











 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
INTRODUCTION
Rice (Oryza sativa L.) is one of the main crops in Brazil. According to CONAB, the Brazilian grain harvest could reach 10.055 million tons. Production in Tocantins is estimated at 636.2 tons in an area of approximately 111.2 hectares and productivity of 5,721 kg ha-1 (CONAB, 2021). Thus, for the culture to achieve high productivity, it is essential to use high-quality seeds. Therefore, seed treatment is an important practice, reducing infection and infestation of pathogens, mainly those that survive in the soil (Henning, 2005).
The microbiolization technique has been used with satisfactory results and presented as a viable strategy to reduce the exclusive use of synthetic products, providing economic and environmental benefits (Junges et al., 2014). The physiological quality of seeds, represented by viability and vigor, can directly influence many aspects of their performance, such as, for example, the emergence rate and total emergence (Pádua et al., 2010). High-vigor seeds present great speed in the metabolic process, providing faster and uniform emission of the primary root in the germination process and a higher growth rate, producing seedlings with greater initial size.
The genus Bacillus is composed of aerobic and gram-positive bacteria that form endospores resistant to adverse environmental conditions such as heat and desiccation, presenting a high potential for biocontrol of plant parasite fungi and acting as growth promoters (Madigan et al., 2016). The first step in biological control is the selection of microorganisms from the environment, such as soil and plants, and then the verification of their antagonistic effect, identification of the mechanisms of action present, and the effects of combination with other strains, seeking a broader spectrum of action of microorganisms against the pathogen (Timms-Wilson et al., 2000; Huang et al., 2004; Kilic-Ekici & Yuen, 2004).
Therefore, the search for the management of plant parasite in a sustainable way is fundamental to the control of plant disease. Thus, the objective was to evaluate the phytosanitary protective effect and physiological quality of irrigated rice seeds inoculated with Bacillus sp. isolates in order to minimize damage caused by plant parasite fungi.
MATERIAL AND METHODS
Concentration of Bacillus sp. to seed treatment. The tests were developed in the Laboratory for the Improvement of Postgraduate Studies in Plant Production, UFT, Gurupi - TO Campus. Six isolates chosen according to the adapted methodology of Praça et al. (2009) were evaluated (Table 1). Each bacterial isolate was cultured in an Erlenmeyer flask with a capacity of 250mL, containing a medium with sterile nutrient broth, and incubated at 30℃ for 36 hours in a shaker-type reactor. Subsequently, the isolated were used for seed treatment, inoculating 1mL of bacteria in each Gerbox at a proportion of 2.5 times the mass of the autoclaved dry paper (MAPA, 2009).
Table 1 Isolated description utilized in this study.
| Isolated | Specie | UFC.mL-1 |
|---|---|---|
| 1 | Bacillus sp. | 2.32 x 1011 |
| 2 | Bacillus sp. | 2.3 x 104 |
| 3 | Bacillus sp. | 1.3 x 104 |
| BSRW | Bacillus sp. | 5.6 x 109 |
| 47b | Bacillus sp. | 2.48 x 1017 |
| UFT 38 | Bacillus thuringiensis | 6.8 x 109 |
Germination test, germination speed index (GSI) and fresh mass weight. For the germination test, six replicates of 25 rice seeds of the cultivar BRS A701 CL produced in the Dois Rios farm in the municipality of Lagoa da Confusão, Tocantins, were used (10°47' S, 49°37’ W, altitude 186 m). The seeds were previously treated with the bacteria: isolated 01, isolated 02, isolated 03, BSRW, 47b, and UFT38, and the control treatment (where there was no inoculation with the bacteria). The seeds were distributed evenly over the substrate of two sheets of paper and placed in a sterilized Gerbox moistened with 50ml of distilled water. The temperature in the germination test was 25℃.
The evaluation of final germination occurred at the end of the seventh day, and the germination speed index (GSI) was evaluated daily until the seventh day (Kotowski, 1926). In the end, the characteristics of shoot fresh mass (g), root fresh mass (g), and seedling size (cm) were evaluated. Germination speed index: GSI = (G1+G2+G3+...Gi)/(G1T1+G2T2+G3T3+…GiTi) x 100; where: G1 to Gi is the number of seedlings germinated per day; T1 to Ti is the time in days. After significant results for isolate 47b in the germination test and GSI test, the isolate was selected for subsequent tests: indoleacetic acid test (IAA), production of extracellular enzymes, antagonism test and phytosanitary analysis of seeds.
