










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Entomología
Print version ISSN 0120-0488
Rev. Colomb. Entomol. vol.38 no.2 Bogotá July/Dec. 2012
Aspectos biológicos de Dictyla monotropidia (Hemiptera: Tingidae),en nogal cafetero Cordia alliodora (Boraginaceae)
Biology aspects of Dictyla monotropidia (Hemiptera: Tingidae) in laurel trees Cordia alliodora (Boraginaceae)
HAROL ENRIQUE MARTÍNEZ C.1, CARLOS MARIO OSPINA2, ESTHER CECILIA MONTOYA3, LUIS MIGUEL CONSTANTINO4 y PABLO BENAVIDES MACHADO5
1 Ing. Agrónomo, M. Sc. Universidad Nacional de Colombia, Medellín, Colombia, harolmartinez@hotmail.com
2 Ing. Forestal, estudiante M. Sc. Investigador Científico I, Cenicafé, Chinchiná, Colombia, carlosmario.ospina@cafedecolombia.com
3 Estadístico, M. Sc. Investigador Científico III, Cenicafé, Chinchiná, esthercecilia.montoya@cafedecolombia.com
4 Entomólogo, M. Sc. Investigador Científico II, Cenicafé, Chinchiná, luismiguel.constantino@cafedecolombia.com
5 Ing. Agrónomo, Ph. D. Investigador Científico II, Cenicafé, Chinchiná, pablo.benavides@cafedecolombia.com, autor para correspondencia
Recibido: 1-sep-2011 - Aceptado: 2-nov-2012
Resumen: La chinche de encaje, Dictyla monotropidia, es la principal plaga del nogal cafetero Cordia alliodora en Colombia. Al alimentarse del follaje, necrosa y ocasiona la caída prematura de las hojas del árbol. Con el propósito de conocer aspectos de la biología de D. monotropidia, se estudió su fluctuación poblacional y el ciclo de vida. Para esto se estimó mensualmente durante un año, el número de insectos en los árboles y el suelo en una plantación de C. alliodora de 0,8 ha en Manizales. El ciclo biológico del insecto se obtuvo mediante su cría en casa de malla a 22,4 ± 5,5 ºC y HR de 66,8 ± 16,6%. Se presentaron diferencias significativas en el porcentaje de hojas afectadas en los árboles siendo, en promedio, mayor en marzo 2009 (57,4%) y menor en enero 2010 (1,6%). Se encontraron diferencias estadísticas en el número de insectos en los árboles a través del año de evaluación, con un promedio de 1.655 ± 361 individuos en abril y 135 ± 64 en enero. Los individuos en las hojas del suelo oscilaron entre 10,5 ± 2,4 y 245,5 ± 76,8 durante el estudio. La duración promedio del ciclo de vida de D. monotropidia, desde la postura hasta la aparición del adulto fue estimada en 44 ± 0,95 días, siendo de 15,5 ± 0,1 para el estado de huevo, 28,5 ± 0,85 para los cinco estados ninfales y una duración del adulto de 115 ± 2,4 días. El manejo, deberá realizarse en los árboles y en las hojas infestadas del suelo.
Palabras clave: Chinche de encaje. Fluctuación poblacional. Ciclo de vida. Plaga.
Abstract: The lace bug, Dictyla monotropidia, is the main insect-pest of laurel tree Cordia alliodora in Colombia. This insect feeds on leaves causing their dead and making them fall prematurely onto the ground. With the purpose of understanding some aspects of the biology of D. monotropidia, we studied its population dynamics and life cycle. We estimated the total number of insects on the trees and onto the soil, monthly during a year, in a 0.8 ha C. alliodora crop in Manizales. The life cycle of the insect was obtained from insects reared in a screen house at 22.4 ± 5.5 ºC and 66.8 ± 16.6% HR. We observed significant differences in the average percentage of infested leaves in the trees being higher in March 2009 (57.4%) and lower in January 2010 (1.6%). We found statistical differences in the number of insects in the trees along the year, with an average of 1.655 ± 361 individuals in April and 135 ± 64 in January. Individuals onto the ground ranged between 10.5 ± 2.4 and 245.5 ± 76.8 along the year. The average life cycle of D. monotropidia, from egg to adult was estimated in 44 ± 0.95 days, being 15.5 ± 0.1 for eggs, 28.5 ± 0.85 for the five nymph stages and a total duration of the adult of 115 ± 2.4 days. The control of this insect should be performed on the trees and on infested leaves on the ground.
