











 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Citado por Google
Citado por Google  Similares em
SciELO
Similares em
SciELO  Similares em Google
Similares em Google 
 Permalink
PermalinkIntroduction
In Ethiopia, the livestock subsector makes significant contributions to the national income and the livelihoods of households (CSA 2016), contributing 15‒17% of gross domestic product (GDP), 35‒49% of agricultural GDP and 37‒87% of household incomes (ILRI 2010; Behnke and Menagerie 2011). However, livestock’s contribution is below potential because of inadequate nutrition, with the critical feed nutrient, crude protein (CP), of herbaceous plants declining during the dry season, leading to prolonged periods of under-nutrition of livestock (Yayneshet 2010). This shortage of CP could be mitigated through integration of multipurpose forage legume species such as lupin into feeding systems. Narrow-leafed sweet blue lupin (Lupinus angustifolius L.), hereafter referred to as narrow-leafed lupin, is adapted to low external input production systems, produces higher yields than local native lupins and is highly palatable to livestock, with Vitabor, Probor, Sanabor and Bora giving the highest grain yields. Vitabor and Sanabor are nationally registered varieties for use as multipurpose pulse crops in Ethiopia. These varieties have potential as both feed for livestock and food for humans and are suitable for intercropping with maize (Alemayehu et al. 2014; Yeheyis et al. 2015). While some research into adaptability and feeding value of narrow-leafed lupin has been conducted, no attempt has been made to characterize varieties. This study was conducted to characterize different varieties of narrow-leafed lupin based on growth characteristics and physical features.
Materials and Methods
Description of the study area
The study was conducted during June‒November 2016 at Gondar Agricultural Research Station, which is located near Dabat town (13°05' N, 37°50' E; 2,740 masl) in the Dabat District of North Gondar Administrative Zone, Ethiopia (Figure 1). Rainfall and temperature data calculated for the research station are presented in Figure 2. The soil in the area is a well-draining Nitosol with low organic matter content (Table 1).


Figure 2 Climatic data calculated for the Gondar Agricultural Research Station, from Climate Hazard Group InfraRed Precipitation with Station (CHIRPS); long-term temperature data from WorldClim Version 2. Long-term data refer to 2007‒2017. Data accessed through CCAFS GCM Downscaled Data Portal ccafs-climate.org/weather_stations.

Experimental design and treatments
The experimental design was a randomized complete block design with 4 replications. Plot size was 2.1 × 3.6 m (7.56 m2), with 1 m between plots and replications. The land was plowed 3 times before planting to provide a fine seedbed. Seed was planted on 7 July 2016 in rows with 7 cm between plants within rows and a row spacing of 30 cm. Di-ammonium phosphate (DAP) fertilizer was applied at the rate of 100 kg/ha to provide 18 kg N and 22 kg P/ha at planting. Hand-weeding was conducted at the seedling stage and again prior to flowering.
Data collection
For sampling purposes half of each plot (subplot) was used to determine forage yield and the other half to determine grain production. Time from planting to flowering was assessed when 50% of plants in a plot were flowering and time to maturity when 90% of plants had reached maturity, i.e. set seed. Harvesting to determine forage biomass yield was performed at the 50% flowering stage from the middle 4 rows of half the subplots by cutting at ground level. Fresh biomass yield was recorded in the field and 500 g-samples were taken from each plot. Number of leaflets per plant was counted based on green leaves of 5 tagged plants at 50% flowering. These samples were separated manually into leaf and stem, before oven-drying at 65 °C for 72 hours until constant weight was obtained. Leaf:stem ratio was calculated. Plants on the remaining subplots were harvested at ground level at the optimum seed harvesting time (at 12.5% moisture) and seed was removed by threshing. Fresh weight of seed was determined and samples of seed were air-dried to constant weight.
Phenological and morphological parameters
Plant flower color was assessed based on close observation of flowers and recorded using a subjective scale from white to blue. Plant height was measured on 10 plants per plot, selected at random, from the soil surface to the tip of the longest leaflet at the forage harvesting stage. Number of leaflets and number of branches per plant were determined on 5 randomly tagged plants/plot at 50% flowering. Number of pods/plant, pod length and number of seeds/pod were measured on 5 randomly tagged plants/plot at maturity. To determine 1,000-seed weight, 1,000 seeds were taken randomly from the harvested seed lot for each plot and weighed.
