











 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Cited by Google
Cited by Google  Similars in
SciELO
Similars in
SciELO  Similars in Google
Similars in Google 
 Permalink
Permalink
Introducción
En la región Neotropical, específicamente en Paraguay Euschistus heros (Fabricius, 1798) (Hemiptera: Pentatomidae), popularmente conocida como chinche marrón es la plaga más dañina de la soja en estadio reproductivo (Ferreira-Agüero et al., 2018). Ninfas y adultos se alimentan en las vainas perjudicando la calidad y el rendimiento del grano o semilla. Mantener sus poblaciones por debajo del umbral económico es muy difícil debido a la tolerancia a insecticidas químicos tóxicos y las malas prácticas de manejo (Ferreira-Agüero et al., 2018).
En toda Sudamérica no solo han fallado constantemente los insecticidas para el control de E. heros, también desencadenaron efectos secundarios negativos principalmente al ambiente y la salud humana, por tal motivo, Telenomus podisi Ashmead, 1893 (Hymenoptera: Scelionidae) es su agente de control biológico con elevada capacidad de parasitismo y control (De Freitas Bueno et al., 2020).
Por lo tanto, es necesaria la utilización de una alternativa de manejo más racional, donde el parasitismo producido por T. podisi en huevos de chinches Pentatomidae, específicamente sobre E. heros, puede ser de suma importancia por su eficiencia demostrada mediante liberaciones en cultivos de soja y fundamental en el desarrollo de paquetes tecnológicos para su utilización en programas de control biológico (Pernambuco Filho et al., 2022).
La familia Scelionidae presenta una distribución cosmopolita exceptuando las regiones polares, es diversa en regiones tropicales, subtropicales y selvas húmedas (Masner, 1993). En el mundo se han registrado aproximadamente 244 géneros y 3.308 especies (Austin et al., 2005; Masner, 1993). En la Región Neotropical se han registrado 56 géneros y 334 especies, tradicionalmente distribuidos en tres subfamilias: Scelioninae, 44 géneros y 227 especies; Teleasinae, seis géneros y 16 especies; y Telenominae, seis géneros y 91 especies (Fernández, 2000).
El género Telenomus posee al menos 300 especies en la región Neártica y 69 especies la región Neotropical (Johnson, 1984, 1992) y se dividen en 11 grupos de especies consideradas monofiléticas para facilitar su identificación. Telenomus es un parasitoide de huevos que hospeda varios insectos pertenecientes a las ordenes: Diptera, Hemiptera, Lepidoptera, y Neuroptera (Johnson, 1984). En Paraguay no se tiene registro de la ocurrencia natural de especies de Telenomus.
Por lo expuesto, los objetivos de esta investigación fueron reportar la ocurrencia natural de Telenomus sp. aff. podisi y el parasitismo de huevos de Euschistus heros en el departamento de Amambay, Paraguay. También, cuantificar el número de huevos parasitados, estimar el índice de parasitismo y porcentaje de emergencia de los parasitoides.
Materiales y métodos
La investigación fue realizada en cultivos de soja zafra 2021/2022 en seis municipios del departamento del Amambay, Paraguay. Los puntos de muestreo fueron distribuidos y localizados en tres distritos según sus coordenadas geográficas (Tabla 1 y Figura 1).
Tabla 1 Puntos de muestreo localizados por coordenadas geográficas según los distritos y municipios en diferentes zonas de cultivo de soja del Departamento de Amambay, Paraguay.