Indoleacetic acid test (IAA). Based on the results of previous tests and the finding of the superiority of isolate 47b, it is assumed that the high germination rate could be related to the presence of growth hormones, such as indoleacetic acid. Thus, it was isolated in Luria Bertani (LB) liquid medium with the addition of L-Tryptophan (1g L-1) precursor of IAA, covered with aluminum foil to prevent light, and taken to the shaker under agitation at 200 rpm for 72 hours using the calorimetric method explained by Gordon & Weber (1951).
Then, in Eppendorf tubes at 1000 rpm, the bacterial culture was centrifuged for 5 minutes. After centrifugation, 500 µL of the supernatant was pipetted with the addition of 1mL of Salkowski’s reagent (2% 0.5 M FeCl3 in 35% perchloric acid). Subsequently, the Eppendorfs were covered with aluminum foil for about 30 minutes, and then the absorbance reading was performed using a spectrophotometer (530 nm). The equation 1 of the straight line obtained from the standard curve was:

Production of extracellular enzymes. The test for cellulase production was performed with isolated 47b in liquid CCY medium already fermented with the pH adjusted to 4, 6 and 8. The microdorp technique was carried out in CMC medium - carboxymethylcellulose for 48 hours at 30℃. After two days of incubation, the plates were submerged with 10 ul of a 0.2% congo red solution for 5 minutes and dissolved with 1M NaCl for 15 minutes. The light color indicated the presence of cellulase (Table 2).
Table 2 Classification of the production of extracellular enzymes byy isolate 47b at different pH (4, 6 and 8).
| Results | Halo |
|---|---|
| Strong (++) | Vary evident halo |
| Positive (+) | Absence of halo |
| Negative (-) | without absence of halo |
Antagonism test against plant parasite fungi. The antagonism test was performed with isolated 47b, being the best and most responsive in the germination test and previous works in the Molecular Biology Lab. Isolate 47b was grown in the described culture medium at pH 4, 6, and 8. The fungi used were Colletotrichum sp., Curvularia sp., and plant parasitic fungi by bacteria were evaluated according to Matić et al. (2014), in which a streak of the bacteria was paired with a disk (8 mm) in a Petri dish (90 x 15 mm) in PDA culture medium (Kasvi).
For the double culture assay, a loop of the bacteria was strained 30mm from the edge of the plate, and the fungus disk was placed centered 26 mm from the opposite edge and 26 mm from the striae of the bacteria, in triplicate for each double culture. The plates were incubated at 25℃ being evaluated at 2, 4, 7 and 15 days. Mycelial growth was measured daily with the aid of a caliper, and the inhibition of fungal growth was calculated by the percentage of inhibition of radial growth (PIRG)according to the formula: PIRG = ((R1 - R2)/R1) x 100, where R1 corresponds to the radial growth of the plaque and R2 to the radial growth of the fungus towards the antagonist (Fokkema, 1976).
Phytosanitary analysis of seeds. In the phytosanitary analysis, rice seeds of the cultivar BRS A701 CL were used, inoculated with the bacterium 47b, pre-selected in the previous tests under pH4 and pH6 conditions, which presented the best results in the antagonism test.
The seeds were evenly distributed in the pepper substrate in six replications, placed in a Gerbox disinfected with 70% alcohol, containing two sheets of sterile filter paper, and moistened with 50 mL of sterile distilled water, using 25 seeds per treatment, evaluating on the 7th and 25th days the emergence of plant parasites. The survey of the fungal population associated with rice seeds was carried out using the filter paper method or blotter test (MAPA, 2009).
The seeds from each group were submitted to asepsis. Disinfestations were carried out by immersing the seeds in a sequence of 50% alcohol solution for 30 seconds, 2% sodium hypochlorite for 2 minutes, and then two sequences of washing in sterilized distilled water. The seeds were placed individually and, later, placed in an incubation room at a temperature of 25℃ and a photoperiod of 12 hours for 24 hours. Subsequently, the Gerboxes were placed again in an incubation chamber for five days with a 12-hour photoperiod, where they remained until evaluation (MAPA, 2009).