Key words: Lace bug. Population dynamics. Life cycle. Pest species.
Introducción
El nogal cafetero o laurel, Cordia alliodora (Ruiz & Pavon) Oken (1883), es una especie forestal de la familia Boraginaceae que se distribuye desde México hasta el norte de Argentina, incluyendo las islas caribeñas. En América Central es común en las zonas bajas y húmedas, se cultiva generalmente, a menos de 800 msnm, con temperatura promedio superior a 23 °C y precipitación anual entre 2.000 y 5.000 mm. Los árboles son de crecimiento rápido en comparación con otras especies forestales y pueden alcanzar hasta 25 a 30 m de altura y 50 a 80 cm de diámetro; posee fuste cilíndrico y desprovisto de ramas hasta un 60% de la altura total, lo cual ofrece un volumen de madera comercial alto por árbol, con densidades de 1.100 árboles por hectárea se ha obtenido entre 15 y 23 m3/ha/año (CATIE 1991).
En Colombia, el nogal cafetero, es la especie nativa de mayor interés en la actividad reforestadora por la calidad de la madera y su adaptabilidad a diferentes condiciones climáticas y edáficas. Existen aproximadamente 10.104 hectáreas establecidas en cuatro proyectos nacionales: Procuenca en la zona central cafetera (369 has), Programa Forestal del río Magdalena KFW (8.494), Corporación Tolimense de Cuencas Hidrográficas - CORCUENCAS 400 ha, Proyecto USAID- Midas 841 ha (Proexport 2009; Programa KFW 2009; Procuenca en marcha 2007). Igualmente, es una especie ampliamente usada como sombrío en cafetales en el país.
Bajo condiciones de Costa Rica, el nogal cafetero después de fructificar, se defolia totalmente. Este proceso fisiológico se produce dos veces al año y generalmente coincide con los periodos secos de diciembre a febrero y de junio a agosto (Johnson y Morales 1972). También se pueden producir defoliaciones adicionales por la acción de la chinche de encaje del nogal cafetero, Dictyla monotropidia Stål, 1858 (Hemiptera: Tingidae), insecto que causa daño a las hojas, las hace caer prematuramente e impide que cumpla su ciclo fisiológico normal, causando baja floración, fructificación y retraso en el desarrollo del árbol (Arguedas y Fallas 1993). Defoliaciones causadas por la chinche de encaje también se han reportado en Colombia (Madrigal 1987), siendo ésta la principal plaga de C. alliodora en el país. El daño que causa a los árboles lo ocasiona cuando el insecto se alimenta del follaje y al parecer inyecta una toxina que produce necrosamiento, disminución del área de captación de luz y, por tanto, baja la tasa de fotosíntesis. El mayor ataque se observa en las hojas de los estratos bajo y medio del árbol y cuando es muy severo, se ven afectados todos los estratos, lo que ocasiona al final la caída de las hojas. En las plantaciones establecidas en el país se han detectado ataques continuos y frecuentes de D. monotropidia (Madrigal 1987; Fallas et al. 1993; Procuenca en marcha 2007).
Stål (1873) citó la especie D. monotropidia por primera vez para Colombia. Drake y Ruhoff (1965) anotaron como distribución para D. monotropidia: Brasil, Colombia, Guatemala, Panamá, Jamaica, Cuba, Haití, México, Honduras, Costa Rica, Paraguay, Bolivia, Venezuela, Puerto Rico, Argentina, Perú, El Salvador y Trinidad. En estudios realizados por Cenicafé, a lo largo de la zona cafetera nacional, se detectó la presencia de la chinche de encaje desde la Sierra Nevada de Santa Marta hasta Gigante (Huila) (Gómez y Ospina 2003). Como el hospedante es nativo y el insecto también, éste se encuentra en los árboles del género Cordia.
Un estudio realizado por Madrigal (1987), en condiciones de laboratorio de la Universidad Nacional de Colombia- Medellín, ubicado a una altitud de 1.526 m, con una temperatura y humedad relativa promedio de 24 °C y 71%, respectivamente, encontró que la duración total del ciclo de vida de D. monotropidia, desde huevo hasta la emergencia del adulto varió de 28 a 60 días; la etapa inmadura pasa por cinco estadios, cuya duración promedio en días para cada estadio fue de 2,6; 4,1; 4,2; 4,1 y 6,4. La etapa adulta presentó una duración promedio de 32 días. En otro ensayo sobre el ciclo biológico de la chinche de encaje, realizado a nivel de laboratorio en cajas Petri con hojas individuales de C. alliodora se encontró que la duración promedio de la etapa de huevo fue de 9,8 días, los cinco estados ninfales presentaron una duración promedio de 16,1 días, mientras que la del adulto fue de 14,3 días (Fallas et al. 1993).