| Distritos | Municipios | Coordenadas geográficas | |
|---|---|---|---|
| Pedro Juan Caballero | Yvypé | 22°33′57″S | 55°48′03″W |
| 22°36′31″S | 55°48′42″W | ||
| Cerro Cora’ i | 22°38′13″S | 55°53′39″W | |
| 22°41′10″S | 55°50′24″W | ||
| 22°39′17″S | 55°53′35″W | ||
| Potrero Sur | 22°37′32″S | 55°41′34″W | |
| Sanja Pytã | Fortuna Guazú | 22°41′40″S | 55°46′12″W |
| Y’ambué | 22°36′36″S | 55°45′11″W | |
| Cerro Corá | Cerro Corá | 22°36′36″S | 55°45′11″W |
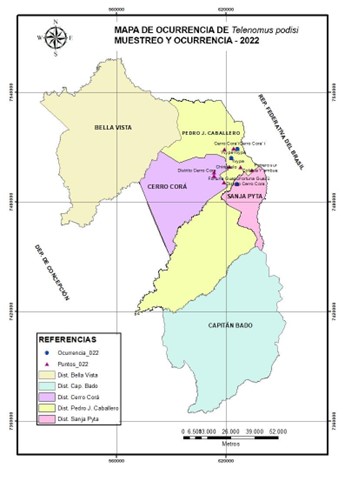
Figura 1 Mapa con puntos de muestreo y presencia de Telenomus sp. aff. podisi en cultivos de soja, en zonas del Departamento de Amambay, Paraguay.
Recolecta de posturas de E. heros.
El muestreo se realizó mediante la recolección manual de las posturas de Euschistus heros en cultivos de soja a través de la inspección aleatoria de 50 plantas de soja para cada zona de muestreo. Las recolecciones de los huevos de E. heros se realizaron al atardecer, fueros recogidos con las hojas, ramas y vainas de la soja en las cuales estaban adheridas las posturas. Para cada localidad se registró la coordenada geográfica con el GPS para indicar los puntos de muestreo y la ocurrencia del parasitoide en el mapa.
Luego las posturas recolectadas fueron depositadas en placas de Petri y acondicionados en conservadora de isopor de 8L para transportar al laboratorio de Protección Vegetal de la Facultad Ciencias Agrarias de la Universidad Nacional de Asunción, filial Pedro Juan Caballero para permitir la emergencia de los adultos.
Recuperación de parasitoides en laboratorio.
Dentro de las placas de Petri que contenían los huevos de E. heros adheridas a las superficies de las hojas, ramas y vainas de la soja se puso algodón humedecido con agua destilada para evitar la deshidratación de los huevos y posteriormente fueron depositadas en cámara climatizada tipo BOD (Biological Oxygem Demand) bajo condiciones controladas (28 ± 2 °C, 65 ± 5 % HR, 14:10 L:O Fotofase), para permitir la emergencia de los parasitoides y/o chinches conforme al tiempo de desarrollo de los insectos que podía variar entre 5 a 7 días aproximadamente. Luego de la emergencia de los parasitoides, las placas con los insectos se coloraron en congelador durante cinco minutos para producir su muerte, en secuencia se depositaron en tubos de vidrio con alcohol 70 % para su preservación.
Observación de características morfológicas en microscopio.
Inicialmente los parasitoides fueron retirados del tubo de vidro con alcohol 70 %, vertidos en una placa de Petri y mediante un alfiler fueron recolectados y depositados sobre una lámina de vidrio para fijar la muestra para la observación en microscopio estereoscopio (4x de magnificación). Los insectos fueron posicionados de acuerdo con las características morfológicas que se pretendía mostrar: vista lateral del insecto adulto hembra y macho con dimorfismo sexual diferenciados por el número de antenómeros (Figuras. 1 A y B), cabeza (Figura. 2 C), alas, anterior y posterior (Figura. 2 D), mesopleura (Figura. 2 E), mesonoto y metanoto (Figura. 2 F) siguiendo las descripciones de Masner (1980) y Johnson (1984). Para la elaboración de figuras con microscopio y cámara digital, las fotografías fueron tomadas con un microscopio binocular Iscope® (4x/10x) equipada con cámara digital Euromex®. Las series de imágenes fueron combinadas con la aplicación Helicon Focus 8 para obtener las imágenes con foco extendido.
Identificación de los especímenes.