The sanitary analysis of the seeds was carried out individually in each Gerbox, with the aid of a stereoscopic and optical microscope. To identify the fungi, slides were prepared and visualized under an optical microscope, recording the incidence (%) of all genera found. Specialized literature, such as Barnett & Hunter (1998) and Watanabe (2010), was used in the identification.
Statistical analysis. For the variables germination, GSI, root fresh weight, shoot fresh weight, and seedling size, the treatments consist of the evaluating the six isolates in addition to the control treatment (without inoculation) in randomized blocks with six repetitions. The determination of AIA and the cellulase at three pHs (4, 6 and 8) were carried out on a single isolate (47b) pre-selected in previous tests. For the antagonism tests against plat parasite fungi, the treatments consisted of a factorial arrangement (4x4), with the first factor being isolate 47b in three pHs (4, 6 and 8), plus the control treatment, and the second factor determinated by the four evaluation times (2, 4, 7, and 15 days), also carried out in randomized blocks with three repetitions. For the phytosanitary analysis of the seeds, the treatments consisted of a factorial arrangement (3x2), the first factor being the isolate 47b at pH 4 and 6, plus a control treatment, and the second being the evaluation times (7 and 25 days). The data were subjected to analysis of variance using the F test. To compare means, the Tukey test at 5% probability was used for the qualitative factor (pH), as well as regression analysis for the quantitative variable (assessment periods).
RESULTS AND DISCUSION
In the analysis of variance (P≤0.05), there was no significance for the seed treatment factor in the germination; however, a difference was detected for the germination speed index (GSI). In the germination of rice seeds (Figure 1A), there is a similarity between the tested isolates as well as the control treatment. The growth velocity index (GVI) (Figure 1B) had a germination superiority of 4, although it did not differ from the isolated BSRW, showing growth promotion potential. Germination potential was also verified for isolates 3, UFT 38, and BSRW when compared, emphasizing the importance of isolating new regional strains to promote better performance between the isolate and the cultural and local bioprospecting.
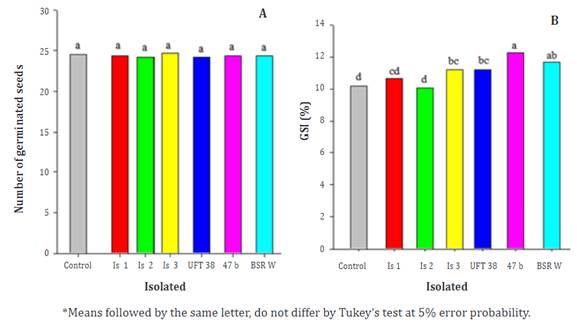
Figure 1 Germination (A) and germination speed index (GSI) (B) for the cultivar BRS A701 CL evaluated during seven days submitted to different seed treatments.
Comparing the GSI of isolated 47b that showed better growth promotion with the control treatment, the positive effect of Bacillus allowed better initial seedling development. The accelerated germination resulting from the growth promotion of the microorganism can anticipate the emergence of seedlings, resulting in an increase in photosynthesis and, consequently, an increase in productivity, in addition to avoiding a lower potential for the emergence of possible pathogens in the soil and seeds. Several studies have described bacteria of the genus Bacillus as plant growth promoters due to the production of phytohormones, solubilization of phosphates, production of siderophores, and inhibition of ethylene synthesis.
According to Soares et al. (2012), the treatment of rice seeds with Bacillus increases the physiological potential of low-quality rice seeds, with positive effects on the percentage of germination, first germination count, and emergence at 14 days. It is possible to positively evaluate the benefits of the use of biological products in several cultures to promote growth, high germination rate, and control of plant parasites and insects. The use of microbiolization techniques, like others used with microorganisms, aims to reduce the use of chemical agents with less environmental impact and production costs.
As for the fresh mass weight, there was significance for the seed treatment factor in the aerial part, root, and seedling size evaluations (P≤0,05). It is noted (Figure 2) that isolate 47b stood out for being the only one to differ statistically from the control treatment and UFT 38, corroborating the data on germination speed index, where isolate 47b also showed superiority to most treatments. It is understood that, with the faster germination of the seed, the seedling may present more aerial parts due to the synthesis of growth hormones such as IAA, produced by Bacillus, characterizing it as a microorganism that promotes plant growth.

