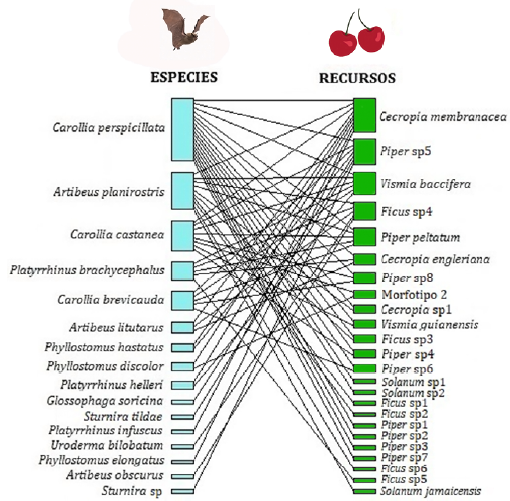
Figura 4 Red de interacción mutualista entre murciélagos frugívoros y su recurso alimenticio para la vereda Playa Güío en San José del Guaviare, para los datos unificados de las dos temporadas.
Con respecto a la composición de especies en la red de interacción los géneros de murciélagos más abundantes y con la mayor cantidad de interacciones son Artibeus y Carollia, similar a lo encontrado por Zapata-Mesa et al. (2017) en Bosque Seco, mientras que para el Bosque Húmedo las especies mejor representadas se encuentran en los géneros Platyrrhinus y Dermanura. Por su parte, los frutos más consumidos durante nuestro estudio se incluyen en los géneros Cecropia, Piper, Vismia, Ficus y Solanum, similar a lo encontrado por Zapata-Mesa et al. (2017) en Bosque Húmedo.
A diferencia de lo encontrado por Zapata-Mesa et al. (2017), donde las especies de plantas encontradas con mayor frecuencia no presentan un gran número de interacciones, en nuestro estudio las plantas más encontradas en las heces de los murciélagos son las especies que tienen la mayor cantidad de interacciones (Cecropia membranacea y Piper sp5).
Con respecto a las métricas estimadas, tanto los valores de modularidad como los de robustez de las redes de interacción son altos, se encuentran dentro del intervalo reportado para diferentes localidades del Neotrópico (Mello et al. 2011) y concuerdan con Zapata-Mesa et al. (2017); las estructuras modulares son importantes pues permiten minimizar el impacto de la pérdida de especies y aumentan la robustez de las redes, haciendo que a pesar de la pérdida de algunas especies el sistema no colapse.
Por su parte, los valores del algoritmo de anidamiento (NODF) se encuentran también en el intervalo reportado para el Neotrópico, entre 0,39 y 0,75, por Mello et al. (2011). Los valores no tan altos en el anidamiento se relacionan con el tipo de mutualismo difuso porque las especies generalistas tienden a consumir frutos de gran cantidad de plantas y las plantas a su vez se benefician de los servicios de dispersión de múltiples especies de frugívoros (Mello et al. 2011, Zapata-Mesa et al. 2017). A partir de estas medidas, se evidencia la formación de núcleos o módulos que permiten que especies poco abundantes como en nuestro caso Platyrrhinus infuscus y Phyllostomus elongatus, así como algunos morfotipos de Piper y Solanum, se mantengan en el sistema.
Con respecto al análisis temporal de las redes, existe una alta variación en el número de especies relacionadas y la cantidad de las interacciones, la red construida al inicio de la temporada de lluvias en 2014, presenta una menor cantidad de especies que la red en la temporada de lluvias del 2017. Así mismo los valores de modularidad, robustez y anidamiento son diferentes, la modularidad es menor en la primera pues no se alcanzan a formar módulos por la baja cantidad de especies, mientras que la robustez y el anidamiento son mayores indicando una red altamente cohesiva y resistente a las perturbaciones (Bascompte et al. 2003, Bascompte y Jordano 2006).
