











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
Worldwide, several research studies describe the role that plant growth plays in plant biomechanics and in current and future forest management decisions. These aspects are based on information obtained from inventories (Benavides et al., 2019) and are estimated considering development and performance models obtained from mathematical scaling. Through this method, one or several parts of a species of interest are evaluated, in order to estimate the overall development of a component that makes up part of it (Jagodziński et al., 2017). In general, the most commonly used mathematical models in forestry are allometric models, which help to establish the development of correlative dasometric relationships among forest species in a given region (Hernández-Ramos et al., 2019).
In forestry, measuring the total height (TH) of the stand is a common practice; however, it is complex, costly and time-consuming compared to measuring the diameter at breast height (DBH) (Corral-Rivas et al., 2019). DBH is an important component of growth models because it correlates with TH and allows simulating the survival and growth of individual trees in a forest stand (Bronisz & Mehtätalo, 2020); by doing so reliable and easily measurable results are obtained and the standard error is reduced.
On the other hand, studying the growth dynamics of a forest species based on DBH and TH allows the construction of height-diameter (HD) models which predict TH of missing trees and yield estimates, dominant height, competition indices, and carbon storage potential (Souza et al., 2023; Gorgoso-Varela et al., 2019; Mugasha et al., 2019). These procedures offer flexible perspectives to understand specific differences between species or functional groups. As a consequence, silvicultural research reduces inventory costs and facilitates dasometric records during a determined period of time (Liu et al., 2023). The development of allometric equations specific to each species, site and plantation system is essential to obtain accurate and reliable estimates that contribute to the productive and sustainable development of forest plantations (Lucero et al., 2018).
In this regard, in the high Colombian Andean zone, one of the native forest species of economic importance is Aliso (Alnus acuminata Kunth). It is characterized by its rapid growth, light wood, assistance in recovery of degraded areas fixation of atmospheric CO2 and nitrogen (N) to the soil; overall, these features make it a suitable species in agroforestry (Silva et al., 2018; Tafur, 2021). This species, native to dry mountainous areas of Latin America, extend from the Mexican northwest to the Argentine north, in latitudes between 20° north and 20° south, with an altitudinal range from 2400 to 3200 meters above sea level; in Colombia, it is found in the Andean region in the Central and Eastern Cordilleras (Paniagua-Zambrana et al., 2020).
A. acuminata is characterized by being tolerant of infertile soils due to its ability to form ectomycorrhizal relationships with actinomycetes of the genus Frankia (Saucedo et al., 2020), with which A. acuminata develops symbiosis and is responsible for nitrogen (N) fixation and phosphorus (P) absorption, which represents an advantage for the growth of associated plant species (Silva et al., 2018; Vásquez et al., 2020).
However, despite the economic benefits and ecosystem services it generates, little research exists on allometric equations that allow establishing the development of A. acuminata in the high Andean region; therefore, it is necessary to conduct studies regarding its growth rate under the natural conditions of the study area. This species shows rapid growth due to its good productive development, which represents an alternative to the establishment of forest plantations and agroforestry systems.
Considering the above and the need to increase the information in repositories on the characterization of native forest species, The objective of this study was to develop and validate a mathematical model to predict the total height of Alnus acuminata Kunth based on diameter, using an allometric equation of mixed effects models for the high tropics of Nariño.
The information generated is expected to provide a tool that facilitates quick and efficient height estimation for the management and planning of A. acuminata plantations.
MATERIALS AND METHODS
Location and description of the study site. This research was carried out in the forest area of the Obonuco Research Center of the Colombian Corporation for Agricultural Research - AGROSAVIA, in San Juan de Pasto, Colombia, at 3230masl with coordinates LN 01°11'4.13', LW -77°19'0.19', in the moorland where Galeras Flora and Fauna Sanctuary is located. According to the IDEAM-Instituto de Hidrología, Meteorología y Estudios Ambientales (2020), the average annual rainfall is 796mm, with an average temperature of 12.8°C, sunshine between 2.5 and 3 hours a day and a relative air humidity of 74%.