Las muestras de parasitoides contenidas en frasco ámbar de 5 mL con alcohol 70% fueron etiquetados y empaquetados dentro de un isopor con un formulario de solicitud de identificación de parasitoides con todos los datos de cada zona y enviados al laboratorio del Instituto Biológico (Secretaría de Agricultura e Abastecimiento, Agencia Paulista de Tecnología dos Agronegocios) de Campinas, São Paulo, Brasil. Los especímenes fueron identificados mediante la llave taxonómica de Johnson (1984) y quedaron depositados en la colección de Insectos Entomófagos “Oscar Monte” en el Laboratorio de Controle Biológico, bajo número de referencia IB-CBE-S-826.
Parasitismo.
En una planilla Excel fueron registrados el número de huevos recolectados, parasitados y adultos emergidos para el cálculo de los siguientes parámetros: índice de parasitismo (%) a través de la formula P = (Nº de huevos parasitados/total de huevos del hospedero) x 100 (Zachrisson et al., 2014). Número de huevos parasitados: (números de huevos parasitados/número de huevos existentes) y adultos emergidos: (número de avispas emergidas/número de huevos parasitados) x 100 (Cingolani et al., 2014; Wengrat, 2022).
Resultados y discusión
Identificación de los especímenes.
Se obtuvo un total de 12 ejemplares (5 hembras y 7 machos) que fueron identificados como Telenomus sp. aff. podisi (Hymenoptera: Scelionidae). La especie solo pudo ser determinada con la finalización de los estudios ejecutados por un equipo compuesto por investigadores del Instituto Biológico (Campinas, SP, Brasil), Escola Superior de Agricultura “Luiz de Queiroz”/Universidad de São Paulo (Piracicaba, SP, Brasil) y The Ohio State University (Columbus, OH, E.U.A.).
Los Telenominae son endoparasitoides primarios de huevos, idiobiontes, es decir, detienen el desarrollo del hospedero de Diptera, Fulgoridae, Hemiptera y Lepidoptera. Las hembras buscan al hospedero en hábitats específicos, por ejemplo, Telenomus podisi Ashmead que es parasitoide de Pentatomidae, busca al hospedero en plantas herbáceas y al igual que otros parasitoides, localizan a su hospedero mediante kairomonas y para reconocerlo se basan en los terpenoides que liberan las plantas al momento de recibir el daño por los Pentatomidae. Las hembras pueden reconocer si el hospedero ha sido parasitado previamente, ya que detectan feromonas externas o probando el huevo con su ovipositor (Masner & Arias-Penna, 2006).
Uno de los caracteres diagnósticos de la llave de identificación para Telenomus es la coloración de la procoxa (Johnson, 1984), donde se observa una gran variabilidad de coloración entre las especies. Entonces estos son separados en morfotipos conforme a la gama de variaciones encontradas para las coxas. Conjuntamente también se evalúan en las antenas las coloraciones del escapo y la proporción del tamaño de la antena en los segmentos A1 y A3 (Wengrat, 2022).
Parasitismo.
Del total de muestras recolectadas se obtuvieron 737 huevos de E. heros, de los cuales, 683 huevos estaban parasitados, con un índice de parasitismo de 92,67 % y un promedio de 22,55 % de parasitoides emergidos. La presencia natural de Telenomus sp. aff. podisi parasitando huevos de E. heros es un fuerte indicio de que este parasitoide tiene las condiciones adaptativas para la supervivencia, colonización y parasitismo de especies de Pentatomidae en condiciones de campo en Paraguay. En su investigación Bueno et al. (2009) informaron que el parasitismo es la característica biológica más importante al evaluar los parámetros biológicos de un insecto con potencial de control de una plaga, ya que eso determina la eficiencia del control biológico en el campo. La mayor eficacia de control de huevos de E. heros en soja (91,12 %) se obtiene con la liberación de 6500 parasitoides por hectárea para mantener la población de las chinches por debajo del nivel de acción propuesto por el MIP-soja (Pernambuco Filho, 2022).