Statistical analysis
The collected data were subjected to analysis of variance (ANOVA) using the general linear model (GLM) procedure of SAS (2016), Version 9.1.3. Mean separation was conducted using the least significant difference test (LSD), when the ANOVA showed significant difference among treatments. Simple pair-wise correlation and regression analyses were conducted using JMP 13 (SAS 2016) to show the relationship between growth, yield and yield components. The statistical model used for data analysis was:
Yij = µ+ti+bj+ eij
where: Yij = the response variable (the observation in jth block and ith treatment); µ = the overall mean; ti = the treatment effect; bj = the block effect; and (gradient) eij = the random error.
Results
Days to flowering and maturity and flower color
Variety significantly (P<0.01) influenced both days to flowering and days to maturity (Table 2). The longest time to flowering was for Bora (87.5 days) and the shortest for Haags blaue (74.5 days), while corresponding times to maturity were 147 and 134 days, respectively. Haags blaue variety was seriously affected by lack of moisture at the flowering stage and aborted flowers before it reached maturity. Varieties Probor, Vitabor, Haags blaue and Borlu showed blue flower color, while flowers of Bora and Sanabor were bluish white; only those of Boregine were white.
Table 2 Effects of variety on days to flowering (FD), plant height (PH), branches per plant (NB), leaflets per plant (NL), leaf:stem ratio (L:S), days to maturity (MD) and forage dry matter yield (DMY) of 7 narrow-leafed lupin varieties.
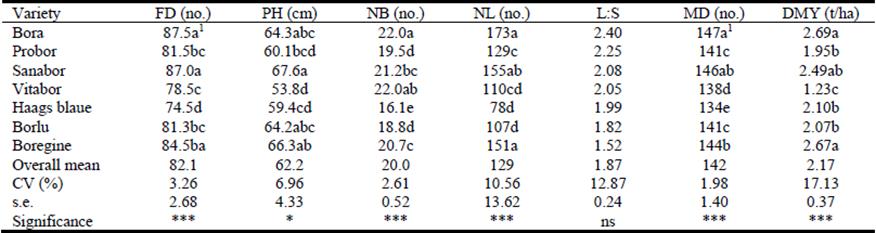
1Values within columns followed by different letters are significantly different.
Plant height
Plant height was significantly (P<0.05) affected by variety with tallest plants for Sanabor (67.6 cm) and shortest for Vitabor (53.8 cm; Table 2).
Number of branches and leaflets per plant
Lupin has an indeterminate growth habit and can continue to initiate new lateral branches with flower spikes after it reaches the reproductive phase. The highest number of branches was observed for Bora (22) and lowest for Haags blaue (16) (P<0.01). Number of leaflets per plant followed a similar pattern with highest number for Bora (173) and lowest for Haags blaue (78) (P<0.01) (Table 2).
Forage dry matter yield
Forage dry matter yield at flowering varied significantly (P<0.01) among varieties with yields varying from 2.69 t/ha for Bora to 1.23 t/ha for Vitabor. While leaf:stem ratio varied from 2.40 for Bora to 1.52 for Boregine, differences were not significant (P>0.05) (Table 2).
Number of pods per plant, seeds per pod and seed (grain) yield
The number of pods per plant was not significantly (P>0.05) affected by variety with an overall mean of 11.0 and a range of 8.1 to 16.9 pods/plant (Table 3). Similarly, pod length was not affected by variety. However, number of seeds per pod was significantly (P<0.001) affected by variety ranging from 3.18 to 4.15.
Table 3 Effects of variety on number of pods per plant (NP), pod length (PL), number of seeds per pod (NS), grain yield (GY) and 1,000-seed weight (TSW) of 7 narrow-leafed lupin varieties.

1Values within columns followed by different letters differ significantly (P<0.05).
Grain yields varied widely between varieties (P<0.01), ranging from 1,901 kg/ha for Boregine to 786 kg/ha for Haags blaue (Table 3). Thousand-seed weight also varied significantly between varieties from 125 g for Sanabor to 91 g for Borlu.
Correlation between plant parameters and grain yield
Almost all agronomic attributes were strongly correlated with grain yield. Plant height (r = 0.59**), days to flowering (r = 0.51**), days to maturity (r = 0.56**), number of seeds per pod (r = 0.92**) and number of branches per plant (r = 0.52**) were positively related to grain yield (Table 4).
Table 4 Correlations between plant parameters, yield and yield components of 7 narrow-leafed lupin varieties.