According to Méndez (2004) and Lince-Salazar & Sadeghian (2021), the study area presents soils of the Andisol type, characterized by being deep, well-drained, and located in Vitric Haplusstands land (AMBa, AMBb and AMBc). Table 1 presents the most relevant information regarding the physicochemical characteristics.
Table 1 Physicochemical analysis of the soil of the A. acuminata plot of Obonuco.
| Texture | pH | MO (%) | P (mg/kg) | Ca | Mg (mEq/100g) | K | ECEC | B | Cu | Mn (ppm) | Fe | Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sandy loam | 5.7 | 10.2 | 4.87 | 4.5 | 1.0 | 0.4 | 6.2 | 0.1 | 3.7 | 5.2 | 335.9 | 2.1 |
Dasometric variables. Dasometric data were obtained from samplings carried out during 75 months (September 2014 to December 2020) on 200 contemporary A. acuminata trees; these were established in permanent sampling plots in an area of 1800m2 and planted in a 3 x 3m square. Half-yearly measurements were made to the entire plantation, in which TH (m) information was recorded with a precision clinometer (Suunto Tandem 360PC/360R); DBH (cm) at 1.30m from the soil with diameter tape, age (in months from establishment) and survival (%) (Manzanilla et al., 2022).
With respect to the dasometric mean annual increase (MAI), it was calculated by means of the relationship of the average value of the last measurement made to the variables DBH and TH in the plot and age from the time of establishment. The result represents the average total growth at a certain age and the annual measurement of growth for any age (Vasquez-Miñope et al., 2023) ( Equation 1).
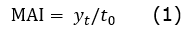
MAI= mean annual increase
yt= dimension of the variable (DBH, TH)
t0= planting age
Allometric models. In order to determine the relationship between the TH-DBH variables and determine the total height, four models were used in forestry sciences; these were evaluated based on the analysis of linear, modified linear and polynomial regression models using restricted maximum likelihood (REML) (Table 2). For the selection of the best model, the Akaike (AIC) and Bayesian (BIC) evaluation criteria were applied. The information was analyzed through the "Nonlinear Mixed-Effects Models" package of the R software (Bates et al., 2015).
Table 2 Allometric models evaluated to estimate TH of A. acuminata based on DBH and age in Nariño, Colombia.
| Model | Type | Equation |
|---|---|---|
| 1 | Linear | H=β0 + β1*DBH |
| 2 | Polynomial | TH=β0+β1*DBH+β2*DBH² |
| 3 | Mixed linear (random effect per tree) | TH=β0 + β1*DBH + α*id. Tree |
| 4 | Mixed linear (random effect per tree age) | TH=β0 + β1*DBH+Ζ*age. Tree |
β0, β1, β2, α and Ζ are parameters of the models: DBH is the diameter of the tree at breast height in centimeters - measured at 1.3 m, age of the tree in months, id is tree identifier and TH is height measured in meters from the base to the apex.
For the selection of the model, it was taken into account that it should be a good fit to the observed data, i.e. have a high coefficient of determination (R2), a low standard error of estimation (SEE) and a low estimation bias (EAR and EAM). Comply with the mathematical assumptions of regression, such as normality, homoscedasticity and independence of the residuals. Also, be parsimonious and easy to interpret, having few parameters and a simple functional form.
RESULTS AND DISCUSSION
Comparing the development of forest species established in different environmental conditions allows evaluating the productive and ecological potential. However, it is unlikely to find studies with exact environments to those implicit in each study; therefore, the results are compared with other studies under similar conditions. In this sense, in the evaluation of Alnus acuminata Kunth, differences in the dasometry can be observed. These differences may reflect the adaptation of the species to environmental conditions, such as climate, soil, topography, and hydrology, as well as the influence of forest management practices. (Hernández-Ramos et al., 2022).