Las hembras ovipositan en grupos incrustando los huevos en las nervaduras (central y secundaria) por el envés de las hojas. Las ninfas emergidas de cada postura permanecen agrupadas, hasta llegar al estado adulto. La cópula ocurre uno a tres días después, la cual hacen aún en el grupo, aunque a veces la hembra se aleja, seguida de un macho a una hoja cercana o a otra parte de la misma. Una vez fecundada, busca en las cercanías del sitio un lugar adecuado, hace la postura la cual cuida hasta su eclosión, aunque algunas hembras la abandonan un poco antes (Madrigal 1987). Los resultados encontrados en la literatura sobre la duración de los estados biológicos de este insecto son muy variables, lo cual no permite establecer estrategias de manejo basados en la duración del ciclo de vida y el número de generaciones que puede alcanzar en relación con el tiempo.
Siendo el nogal cafetero la especie nativa de mayor aceptación para ser usada en programas de reforestación comercial en Colombia (Procuenca en marcha 2007; Proexport 2009), fue necesario estudiar la chinche de encaje D. monotropidia. En evaluaciones realizadas por Procuenca en marcha (2007), en la cuenca del río Chinchiná (Caldas), se encontraron niveles críticos de la plaga en el follaje, superando los 40 individuos por hoja y provocando defoliaciones sucesivas en periodos secos. Las poblaciones de D. monotropidia sugieren la existencia de varias generaciones superpuestas que dificultan la recuperación del árbol cuando no se realiza algún tipo de manejo. Es poco lo que se conoce sobre la biología del insecto y existen aún dudas respecto al número de generaciones por año bajo las condiciones actuales de la zona cafetera de Colombia. El conocimiento de las condiciones ecológicas y biológicas es fundamental para proponer un plan de manejo del insecto plaga. Por lo tanto, se propuso el presente trabajo de investigación para conocer la fluctuación poblacional durante un año de la chinche de encaje D. monotropidia en una plantación de nogal cafetero en Manizales y determinar el ciclo biológico en plantas de nogal en casa de malla de Cenicafé en Chinchiná.
Materiales y Métodos
Este experimento fue realizado en condiciones de campo de febrero de 2009 a enero de 2010, en la fincaLa Ramada, ubicada en la vereda La Manuela en el municipio de Manizales (Caldas) a una altitud promedio de 1.161 m, con temperatura y humedad relativa promedio anual de 22,5 ºC y 78,8%,* respectivamente y en una casa de malla en Cenicafé “La Granja”, ubicada en el vivero forestal, a una altura de 1.310 msnm con temperatura y humedad relativa promedio durante el experimento de 22,4 ± 5,5 ºC y 66,8 ± 16,6%, respectivamente.
Para cumplir con los objetivos de esta investigación se llevaron a cabo dos actividades: la evaluación del número promedio de individuos de D. monotropidia por árbol a través del tiempo a nivel de campo y la determinación de la duración de los estados biológicos del insecto en casa de malla.
Evaluación del número de individuos de D. monotropidia a través del tiempo en condiciones de campo. El estudio del número de individuos de D. monotropidia a través del tiempo (fluctuación poblacional) se realizó en la finca La Ramada, en un lote experimental de 0,8 hectáreas sembradas con 500 árboles de C. alliodora de cuatro años de edad, a una distancia de siembra de 4 m x 4 m.
En el lote seleccionado se realizó un censo preliminar, y se identificó la cantidad de árboles afectados y no afectados por el insecto y se les llevó el registro de la siguiente información por árbol: número total de ramas, número de ramas con hojas dañadas, las cuales se bajaron con un corta ramas, en cada rama afectada, el número de hojas y la cantidad de éstas lesionadas, número total de individuos en cada hoja con daño, número total de hojas caídas en el suelo por árbol (con radio de 1,5 m a partir del tallo), total de hojas lesionadas en el área del suelo y número total de individuos en las hojas afectadas del suelo por árbol. Las hojas se separaron de las ramas con tijera podadora. Tanto las hojas con chinches de las ramas y del suelo se embolsaron individualmente para contar los insectos por hoja. Se colocó una manta blanca en el suelo de los árboles evaluados, para facilitar el proceso de recolección y selección de las hojas con daño. Con los resultados de esta evaluación se determinó el tamaño de muestra (número de árboles a evaluar mensualmente), con el parámetro promedio de la proporción de hojas lesionadas por árbol, bajo muestreo sistemático, con un error de estimación del 5% de hojas afectadas por árbol y una probabilidad asociada del 95%, el cual fue aplicado mensualmente hasta completar un año de evaluación. Se seleccionó sistemáticamente desde abajo hacia arriba, un máximo de cinco ramas infestadas. De las hojas con lesión, se tomaron al azar el 10% de ellas, de acuerdo con la estimación de la varianza asociada al número de individuos por hoja.