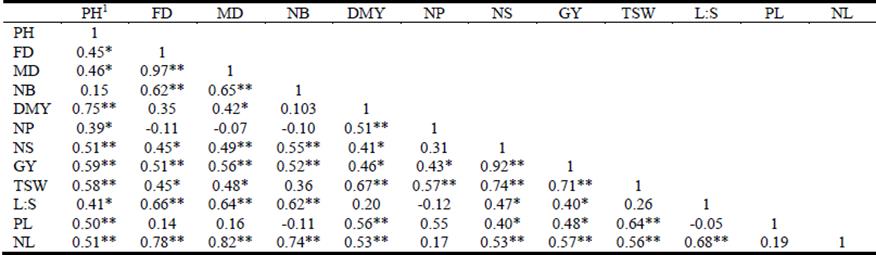
1PH = plant height; FD = days to flowering; MD = days to maturity; NB = number of branches per plant; DMY = forage dry matter yield; NP = number of pods per plant; NS = number of seeds per pod; GY = seed yield; TSW = 1,000-seed weight; L:S = leaf:stem ratio; PL = pod length; NL = number of leaflets per plant.
Discussion
This study has provided useful information on the possible yields of forage and seed from narrow-leafed lupin varieties in Ethiopia. The variation in these parameters among different varieties suggests that there is room for selection if the aim is to establish breeding programs to improve yields.
Days to flowering
Mean time to flowering for Sanabor of 87 days was similar to that reported by Herbert (1977), who showed that flowering began at about 84 days after sowing and most plants were flowering by 87 days.
Flower color
We observed that Boregine was the only variety that had white flowers, which was in agreement with Tarekegn (2016), who indicated that Boregine displayed different flower colors including white. Lupin flowers do not contain nectar, but insects are attracted by their bright color and presence of pollen (Kurlovich et al. 2011). Klamroth et al. (2011) suggest that different flower colors in lupins might improve agronomic characters such as seed yield by attracting pollinating insects.
Days to maturity
As might be expected, varieties that flowered early also reached maturity before other varieties. Berger et al. (2011) indicated that delayed phenology has a huge effect on yield. Based on a study with white lupins, where varieties had set seed by 4.5 months, Azeze (2016) suggested that, as compared with other pulse crops, white lupin must remain in the field for 7‒8 months until the plant fully matures. Mulugeta et al. (2015) suggested that the presence of highly significant variation in maturity between Sanabor and the Ethiopian landraces indicates the possibility of using Sanabor as a source of genes to advance flowering in late-maturing Ethiopian landraces.
Plant height
The difference in plant height between varieties was in agreement with Tarekegn (2016), who stated that the tallest plants were recorded in Bora and Sanabor and the shortest plants in Vitabor. We recorded an overall mean plant height of narrow-leafed lupin varieties of 62 cm (range 54‒68 cm) as opposed to the mean height of 82 cm reported for the same varieties by Rudloff (2011). This discrepancy might be due to environmental variations of experimental areas. However, Edwards et al. (2011) reported that plant height of other narrow-leafed lupin varieties varied from 20 to 100 cm.
Number of branches per plant
Growth habit of lupin is characterized by a high number of branches per plant when plants have access to adequate moisture, nutrients and sunlight (Edwards et al. 2011). While we recorded an average of 20 branches per plant, Rudloff (2011) reported for Boruta lines (also L. angustifolius) that the maximum number of branches per plant was 7. This might be a function of agro-ecological differences and management variations. It was interesting that there was no significant correlation between plant height and number of branches per plant. In contrast with this study, Klamroth et al. (2011) found that the strongest vigor was observed in Boregine, followed by Haags blaue, with Probor showing only marginal strength.
Seeding
The mean grain yields of all varieties in this study were below expected levels which might be due to the poor soil in the study area. Root elongation of narrow-leafed lupin slows when the pH (CaCl2) increases from 5.5 to 6.0, and at a pH greater than 6.0 the root surface can be damaged as surface cells are peeled off and root hair formation slows (Edwards et al. 2011). In this experiment the pH of the soil was 6.3. In addition, the flowering process of lupin is affected by high temperatures which cause blasting of flowers and a subsequent grain yield reduction (Klamroth et al. 2011); blasting of flowers was also observed in this experiment.
Late-flowering varieties such as Boregine and Sanabor had the highest grain yields as they had the highest numbers of pods per plant, seeds per pod and 1,000-seed weight compared with the other genotypes. Late flowering and hence late seed set could have negative consequences in some circumstances as the grain-filling period may coincide with increasing temperatures and decreasing soil water availability with associated decrease in availability of nutrients, leading to terminal drought and yield reductions (Thornton et al. 2014). Lupin appears to rely on current photosynthesis to fill grain, as opposed to e.g. wheat, which remobilizes stored carbon fixed earlier in the season. This phenomenon may account for high pod abortion, poor yield stability and low harvest index (Sandaña et al. 2009; Edwards et al. 2011).