Por su parte, la modularidad de la red de la temporada de lluvias de 2017 aumenta al aumentar el número de especies y de interacciones, y la robustez y el anidamiento disminuyen pues se adicionan nuevas especies y también nuevas interacciones entre diferentes especies (Bascompte et al. 2003). Las diferencias temporales entre las redes podrían estar relacionadas con la fenología de las plantas, donde los picos de fructificación tienden a concentrarse en la época de lluvias (Suárez-Castro y Montenegro 2015); esto se ve reflejado en el aumento del tamaño de la red tanto en especies de murciélagos como en especies de plantas.
En las redes de interacción se observa también una estrecha relación entre los murciélagos de los géneros Carollia y las plantas del género Piper. Este patrón se ve a lo largo de diferentes localidades tanto de Colombia (Estrada-Villegas et al. 2010, Ríos-Blanco y Pérez-Torres 2015, Suárez-Castro y Montenegro 2015) como del Neotrópico, pues se ha encontrado una sobreposición en la distribución espacial de estas especies (Lobova et al. 2009).
Lo anterior se relaciona con la amplitud de nicho pues indica una posible preferencia o selectividad hacia uno de los recursos consumidos, por ejemplo, especies como C. castanea, C. perspicillata y A. planirostris tienden a ser generalistas, pero consumen en mayor proporción uno de los recursos, tal y como se ha observado en otros estudios realizados en el Neotrópico (Fleming 1986, Marinho-Filho 1991, Giannini y Kalko 2004, Lou y Yurrita 2005).
Los valores de traslape de nicho indican que las especies tienen dietas muy similares, lo que se relaciona también con la agrupación en las categorías nómada y sedentario de los murciélagos frugívoros (Soriano 2000), pues se encontró que las especies al interior de estos grupos tienen los valores más altos de traslape.
Las especies del género Cecropia representan recursos importantes para especies de los géneros Artibeus y Platyrrhinus, similar a lo encontrado por Suárez-Castro y Montenegro (2015) en la Orinoquia colombiana. Estos murciélagos, considerados frugívoros nómadas, presentan un traslape amplio de nicho indicando una fuerte relación con especies de este género de plantas (Tabla 2). En estudios realizados en el país y a lo largo del Neotrópico, se muestra que los murciélagos frugívoros tienden a consumir frutos con patrones de fructificación continuos, pero son flexibles y responden positivamente a cambios en la distribución y abundancia de estos (Lobova et al. 2009, Suárez-Castro y Montenegro 2015).
Cabe resaltar que uno de los aspectos más importantes de nuestro trabajo radica en que las especies de plantas dispersadas por los murciélagos de esta zona, como las de los géneros Cecropia, Piper, Vismia y Ficus, constituyen los taxones que aportan la mayor cantidad de registros, tienen frutos con gran cantidad de semillas, presentan fructificación continua durante el año (Suárez-Castro y Montenegro 2015, Torres-Vásquez et al. 2016) y son especies pioneras cuya dispersión es fundamental en los procesos de regeneración de los bosques en el Neotrópico (Galindo-González 1998, Lobova et al. 2003).
Además, se ha encontrado en la amazonia colombiana que al dejar un parche abandonado tres años después del proceso de tala y quema, el 87 % de individuos que han colonizado el área, corresponden a especies de estos géneros (Lobova et al. 2009). El papel de los murciélagos en la dispersión de este tipo de plantas pioneras ha sido reconocido desde hace varias décadas (Charles-Dominique 1986, Fleming 1986, Galindo-González 1998). Así, la presencia de estas especies y las características de las redes de interacción obtenidas en este estudio, permiten evidenciar que han sido satisfactorios los esfuerzos por recuperar y conservar los bosques presentes en la vereda Playa Güío tras la erradicación de los cultivos ilícitos.
CONCLUSIONES
Las reservas de la vereda Playa Güío mantienen una red de interacción de murciélagos frugívoros caracterizada por una alta riqueza de especies en comparación con varias localidades del Neotrópico, y con una variación temporal asociada con la época climática. Las especies centrales tanto de plantas como de murciélagos son generalistas y se encuentran asociadas a procesos de regeneración natural importantes para los objetivos de conservación de las reservas, además, las métricas indican que esta red de interacción mutualista es estable y se puede mantener en el tiempo.