Con la información registrada, se obtuvo las siguientes variables de interés: a) proporción de hojas afectadas por árbol, en la parte aérea y en el suelo, y b) número de individuos en las hojas afectadas por árbol, de la parte aérea y del suelo. Con estas variables se realizó el siguiente análisis estadístico:
– En cada tiempo de muestreo, promedios y variación por árbol.
– En cada tiempo de muestreo, intervalos para el promedio de cada una de las variables de interés con un coeficiente de confianza del 95%.
– En cada tiempo de muestreo, se estimó el total del número de individuos por árbol, en la parte aérea y en el suelo, a nivel del lote.
Se llevaron diariamente, los registros de temperatura, humedad relativa y precipitación de la vereda “La Manuela” en Manizales, con el fin de soportar la discusión de los resultados del análisis con las variables de interés.
Determinación de la duración de los estados de vida de D. monotropidia. Para determinar la duración de los estados de vida de D. monotropidia y disponer de insectos en forma permanente, se crió el insecto en la casa de malla de Cenicafé, bajo las siguientes condiciones; temperatura de 22,4 ± 5,5 ºC y humedad relativa de 66,8 ± 16,6%. Se utilizaron 20 plantas de nogal cafetero con aproximadamente 90 cm de altura en bolsas, de siete meses de edad, provenientes del vivero forestal de Cenicafé, cubiertas con jaulas entomológicas de 100 x 50 x 50 cm. En cada una de estas se colocaron 50 estados biológicos (ninfas de quinto estadio y adultos) de D. monotropidia colectados en un lote de nogal cafetero de la estación experimental “Naranjal” de Cenicafé, ubicado en el municipio de Chinchiná, Caldas, a una temperatura promedio anual de 20,9 ºC y 78% de humedad relativa. Se renovaron las plántulas en el momento que se defoliaron, para el mantenimiento continuo de los estados biológicos de D. monotropidia.
En septiembre de 2009, se inició el ciclo biológico. Para esto se tomaron 300 adultos de D. monotropidia del pie de cría los cuales se trasladaron al laboratorio en una caja Petri plástica de 14,5 cm de diámetro, y se dejaron por un periodo de cópula de 48 horas corroborándose el apareo. Se establecieron diez unidades de muestreo las cuales consistieron en plántulas de 6 meses, con una hoja cubierta con una manga entomológica de 20 x 12 cm, las cuales se infestaron con 10 hembras de D. monotropidia durante dos días, hasta obtener posturas, después de este tiempo se retiraron las hembras. Estas unidades de muestreo se ubicaron en la casa de malla. Diariamente, se contabilizó en cada unidad de muestreo el número de estados biológicos, hasta que se presentó una mortalidad natural del 100%. Con esta información se obtuvo la duración de cada estado de desarrollo (variables de interés), las cuales se determinaron cuando el 70% o más de los individuos de cada estado pasaron al siguiente estado de desarrollo. Los individuos que mudaron se reconocieron por las exuvias que dejaron adheridas a las hojas de nogal.
Con las variables de interés se estimó el promedio por unidad de muestreo, el error estándar y el intervalo para el promedio con un coeficiente de confianza del 95%. Además, se realizó una descripción de cada estado biológico con la ayuda de un estereoscopio.
Resultados y Discusión
Evaluación del número de individuos de D. monotropidia a través del tiempo en condiciones de campo. Con la información del censo preliminar, se estableció el tamaño de muestra para todas las siguientes evaluaciones mensuales basadas en un error de estimación de 5%. Este muestreo consistió en tomar 15 árboles cada mes, seleccionar sistemáticamente desde abajo hacia arriba, un máximo de cinco ramas infestadas, contabilizar el total de hojas y aquellas infestadas, retirar las hojas afectadas y registrar el número total de insectos en el 10% de las hojas de la rama. El muestreo en el suelo se realizó contabilizando el número total de hojas del suelo, aquellas que estaban infestadas, y registrar el número total de insectos presentes en las hojas del suelo por árbol.