Figure 2 Weight for aerial part and root for the cultivar BRS A701 CL subjected to different treatments at the end of the 7th day of evaluation.
For the root fresh weight (Figure 2), the superiority of isolate 47b was again observed; it was the only one to differ from the control treatment. The greater rooting can be explained by the production of IAA stimulant by the inoculation of the bacteria and through the reduction of ethylene levels by the action of enzyme ACC deaminase; thus, the activity of ACC deaminase decreases the production of ethylene in the roots of host plants, resulting in stretching (Prigent-Combaret et al., 2008).
Jain et al. (2016) showed that Bacillus isolates can increase aerial part and root fresh weight in soybean, in addition to increasing the number of lateral roots, thus suggesting that this region may have grown, increasing the ability to obtain available water on the soil. Lazzaretti et al. (2005), using an isolate of B. subtilis in the bean crop, observed that the bacteria introduced an increase in the dry matter weight of the roots and shoots, as well as the presence of a greater number of nodules on the roots of the crop. For seedling size (Figure 3), isolates 47b, UFT 38, and IS 3 were the only ones to differ from the control treatment.
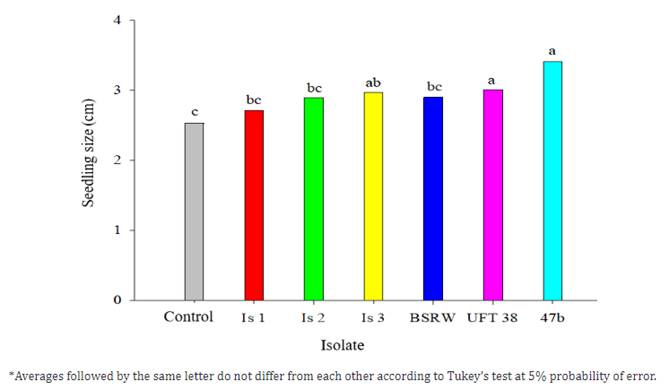
Figure 3 Seedling size (cm) of cultivar BRS A701 CL submitted to different treatments of the 7th day of evaluation.
A larger shoot allows for a better photosynthetic rate, which implies more photo assimilates translocated to growing or reserve organs in the following stages (Taiz & Zeiger, 2004). This increase in shoots may be related to the ability of the bacteria studied to produce indole compounds. Bacillus subtilis promotes plant growth through several mechanisms, which can be direct or indirect, acting together or separately. Among the mechanisms of action are the production of growth-stimulating phytohormones, phosphate solubilization, production of siderophores, and inhibition of ethylene synthesis (Glick et al., 1994). Plant growth and development are often responses to the production of growth regulators produced by different species of plant-associated bacteria (Oliveira et al., 2014).
The in vitro concentration of indoleacetic acid (IAA) for isolated 47b, which showed significant results in the previous test, referring to the others, was 78.69 µg mL-1, with 0.247 absorbance (530 nm). Raddadi et al. (2008) observed IAA production between 1.53 and 9.71 µg mL-1 with strains of B. thuringiensis, confirming that Bt strains can produce IAA in vitro, thus explaining greater growth in seedling size and greater mass weight fresh from the aerial part. Bacillus subtilis can also conduct the hormonal regulation of plants, thus providing control over root growth through the synthesis of auxin, gibberellin, and cytokinin (Tsavkelova et al., 2006).
To produce extracellular enzymes (Table 3), 47b was isolated at pH 8, seen after 48 hours of inoculation (Figure 4), resulting in greater cellulase production, with enzymatic hydrolysis being observed by the formation of a light-colored halo around the colony. The bacterium at pH 6 is also a producer, however, in a smaller amount than the first mentioned, as described by the index in Table 3. At pH 4, there was no halo formation, making it clear that they weakly induced the production of enzymes.