The ranking of varieties in terms of grain yield was at variance with the previous report of Yeheyis et al. (2012), who reported yield rankings in descending order: Vitabor, Sanabor, Probor and Bora, in northwestern Ethiopia. These differences could be due to variation in agro-ecologies among the study areas. Tarekegn (2016) also found that Sanabor gave higher grain yield than other tested narrow-leafed lupin genotypes, while Yeheyis et al. (2015) indicated that Boregine and Sanabor were the best-performing varieties among the present tested genotypes. While Klamroth et al. (2011) showed highest yields for Boregine, Mekonnen et al. (2016) showed that varieties Sanabor, Vitabor, Bora and Probor gave comparable grain yields and 1,000-seed weights when intercropped with maize.
Kurlovich et al. (2011) also reported no significant difference in number of pods per plant among narrow-leafed lupin varieties. Number of seeds per pod is a major yield component and breeding target in lupins that shows great variation and is invaluable for genetic improvement (Yang et al. 2016). Our observations indicated that number of seeds per pod had a much greater influence on grain yields than number of pods per plant. Pods can produce 3 to 6 seeds, depending on the location on the plant, with pods produced on higher branches containing fewer seeds (Edwards et al. 2011). Though the overall mean number of seeds per pod obtained from this experiment was less than that reported by Rudloff (2011), significant differences (P<0.001) in number of seeds per pod were exhibited among varieties as shown by Yang et al. (2016). Differences were not surprising as this is a highly complex trait determined by the number of ovules per ovary, the proportion of fertile ovules (ovule fertility), the proportion of ovules fertilized and the proportion of fertilized ovules that develop into seeds. The proportion of ovules fertilized is determined by the fertilization process, which is dependent on many factors such as pollen sterility, the amount of pollen deposited on the stigma, pollen grain germination (Schreiber and Dresselhaus 2003) and pollination conditions (Brown et al. 1990). Among the tested varieties Klamroth et al. (2011) discovered that flower number and pod set in Boregine were lowest, resulting in low seed production. In another experiment conducted by Mekonnen et al. (2016) numbers of narrow-leafed lupin seeds per pod and pods per plant did not show significant difference among treatments when they were intercropped with maize.
Thousand-seed weight
The finding that Sanabor, Boregine and Bora gave the highest 1,000-seed weights (125, 121 and 118 g, respectively) was in parallel with the finding of Stoddard and Lizarazo (2011). These high 1,000-seed weights could be attributed to large accumulation of assimilates due to a long maturity period. The overall mean 1,000-seed weight (107 g) obtained from this experiment was much lower than the 130 and 143 g reported by Edwards et al. (2011) and Rudloff (2011), respectively, the differences being probably due environmental conditions.
Forage dry matter yield
The pattern of forage DM yield, i.e. highest in Boregine and lowest in Haags blaue, was in agreement with findings of Yeheyis et al. (2011). On the other hand, other studies, e.g. by Tarekegn (2016), showed no difference in mean DM yields per season of narrow-leafed varieties. We also found that the longer plants took to mature the higher were the forage and seed yields as there was sufficient time to accumulate biomass and set greater amounts of seed. Weerakoon and Somaratne (2013) showed that variety had a statistically significant effect on biomass yield. While we did not analyze forage or seed for crude protein concentration, other workers have shown that forage of narrow-leafed lupin contains CP concentration of 15.8‒30% at maturity, which makes it suitable for animal feeding (Yeheyis 2012). A 70-day study conducted with Washera sheep using narrow-leafed lupin grain as a supplement at 290 g/head/day showed that the animals gained 74 g/head/day (Yeheyis et al. 2012). The same study showed that cv. Sanabor has potential to be a suitable substitute for commercial concentrate feed supplement in Ethiopia and concludes that lupin seed is a highly nutritious product which can be used as a protein and energy supplement for livestock during dry times.
Conclusions and Recommendations
This study has shown that the narrow-leafed lupin varieties Boregine, Sanabor and Bora appear better suited to the study area than the other varieties tested because of better forage DM and grain yields. These varieties seem promising for the development of sustainable forage production systems with a limited use of external inputs as they are common in Ethiopian smallholder agriculture. However, performance over a range of seasons and locations is necessary to confirm these initial findings. Future research on narrow-leafed lupin should be directed at a more detailed study of its grain yield components, quality of both forage and seed, the improvement of its agronomy and the possibility of its utilization as forage or green manure crop, as well as to the testing of promising genotypes in diverse locations.