En la tabla 1, se puede observar que el porcentaje de árboles con hojas afectadas en el suelo fue mayor que el porcentaje de árboles con hojas infestadas en la parte aérea en algunos meses evaluados, esto sugeriría que las hojas se desprendieron del árbol como consecuencia del daño ocasionado por el insecto.

Entre julio de 2009 a enero de 2010, se presentaron los menores promedios por árbol del porcentaje de hojas infestadas por D. monotropidia, con respecto a los promedios febrero y marzo de 2009, siendo el promedio de marzo el mayor presentado en todo el estudio (Tabla 2). El porcentaje por árbol de hojas del suelo infestadas fue del 100% durante los doce meses de evaluación, razón que puede explicar la caída de las hojas debido al efecto del ataque del insecto en las hojas de la parte aérea de los árboles.
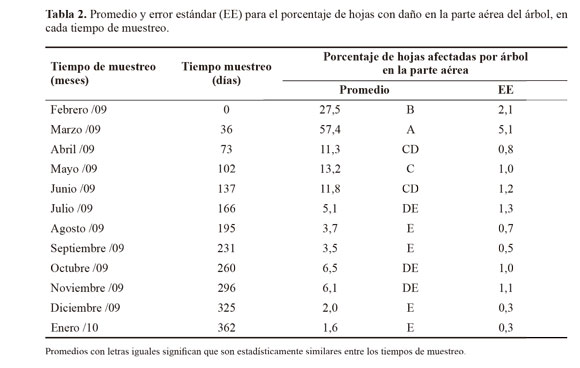
Del día cero al 102 del muestreo (febrero a mayo) y del día 260 a 296 (octubre a noviembre) de 2009, se presentaron los mayores promedios de individuos de D. monotropidia en las hojas afectadas de la parte aérea de los árboles de nogal cafetero (Fig. 1). Dichos promedios, a través de todas las fechas de muestreo, fluctuaron entre 1.654,8 ± 360,7 al día 73 (abril de 2009) y 134,9 ± 63,5 en el día 362 (enero de 2010).
En las hojas de la parte aérea del árbol, no se encontraron diferencias estadísticas significativas en el número de individuos por árbol desde el día cero al 102 del muestreo (febrero a mayo), presentándose en promedio entre 1.519 ± 305,1 y 1.525 ± 345,2 individuos. Del día 137 al 231 (junio a septiembre) los promedios de insectos por árbol se redujeron a una tercera parte (Fig. 1). No se encontraron diferencias estadísticas durante este tiempo. En las hojas del suelo, en promedio por árbol, el número de individuos de D. monotropidia oscilaron entre 10,5 ± 2,4 y 245,5 ± 76,8 durante el año de evaluación, donde no se observaron diferencias estadísticas entre los meses de abril y agosto de 2009 cuando se presentaron las mayores poblaciones del insecto (Tabla 3). Estos resultados indicarían que las altas poblaciones del insecto en las hojas de los árboles durante los primeros meses del año, pudieron haber causado la caída de éstas. Dadas las diferencias en los individuos promedio por árbol del insecto en la parte aérea con respecto a las del suelo, se podría deducir que los insectos se dispersan rápidamente durante la caída de las hojas de los árboles y reinfestan hojas nuevas en los árboles cercanos que están en crecimiento vegetativo.


Dado que el ataque de la chinche de encaje es repetitivo en las plantaciones de nogal, pueden ocurrir defoliaciones sucesivas que retrasarían el crecimiento de los árboles, y en algunos casos, produciría su muerte (Arguedas y Fallas 1993). De acuerdo con lo observado en las plantaciones de C. alliodora de la zona cafetera de Colombia, evidentemente la muerte de los árboles ha venido ocurriendo en plantaciones de menos de tres años de sembradas.
El nogal cafetero después de florecer y fructificar, se defolia totalmente. Este proceso fisiológico se produce dos veces al año que coinciden con los periodos secos de diciembre a febrero y de junio a agosto bajo condiciones de Costa Rica (Johnson y Morales, 1972). Durante este estudio, en las condiciones climáticas y topográficas de la vereda La Manuela, del municipio de Manizales, Colombia, se presentó defoliación antes del tiempo fisiológico normal, a partir de marzo (Fig. 2). Esta defoliación anticipada debió estar relacionada con las altas poblaciones del insecto que la antecedieron, la cual continuó hasta el tiempo normal de caída fisiológica de las hojas al comienzo del tiempo seco, junio (Fig. 3). La segunda defoliación del año, siendo menor, ocurrió durante octubre y noviembre, previo aumento de las poblaciones de D. monotropidia en la parte aérea de los árboles (Figs. 1 y 3).