Table 3 Cellulase production of isolated 47b at different pH.
| pH | Results obtained: |
|---|---|
| 4 | Negative (-) |
| 6 | Positive (+) |
| 8 | Strong (++) |

Some Bacillus species secrete cellulase, and B. subtilis and B. cereus can produce cellulase and other compounds (Rocha et al., 2017). It is of greater interest the use of cellulases, but also other enzymes such as pectinases and hemicelluloses in agriculture since they can control plant diseases and improve crop growth (Behera et al., 2016). Among the known benefits of cellulase are the facilitation of seed germination, plant growth, and flowering, as well as the control of plant diseases (Behera et al., 2016), thus evidencing its potential use as a biocontrol to protect plant seeds from soil pathogens that can infect the seed, thus impairing emergence in the field.
For the antagonism test, there was significance (P≤0,05) of isolate 47b at different pHs for all evaluated fungi, Curvularia sp., Colletrotrichum sp., and Macrophomina sp., as well as the interaction pH vs. days of evaluation at the 2nd, 4th, 7th, and 15th days. As for Curvularia sp. (Figure 5A), no difference was observed in fungal growth at different pHs at two days of evaluation. At four days, greater growth was observed for the control treatment, followed by pH 8. At 7 and 15 days of evaluation, there is no marked growth for pH 6, as well as for pH 4 and 8, evidencing the antifungal potential of isolated 47b for the mycelial growth of the fungus.

Figure 5 Antagonism test and regression curve (A) with isolate 47b against the fungus Curvularia sp., antagonism test and regression curve (B) with isolate 47b against Colletotrichum sp. and antagonism test and regression curve (C) with isolate 47b against the fungus Macrophomina sp. evaluating diameter (cm) on the 2nd, 4th, 7th, and 15th days.
It was observed that isolate 47b (Figure 6A) under conditions of pH 4, 6, and 8 over the course of a day inhibits the growth of the pathogen on the plaque, presenting a bio protective effect since, compared to the control, the fungus continues its growth during the passing of days, filling the entire petri dish.

Figure 6 Antagonism test with isolate 47b against the fungus Curvularia sp. (A), the fungus Colletotrichum sp., (B) and the fungus Macrophomina sp., (C) at the end of evaluation (15th day).
It is noticeable that the lower acid pH levels of the environment where the bacteria are found respond in an antagonistic way against the phytopathogenic fungus. This result was already expected, as the strain under study was isolated from acid soils of the Cerrado, evidencing the importance of local bioprospecting in the selection of bacteria. It can be said that isolated 47b has a high degree of adaptation when inoculated both in corrected soils (pH close to 6.0) and in acid soils, making possible the inhibitory capacity of fungi such as Curvularia sp.
As for Colletotrichum sp. (Figure 5B), no difference was observed in fungal growth at different pHs at two and four days of evaluation. At seven days, greater growth is observed for pH 6, followed by the control treatment and pH 8. At 15 days of evaluation, there is a marked growth for pH 8, as well as for the control and pH 6.
The bioprotective effect of isolate 47b against Colletotrichum sp., becomes evident at pH 4, where there was no growth of the fungus between the evaluated days, making it partially inactive. When the isolate was in conditions of pH 6 and pH 8, the fungus continued its mycelial growth, demonstrating less efficiency of phytopathogenic control in these pH conditions.
Figure 6B also demonstrates the positive effect of isolate 47b in an acidic environment (pH 4) for the pathogen Colletotrichum sp., as well as the loss of efficiency as a biocontrol of the same isolate in conditions of pH 6 and 8, where continuity is evident in mycelial growth of the pathogen.
As for Macrophomina sp. (Figure 5C), pH 4 was the only one to differ from the control treatment at two days of evaluation. At four days, greater growth was observed for the control treatment, followed by pH 8. At 7 and 15 days of evaluation, there was an accentuated growth of the fungus for the control treatment, as well as the bioprotective effect of isolate 47b, regardless of pH. In the antagonistic test with Macrophomina sp., no difference in pathogen growth was observed, and pH 4 and 8 demonstrated similarity to each other but still had an inhibitory effect against the fungus. The highest growth was evidenced for the control, in constant mycelial growth, as expected because it did not present the biocontrol treatment of the bacteria.
The protective effect of isolated 47b (Figure 6C) occurs because the bacteria produce antibiotics that are directly linked to the control of the phytopathogen. It is important to point out, when comparing the test with the other fungi, that pH control against the pathogen over the course of days may be closely linked to the high degree of mycelial resistance, attack, and damage by the pathogen to the culture, since for Macrophomina sp. the best inhibition was at pH 6.