El Municipio donde se registró el mayor número de huevos parasitados fue Yvypé con 465 huevos parasitados que presentaron 194 adultos emergidos con un índice de parasitismo del 93,73 % y 41,72 % de parasitoides emergidos. El segundo municipio con mayor ocurrencia de huevos parasitados fue Cerro Cora´í con 162 huevos parasitados y un índice de parasitismo del 91,52 %. El porcentaje de emergencia de adultos fue de 9,87 %. El tercer municipio con menor número de huevos parasitados fue Fortuna Guazú con 56 huevos parasitados, de los cuales emergieron 9 parasitoides adultos con un índice de parasitismo de 90,32% y el porcentaje de emergencia de adultos fue de 16,07 %. En los municipios de la Colonia Raúl Ocampos Rojas “Chirigüelo”, Potrero Sur y Colonia Yámbué no se detectaron huevos de E. heros parasitados.
Los enemigos naturales están ampliamente distribuidos en diferentes regiones, donde pueden alcanzar tasas de parasitismo natural de 60 % a 80 % (Corrêa-Ferreira, 2002). Por ejemplo, en su revisión De Abreu et al. (2015), informaron que T. podisi ocurre con más del 80 % de parasitismo en E. heros en varias regiones de Brasil, corroborando los resultados de esta investigación, donde obtuvo un promedio de 92,64 % de parasitismo de E. heros.
Tasas de parasitismos semejantes a los verificados en este estudio se han relatado en cultivos de soja del Brasil con un estudio de eficacia de la liberación masiva de T. podisi para el control de huevos de E. heros donde el índice de parasitismo fue de 78,29 % y 85,39 % con la liberación de 5000 y 6500 parasitoides/ha, respectivamente, lo que mantuvo la población de chinches por debajo del umbral de acción (Pernambuco Filho, 2022). De igual manera, en Panamá, se registró parasitismo entre 80 % y 85 % de T. podisi en huevos de chinches del arroz Oebalus insularis (Stal, 1872) (Hemiptera: Pentatomidae) en diferentes áreas de producción de arroz (Zachrisson et al., 2014).

Figura 2 Adultos de Telenomus sp. aff. podisi con dimorfismo sexual, vista lateral: A. Hembra (♀) antena clavada. B. Macho (♂) antena moniliforme.
Una de las principales características morfológicas es la presencia de dimorfismo sexual en las antenas de adultos de Telenomus sp. aff. podisi, la hembra (♀) presenta antena del tipo clavada con 10 antenómeros, mientras que el macho (♂) presenta antena del tipo moniliforme con 11 antenómeros (Figuras. 2A y B). La estructura completa de la antena: pecidelo, flagerómero y antenómeros de la hembra posee aproximadamente 0,8 mm, mientras que el del macho, 1 mm de largo aproximadamente. El pedicelo y el flagerómero son de color amarillento, mientras que los antenómeros son negruzcos y gris cubiertas por cilios.
En la llave taxonómica para los géneros Neárticos de Platygastroidea que parasitan huevos de Pentatomidae, las antenas de las hembras de Telenomus Haliday presentan sensilas basicónicas en 4 antenómeros apicales, raramente 5 o 6. (T. astrictus, T. calvus, T. goliathus, T. grenadensis, T. persimilis, T. podisi, T. sanctiventris, T. scaber) (Talamas et al., 2015). Los grupos de especies de T. podisi se les puede identificar por tener dos o más pares de setas sublaterales; cabeza transversa; frente suave; carina metapleural presente y son parasitoides de Pentatomidae y Scutelleridae (Hemiptera: Heteroptera) (Johnson, 1984).
El cuerpo del parasitoide está formado por la cabeza, tórax y abdomen que son de coloración negra brillante de aspecto metálico, mientras que, en las piernas, la coxa es de color marrón; ya el fémur y trocánter son amarillentos y revestidos de cerdas (Figuras. 2A y B). En Telenomus, el cuerpo es más o menos cilíndrico, casi tan largo como ancho o apenas ligeramente más largo, en vista lateral con el escutelo acentuadamente más alto que el metanoto y el propódeo inclinado (Masner, 1980).
La región de la cabeza se caracteriza por poseer un par de ojos con ocelos de coloración gris metálico. También se destaca en la parte superior de la cabeza una fosa frontal (fp) profunda y depresión frontal (fd). La frente es lisa (fl) y brillante de color negro y sin cerdas, aspectos fáciles para el reconocimiento de Telenomus en comparación a otros géneros (Figura. 2 C).