Dasometric variables and mean annual increase. Alnus acuminata at 75 months after establishment, presented 81% survival and registered a TH between 4 and 13m with an average mean of 7.40 ± 1.30m. Regarding DBH, it ranged from 4.25 to 16 cm, with an average of 9.51 ± 2.23cm. As for the mean annual increase in height (MAIH), the record was 1.18 m∙year-1 and in relation to the mean annual increase in diameter (MAID), its value was 1.50 cm year-1.
The results indicate that the dasometric variables of TH and DBH were lower than those reported by other studies. Authors such as Ospina et al. (2005) claim that in studies conducted in Colombia in the departments of Huila, Risaralda and Tolima, A. acuminata at 60 months of age presented a TH of 13.67, 13.30 and 11.87m, while the values of DBH were 12.70, 13.40 and 12.37cm for each location respectively. Similarly, Aulestia et al. (2018) in Ecuador, found that at 132 months of age, A. acuminata registered a TH of 15.90m and a DBH of 31cm year-1. By the same token, Cyamweshi et al. (2021) in Rwanda, in the Kabumba, Mutovu and Rusiza regions, A. acuminata plantations of 84, 58 and 54 months of age reported TH values of 9.90, 7.60, and 8.30m and DBH of 22.20, 18.00, and 18.50cm in each area.
In the same vein, the MAID and MAIH values obtained in the previously mentioned investigations were higher than those reached in this investigation (MAIH 1.18 m year-1 and MAID 1.5cm year-1). Ospina et al. (2005), in the departments of Huila, Risaralda and Tolima (Colombia), reported the MAIH for A. acuminata of 2.73, 2.66 and 2.36 year-1 and MAID of 2.54, 2.58 and 2.45cm year-1 respectively. In the high Andean region of Nariño, 2800masl, Escobar-Pachajoa et al. (2019) found that A. acuminata registered a MAIH of 2.41m year-1 and MAID of 2.6cm. Similarly, Aulestia et al. (2018), in Ecuador, established that A. acuminata registered a MAIH of 1.45 m year-1 and IMAD of 2.81cm year-1. Cyamweshi et al. (2021), in the regions of Kabumba, Mutovu and Rusiza, reported a MAIH of 1.41, 1.84 and 1.61m year-1 and IMAD of 3.17 and 3.96cm year-1.
The contrast of the results obtained in relation to other studies is probably due to the negative effect of the altitude on the growth of the species since the forest plot is established at 3230masl, which can alter the variables TH, DBH and the MAI. In this regard, Escobar-Pachajoa et al. (2019) mention that, in the evaluation of tree components for the high Andean region, it was found that altitude influenced data such as DBH and TH. Other authors indicate that the growth rates of forest species possibly decrease due to limitation of nutrients, low microbial activity of the soil and sunlight, reduced transpiration, and strong winds (Sandoya et al., 2021). Other factors may include low ambient and soil temperatures, which are common in higher-altitude regions (Hernández-Ramos et al., 2019) In line with these findings, Cyamweshi et al. (2021) obtained similar results in Rwanda, where the development of A. acuminata was higher at intermediate altitudes and lower at higher elevations.
With respect to the above, Ensslin et al. (2015) mention that studying the growth of trees in a certain region with climatic parameters and particular soils and comparing the results obtained with those reported for the same species in other regions could yield information on the variation of growth rates of the species. Altitude, soil and climate can influence forest development and vigor. Regarding the altitudinal gradient, it is possible that it is one of the factors with the greatest incidence in plant development, considering that it influences the temperature and humidity parameters of the soil and environment; in the plant, this is evident in the reduction in the typical MAIs of each species and the decrease in the vigor of the vegetation (Romahn-Hernández et al., 2020).