La baja fluctuación en las lluvias y temperatura (Figs. 3A y 3B) durante los primeros meses del año, en comparación al resto del tiempo de evaluación, además de la disponibilidad de alimento, debió haber influido en el crecimiento poblacional del insecto en los primeros tres meses de evaluación. Según Fallas et al. (1993), los ataques de la chinche de encaje se intensifican durante los períodos secos. De acuerdo con Speight y Wylie (2001), cualquier cambio en la temperatura del ambiente, tendrá un efecto en la tasa metabólica y de crecimiento del insecto, así como un cambio en la habilidad para dispersarse y reproducirse.
Después de la caída de las hojas al suelo la mayoría de los estados ninfales pudieron ser depredados, principalmente por insectos de las familias Chrysopidae, Reduviidae y Formicidae, arácnidos o morir a causa del hongo entomopatógeno Beauveria bassiana, todos observados durante este estudio; sin embargo, los adultos de D. monotropidia colonizaron, caminando y volando, las hojas que estaban rebrotando, principalmente en las ramas bajas de los árboles adyacentes, donde el daño por alimentación y oviposición fue evidente (Fig. 4A). A medida que aumentó el número de individuos, las ninfas de estadio V y los adultos, colonizaron las ramas del árbol de manera ascendente (Fig. 4B), ovipositando en hojas nuevas con incremento de su población en el mismo árbol (Figs. 4B, 4C y 4D). Los árboles de nogal cafetero después de haber renovado su follaje, sufrieron el ataque de las nuevas generaciones de adultos de la chinche de encaje, impidiendo el desarrollo de estas hojas y causando síntomas de encrespamiento, amarillamiento y necrosis de la lámina foliar. Estas observaciones permiten sugerir que se realicen prácticas que impidan la reinfestación de chinches de hojas desde el suelo, mediante aspersiones de insecticidas químicos o biológicos, incrementos de depredadores en el suelo o prácticas culturales mediante remoción de hojas, en el establecimiento de un programa de manejo integrado de D. monotropidia.

A medida que se agota el alimento disponible (hojas y brotes) los adultos de D. monotropidia vuelan hacia los árboles vecinos a diferentes distancias favorecidos por la velocidad y dirección del viento. En estudios realizados por Fallas et al. (1993), sobre dispersión de adultos de D. monotropidia, encontraron que el mayor número de individuos se capturó en láminas de estereofón de 0,25m X 0,50m cubiertas de plástico blanco impregnado de aceite agrícola, colocadas en círculos concéntricos a una distancia de 2 m y el menor a los 10 m. Se encontraron diferencias entre las trampas colocadas a los 2 y 4 m y entre aquellas a los 2 y 6 m. Algunos de los adultos pueden volar distancias de hasta 10 m, a favor del viento. De acuerdo con lo registrado por la estación meteorológica de la zona, la humedad relativa durante el año de evaluación presentó un valor máximo de 79,3% promedio mensual en marzo de 2009 y un mínimo de 68,8% en enero del 2010 (Fig. 3C). Se presentó déficit hídrico durante junio y agosto, a mediados de septiembre y primeros días de diciembre, bajo las condiciones climáticas del municipio de Manizales (Fig. 3A). La caída de hojas del nogal cafetero ocurrió semanas antes que se presentara el déficit hídrico. La población del insecto disminuyó meses después de la defoliación, debido posiblemente a la carencia de alimento. Esto indica que la defoliación ocurrió antes del tiempo fisiológico reportada para esta especie botánica y que la causa de la defoliación la debió haber ocasionado el insecto.
Según Lewis (1997), el número de insectos es el resultado de nacimientos, muertes y migraciones. Si estos procesos y los factores que influencian en la dinámica poblacional, como los climáticos, pudieran ser predichos; sería posible diseñar estrategias de manejo que proporcionaría alertas tempranas de los crecimientos poblacionales de la plaga.
Determinación de la duración de los estados de vida de D. monotropidia. Bajo condiciones de casa de malla en el vivero forestal de Cenicafé, se obtuvo una cohorte de 144 huevos de D. monotropidia. La duración promedio del ciclo de vida completo fue estimada en 159 días (Tabla 4).