En la cabeza de T. podisi, el vértice suavemente redondeado sobre occipucio al menos medialmente, redondeado o angulado detrás de los ocelos laterales, uniformemente coriáceo; carina hiperoccipital no desarrollada; occipucio coriáceo, escultura que llega a la carina occipital; carina occipital completa medialmente, crenulada; bandas orbitales presentes, frente lisa por lo demás; setas ocelares presentes o no; depresión frontal débilmente desarrollada; frente ligeramente abultada entre inserciones antenales y órbitas internas; ojos peludos; órbitas internas angulado a nivel de los ocelos laterales; espacio malar liso; templos no abultados, generalmente se cae bruscamente detrás de los ojos hacia la carina occipital (a veces en las sienes débilmente redondeado justo detrás del ojo), no estriado, bandas de escultura coriácea a lo largo de las órbitas posteriores que no alcanzan la carina occipital (Ramirez-Ahuja, 2021).
Las alas anteriores (aa) y posteriores (ap) son hialinas, transparentes y translucidas con cilios en la totalidad de su superficie, destacándose los cilios marginales (cm). En el ala anterior (aa), la vena marginal (vm) se extiende hasta la vena estigmal (ve), siendo la vena marginal más extensa que la estigmal. También presenta una vena submarginal (sm) y posmarginal (pm). El ala posterior (ap) solamente tiene la vena marginal (vm) (Figura. 2 D).
En la región del tórax, específicamente mesotórax, se destaca la mesopleura (mspl) con una fosa mesopleural (fm) profunda, ligeramente transversal, con surco levemente pronunciado que se extiende en dirección a la tegula, localizada en la base de las alas. Otras características resaltantes en esta región son el surco mesepimeral (mees) y la carina acetabular (ac) (Figura. 3E).
En T. podisi el mesoscutum es fuertemente convexo, coriáceo-pustulado, sin elementos longitudinales en la escultura, la escultura de fondo se desvanece posteriormente; escudo liso, setoso, bases setales pustuladas, fóveas submarginales casi igual en tamaño a los pinchazos dorsales; dorso ampliamente triangular, superpuesto propódeo, reticulado punteado grueso y carina acetabular (Ramirez-Ahuja, 2021).
En la vista posterior del tórax y abdomen (Figura. 3F) se perciben algunas estructuras características. El mesonoto (ms) se encuentra el escutelo (sc) y el propódeo (pr), segmento del mesossoma posterior al metanoto (mn); originalmente el primer tergito abdominal esculturado se funde con el tórax. El terguito 2 (T2) terguito del metasoma, en Telenomus puede ser identificados fácilmente por poseer la cabeza en comparación a otros géneros, por la forma del (T2) segundo terguito metasomico en vista dorsal que es más largo que ancho con la frente predominantemente lisa y brillante (Figura. 3F) (Wengrat, 2022).

Figura 3 C-F. Características morfológicas de adultos de Telenomus sp. aff. podisi: C. Cabeza vista frontal, D. Alas, E. vista lateral, F. vista dorsal. (fp) fosa frontal, (fl) frente lisa, (oc) carina occipital; aa ala anterior, (vm) vena marginal, (st) vena estigmal, (cm) cerdas marginales, (sm) vena submarginal, (pm) vena posmarginal; ap ala posterior; (mspl) mesopleura, (fm) fosa mesopleural, (mees) surco mesepimeral y (ac) carina acetabular;, (ms) mesonoto, (sc) escutelo, (pr) propódeo, (t1) terguito 1, (t2) terguito 2, (mn) metanoto.
Conclusiones
Se identificó únicamente a Telenomus sp. aff. podisi como parasitoide de huevos de Euschistus heros en soja en el departamento de Amambay, Paraguay.
El porcentaje de parasitismo promedio de Telenomus sp. aff. podisi fue de 92,67 % en huevos de Euschistus heros con un promedio de 22,55 % de parasitoides emergidos.