Although the A. acuminata trees studied present dasometric values lower than the ranges reported by other authors, their survival percentage, greater than 80%, indicates that the species, despite being outside the registered altitude range (2400-3200masl), adapted to the soils and climatic parameters of the moorland where the Galeras Flora and Fauna Sanctuary is located. Its tripartite association and the ability to fix N perhaps influenced this adaptation, and so did the climatic lag between the altitudinal ranges and the vegetation cover caused by global warming that can affect the dynamics, distribution, health, adaptability, and sustainability of the various organisms and ecosystems (Romahn-Hernández et al., 2020).
On the other hand, if the development of A. acuminata is compared with other forest species of economic importance in the high Andean zone, it is seen that growth is relatively good. According to Mendoza et al. (2019), it is believed that A. acuminata has a higher growth compared to the MAIH of Pinus patula which is 0.8 m year-1 and that of Cedrela montana which reaches a MAIH of 0.03m year-1 and a MAID of 0.06cm year-1. In relation to this, Silva et al. (2018) indicates that the growth of A. acuminata is due to the ability to fix atmospheric N, which improves its vegetative development. These values were only exceeded by the Eucalyptus species, which, according to Oliveira et al. (2015) registered a MAID of 4cm and MAIH of 4.5m. However, its implementation and management must be carried out with special technical attention since it is an introduced species, like Pinus patula; therefore, it can have negative effects on biodiversity and the physical-chemical conditions of the soil (Aulestia et al., 2018).
Allometric models. Allometric models were generated; these were related to the growth of the TH variable of the tree with easily measured variables such as DBH and age. These models were tested to determine TH as a function of tree individuality, DBH and age (months). The best equation was selected taking into account the assignment of greater weight to the fit criteria of the model; the lower the AIC and BIC values, the better the equation, a condition that is met in M4 (Table 3).
Table 3 Allometric growth models in total height for Alnus acuminata.
| Model | Coeficient | Selection criteria | ||||||||
| β0 | β1 | β2 | α | Ζ | r² | r². adj. | Sigma | AIC | BIC | |
| M1 | 045 ± 0.06 | 0.65 ± 0.01 | 0.72 | 0.71 | 0.96 | 5575.47 | 5592.3 | |||
| M2 | 0.01 ± 0.12 | 0.81 ± 0.04 | 0.01 ± 0 | 0.72 | 0.72 | 0.95 | 5560.9 | 5583.4 | ||
| M3 | 0.08 ± 0.06 | 0.74 ± 0 | 0.7 | 0.75 | 4759.8 | 4782.3 | ||||
| M4 | 2.07 | 0.35 | 0.95 | 0.86 | 4338.5 | 4360.9 | ||||
M1: linear model: M2: polynomial model; M3: mixed linear (random effect per tree); M4: mixed linear (random effect per tree age)
Based on the results obtained from M4, the following equation was generated, Equation 2:
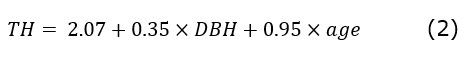
Where,
TH= Total height A acuminata
DBH= Diameter at breast height
Age= Months since establishment
Mixed effects models, in which fixed and random effects stand out, present a well-justified solution for this type of forest data (Ogana, 2022) The models were adjusted by referencing DBH as an independent variable and age as a correction factor given their easy, fast and low-cost measurement; in addition, they account for a high percentage of the total variability (Revilla-Chávez et al., 2021; Sáenz et al., 2021). In this sense, taking into account the results of this research, to estimate the TH of A. acuminata under agroecological conditions of the high Andean zone and during the first vegetative period of the species (up to 75 months of age), it is only necessary to have information on the approximate age and DBH of the tree being evaluated. The use of this type of model allows for flexible prediction of modeled parameters. In cases where measurements (variables) are missing, only fixed effects can be used, but additional measurements allow predicting random effects, considering different levels of clustering (Bronisz & Mehtätalo, 2020).