Huevo. La hembra puede copular con varios machos y luego oviposita dentro del tejido foliar casi por completo, principalmente por el envés en las nervaduras centrales y secundarias, en algunas ocasiones en el borde de la hoja y por el haz en la nervadura central. Los huevos, recién puestos, son blanco cremosos, recubiertos con una sustancia transparente que, posteriormente se oscurece. Presentan forma ovalada, con uno de los extremos más agudo formando un cuello que termina en una corona circular blanca brillante donde se ubica el opérculo. Los huevos presentan una longitud promedia de 0,5 mm y a medida que van madurando se tornan de color marrón oscuro. La duración de esta etapa en promedio fue 15,5 ± 0,10 días (Fig. 5).

Ninfa I. Al emerger del huevo son de coloración amarilla y posterior, a un par de horas, se tornan rojizas. A las cinco horas cambian a negro, permaneciendo agregadas alrededor de los restos de los coriones. La duración del primer estadio fue en promedio de 6,5 ± 0,09 días (Fig. 5).
Ninfa II. Recién a la muda es de color rosado y después de seis horas cambia a negro generalizado. Las antenas y las patas son de color blanco translucido. La duración de este estadio en promedio fue 4,5 ± 0,24 días (Fig. 5).
Ninfa III. Cuando ha melanizado completamente el cuerpo de la ninfa se torna de color negro a excepción del tercer artejo antenal y las tibias que son gris claro. Este tercer estadio en promedio dura 5,5 ± 0,22 días (Fig. 5).
Ninfa IV. Cuando ha melanizado completamente es negro a excepción del tercer artejo antenal y las tibias que son claras. En el mesotórax se empiezan a diferenciar los cojines alares. El último segmento antenal es casi la mitad de largo que el tercero. La duración del cuarto estadio fue en promedio de 5,5 ± 0,16 días (Fig. 5).
Ninfa V. Es de forma ovalada y alargada; el tercer artejo antenal es amarillo claro, el cuarto tan largo como la mitad del tercero. El pronoto es de color negro con un par de manchas de color crema en forma de media luna en cada margen anterior; el escutelo es de color blanco-crema. Los cojines alares son de color negro en el área basal y apical y blanco crema en el área medial. El abdomen es negro y aplanado. La duración del quinto estadio fue en promedio de 6,5 ± 0,14 días (Fig. 5).
Adulto. Los machos son menos robustos que las hembras y presentan dimorfismo sexual: el último segmento abdominal es largo y ovalado mientras que en las hembras es corto y aguzado. La duración del adulto fue de 115 ± 2,43 días en promedio (Fig. 5).
De acuerdo con el ciclo biológico obtenido en la casa de malla se pueden presentar ocho generaciones por año. Esto concuerda con lo enunciado por Madrigal (1987) en condiciones de laboratorio, teniendo en cuenta que en los periodos lluviosos la oviposición se reduce y la duración del ciclo de vida se prolonga.
Los ciclos biológicos obtenidos en laboratorio dan una idea aproximada del tiempo de duración de los insectos, ya que allí se pueden controlar factores bióticos, por ejemplo, la depredación y parasitismo y abióticos como las variables relacionadas con el clima. En la casa de malla y con mangas entomológicas se obtiene una idea más cercana a la realidad del ciclo de vida del insecto estudiado. Aunque lo ideal sería estudiar los ciclos biológicos a nivel de campo, éste presenta la dificultad de controlar variables como la precipitación, que es un factor de mortalidad que contrarresta la supervivencia de los individuos en muchas especies de insectos.
Conclusiones y Recomendaciones
La duración promedio del ciclo de vida desde la postura hasta la muerte de los adultos de D. monotropidia fue de 159 días. La duración de huevo a emergencia del adulto fue, en promedio, 44 días. En esta investigación se detectaron dos periodos de mayor número de individuos de D. monotropidia en las hojas afectadas de los árboles de nogal cafetero, febrero a mayo y octubre a noviembre de 2009. Sin embargo, los tiempos más secos coincidieron con las mayores poblaciones en las hojas de los árboles. D. monotropidia ocasiona caída de hojas de manera anticipada a la defoliación fisiológica de la planta, factor que debe influir en el desarrollo y crecimiento de los árboles de nogal. Una vez las hojas infestadas caen al suelo, a pesar de la cantidad de depredadores naturales que se evidenciaron en el estudio, el insecto comienza un proceso de dispersión hacia hojas tiernas de la parte aérea de árboles adyacentes. El manejo de esta plaga con insecticidas químicos o biológicos, deberá realizarse tanto en los árboles infestados, como en las hojas infestadas del suelo, durante los períodos de mayor presencia de la chinche de encaje. Igualmente se recomienda usar estrategias de control biológico por conservación o aumentación de los enemigos naturales, con el fin de incrementar el control en las hojas infestadas en el momento que caen al suelo. Se sugiere considerar como estrategia de control cultural, la remoción de las hojas infestadas en los árboles.