Bacillus can produce metabolites and antibiotics such as production of siderophores, enzymes, and volatile organic compounds such as iturin and surfactin that have antifungal activity against different phytopathogenic fungi (Patel & Minocheherhomji, 2018). Studies carried out by Wu et al. (2021) indicate that the use of Bacillus sp. species has potential for controlling Alternaria sp., where the main mechanism adopted by the bacteria would be antibiosis. The metabolites produced by Bacillus are considered thermally stable, having activity in a wide range of pHs (Kurata et al., 2017), and have the function of hindering or preventing the development of pathogens on the leaf surface of plants, making their action efficient and satisfactory as a biocontrol agent (Hashem et al., 2019).
Studies have demonstrated that different biological activities are produced by Bacillus metabolites or enzymes, including antifungals (Gu et al., 2017). Two Bacillus strains (velenzis e subtilis) showed antifungal activity against Curvalaria lunata at 36% inhibition, even though it is considered a weak pathogen that does not cause damage to seeds (Xie et al., 2020). Emayavarman et al. (2019) reported that B. subtilis provided an inhibitory potential in vitro test against Macrophomina phaseolina of 31.47% and with Bacillus amyloliquefaciens an inhibited potential of 48.14%. It should be noted that Macrophomina sp. is one of the causative agents in the reduction in plant stand in the crop.
In the phytosanitary evaluation of rice seeds for the cultivar BRS A701 CL, there was a significant difference (P≤0.05) for the seed treatment factor as well as for the day factor in the evaluation. For the interaction between treatment and day (7 and 25 days), there was only significance (P≤0.01) when evaluating the fungus Fusarium sp. As for fungal incidence (Figure 7A), no difference was observed in fungal growth at pH 4 for all evaluated fungi on the seventh day of evaluation. For pH 6, there was a higher incidence of Aspergillus sp. in the control treatment, a higher incidence of Aspergillus sp., and lower incidence of Fusarium sp.

Figure 7 Incidence of fungi with different pH treatments of isolate 47b in rice seeds for cultivar BRS A701 CL evaluated at eh 7th (Figure 7A) and 25th (Figure 7B) days.
At 25 days (Figure 7B) of evaluation, as well as at seven days, no difference was observed in fungal growth at pH 4 for all evaluated fungi. For pH 6, similarly to seven days, there was a higher incidence difference for Aspergillus sp., In the control treatment, again, there was a higher incidence of Aspergillus sp. and lower incidence of Fusarium. It is important to emphasize that the incidence of the fungus over the days can be explained by factors such as humidity, temperature, and storage. Thus, it is noted that, even after the evaluated days, Bacillus continues to present its bioprotective effect on the seed.
Schawrtz et al. (2013) demonstrated that Bacillus simplex 30N-5 inhibits the growth of two members of the complex, Fusarium oxysporumandFusarium solani. It is important to point out that the fungi Penicillium sp. and Aspergillus sp. are storage fungi, which may explain the high occurrence in the phytosanitary test. According to Puzzi (2000), the seed can present a high incidence of Aspergillus sp. and Penicillium sp. due to the proliferation more frequently in stored grains when subjected to relative humidity above 65 or 70%. Soares et al. (2007) evaluated eight Bacillus sp. isolates, where they were able to produce toxic compounds that inhibit mycelial growth for the pathogen Curvularia lunata.
CONCLUSIONS
Isolate 47b demonstrated superiority when compared to the other isolates, showing efficacy in the tests of rice seed germination, growth velocity index, root fresh mass weight, seedling growth, indoleacetic acid, and cellulase indices. The pH 4, pH 6, and pH 8 showed efficiency as a phytopathogenic control of the fungus Curvalaria sp. in the antagonism test. For the fungus Colletotrichum sp., pH 4 was evidenced as having the best inhibition potential. For Macrophomina sp., isolated 47b at pH 6 showed better control of the phytopathogen. Phytosanitary analysis of rice seeds showed that isolated 47b under conditions of pH 4 and pH 6 promotes a bioprotective effect on the seeds, ensuring a low rate of pathogens in up to 25 days of evaluation.