Unlike previous studies such as that of Bacca-Acosta et al. (2021) and Barbosa et al. (2019) on growth modeling in tree species, non-linear allometric models are not always the best at predicting variables such as TH and DBH. The study carried out by Chenge (2021) indicates that linear models are the most common methods to establish allometric relationships between two variables. In the same way, in the present investigation, in Table 3, it can be observed that the mixed linear and the polynomial models yield good results; when looking at criteria like BIC and AIC, the M4 is the best fit. Of the models evaluated, the mixed linear (M2) and polynomial (M4) models show good results in terms of coefficient of determination (R2) and root mean square error (RMSE), indicating that they fit the observed data well. However, when contemplating the Bayes information criteria (BIC) and Akaike's information criteria (AIC), which penalize the complexity of the models, it can be seen that M4 is the best fit, since it presents the lowest values of BIC and AIC. This suggests that M4 is the most parsimonious and efficient model to explain the relationship between the variables.
In Figure 1, for each data group per age, there is a line in which the variation of the slope is represented over time. This demonstrates a random effect of the age of the tree when correlated with the variables TH and DBH.
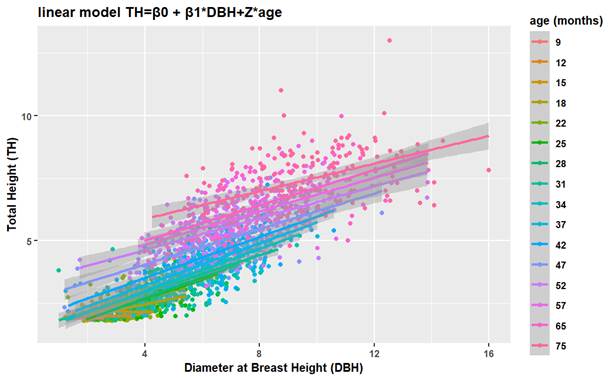
Likewise, in Figure 1, the use of the mixed linear model (M4) can be observed using the evaluation periods or age in months; the trend lines are more prominent. For each data group per age, there is a line where the variation of the slope over time is observed. This shows that there is a random effect due to the age of the tree when the variables TH and DBH are correlated; that is to say, they present a greater degree of inclination during the first months of development of the species, especially during months 12 and 42. Between 52 and 75 months of development, the slope of the trend line decreases. This behavior may be explained by the fact that initially, the forest species present a rapid development with respect to the height variable until they reach the degree of maturity in which the height stabilizes and only the DBH reaches a higher value up to a certain point (Parra-Piedra et al., 2017). In addition, the inclusion of random effects allows for modeling the natural variability in the HD relationship between different locations and time, which provides better estimates and avoids bias (Gorgoso-Varela et al., 2019).
The information presented for this development period allows us to know what the growth dynamics of the species is like and to obtain a reliable equation to determine the height from DBH. If it is necessary to learn more details about the moment when this variable stabilizes, it is important to continue recording dasometric variables in order to obtain an equation for all vegetative development. This information will be relevant to establish adequate silvicultural management, such as cutting cycles or optimal use (Guera et al., 2019).
Comparison of forest allometric models under different environmental conditions is a useful method for assessing the adaptation and performance of forest species. By comparing models, differences in the relationship between tree size and shape can be detected. These differences may reflect the effects of environmental factors, such as temperature, precipitation, radiation, wind and humidity, on tree growth and structure. Comparison of forest allometric models can also be used to select the most suitable species for each environmental condition and to optimize the use of forest resources (Cyamweshi et al., 2021; Escobar-Pachajoa et al., 2019).
CONCLUSIONS
The altitudinal factor and climatic parameters were decisive in the development of the species Alnus acuminata in the high Andean region of the department of Nariño. This was evidenced in the dasometric and mean annual increase in height variables presented by the species, which were lower than those reported in other areas with lower or intermediate altitudes.
The results of this research suggest that when applying the selection criteria and evaluating the type of adjustment presented by model 4; 𝑇𝐻= 2.07+0.35×𝐷𝐵𝐻+0.95×𝑎𝑔𝑒 this turned out to be the best fit to estimate the height of A. acuminata in a simple and efficient way. The model can be applied in agroecological zones similar to the study site and for trees with ages of up to 75 months.