Agradecimientos
Al Ministerio de Agricultura y Desarrollo Rural, convenio 050 proyecto 2007K4683-528-07 “Propuesta de manejo integrado del chinche de encaje Dictyla monotropidia en plantaciones de nogal cafetero Cordia alliodora ubicadas en la zona centro sur del departamento de Caldas”, por su apoyo financiero. A Cenicafé y sus investigadores Juan Carlos López Núñez y al personal de la disciplina de Entomología, a Juan Carlos Ortiz (Museo Entomológico Marcial Benavides - Cenicafé) por su ayuda en la toma de las fotografías del ciclo de vida, a los investigadores Orlando Guzmán M. y Álvaro Jaramillo y al personal de la disciplina de Agroclimatología de Cenicafé. Al personal del Programa ETIA-Forestal de Cenicafé, por su valiosa colaboración en esta investigación. A José Misael Méndez, propietario de la finca “La Ramada”.
Literatura citada
ARGUEDAS, M.; FALLAS, E. 1993. La chinche de encaje del laurel Dictyla monotropidia. Serie Plagas y Enfermedades. Nº 7. ITCR- CIT. 4 p. [ Links ]
CATIE. CENTRO AGRONÓMICO TROPICAL DE INVESTIGACIÓN Y ENSEÑANZA. 1991. Plagas y enfermedades forestales en América Central: Guía de campo. Turrialba, C. R. 262 p. (Manual Técnico No. 4). [ Links ]
DRAKE, C. J.; RUHOFF, F. A. 1965. Lacebugs of the world: A catalog (Hemiptera: Tingidae). Smithsonian Institution United States. Nature Museum Bulletin 243: 634. [ Links ]
FALLAS, E.; ARGUEDAS, M.; BRICEÑO, R. 1993. Dispersión y métodos de cría de Dictyla monotropidia (Hemiptera: Tingidae). Revista Biología Tropical 41 (3): 509-513. [ Links ]
GÓMEZ, D.; OSPINA, C. 2003. Diagnóstico de enfermedades en plantaciones de Nogal Cafetero (Cordia alliodora) y Chaquiro (R. rospigliosii) en la zona cafetera Colombiana. En: Memorias XXIV Congreso de Fitopatología ASCOLFI. 2003. Armenia, Quindío. 31-32 p. [ Links ]
JOHNSON, P.; MORALES, R. 1972. A review of Cordia alliodora (R & p) Oken. Turrialba 22: 210-220. [ Links ]
LEWIS, T. 1997. Thrips as crop pests. CAB International. Wallingford, United Kingdom. 740 p. [ Links ]
MADRIGAL, C. A. 1987. Biología, hábitos, y distribución en Antioquia de la Chinche de encaje Dictyla monotropidia Stål (Hemiptera: Tingidae) en Nogal cafetero Cordia alliodora (Ruiz & Pavón). Revista Colombiana de Entomología 13 (1): 3-11. [ Links ]
PROCUENCA EN MARCHA. 2007. Proyecto forestal para la cuenca del río Chinchiná. Boletín Técnico Nº 26 (Febrero- Marzo), 4 p. [ Links ]
PROEXPORT COLOMBIA. 2009. Sector Forestal: Invierta en Colombia. Bogotá. 1 (1): 1-26. Disponible en: www.inviertaencolombia.com.co. [Fecha revisión: 27 julio 2010]. [ Links ]
PROGRAMA KFW. 2009. PROGRAMA FORESTAL RIO MAGDALENA- FEDERACIÓN NACIONAL DE CAFETEROS DE COLOMBIA. GERENCIA TÉCNICA. [ Links ]
Relación de plantaciones forestales y agroforestales realizadas entre los años 1993 y 2009. Convenios: 9166059, 0066571, 105/05, 089/06 y Midas-F-0157-6-G-234. Compilación Ing. Raúl Jaime Hernández Restrepo. 918 p. [ Links ]
SPEIGHT, M. R.; WYLIE, R. F. 2001. Insect pest in tropical forestry. CABI Publishing. Londres, Reino Unido. 307 p. [ Links ]
