










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Cited by Google
Cited by Google -
 Similars in
SciELO
Similars in
SciELO -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkRevista Colombiana de Ciencias Pecuarias
Print version ISSN 0120-0690On-line version ISSN 2256-2958
Rev Colom Cienc Pecua vol.18 no.1 Medellín Jan./Apr. 2005
El gas metano en la producción ganadera y alternativas para medir sus emisiones y aminorar su impacto a nivel ambiental y productivo
Juan C Carmona1, Zoot Esp; Diana M Bolívar2, Zoot MSc; Luis A Giraldo2, Zoot MSc
1 Universidad de Antioquia, Facultad de Ciencias Agrarias, Ciudadela Robledo, Medellín, Colombia.
2 Universidad Nacional de Colombia, sede Medellín, Facultad de Ciencias Agropecuarias, Departamento de Producción Animal,
A.A. 586, Medellín, Colombia.
jccarmona@agronica.udea.edu.co
(Recibido: 2 marzo, 2004; aceptado: 8 febrero, 2005)
Resumen
Los rumiantes son grandes contribuyentes al calentamiento global y deterioro de la capa de ozono, por la liberación de altas cantidades de gases a la atmósfera, entre ellos, el gas carbónico y el metano. El metano producido se genera principalmente por los procesos fermentativos del alimento que ingresa al rumen. El principal factor biótico a nivel del rumen en la producción de metano son las bacterias anaerobias metanógenas. Estas bacterias utilizan diferentes sustratos para la producción de metano, pero los principales son el H2 y el CO2. La eliminación de estos gases, principalmente del H2 implican la remoción de un factor implicado en la estabilidad del pH ruminal siendo este esencial para una óptima fermentación. Pero a la par se considera la producción de metano como una pérdida de energía potencialmente utilizable. Los efectos de las bacterias metanógenas son dependientes principalmente de los sustratos presentes en la dieta y de las interacciones con otras poblaciones. Intervenciones en la alimentación ofrecida a los animales, orientadas hacia optimizar el proceso de fermentación ruminal, generalmente repercuten en una mejora de los parámetros productivos y reproductivos, debido, entre otros aspectos, a una mejor utilización de la energía. Además, la disminución de las emisiones a la atmósfera cobra gran importancia en la protección del medio ambiente. Diversas evidencias muestran que la tasa de emisión de metano por fermentación ruminal, está relacionada con las características físico-químicas de la dieta, las cuales afectan el nivel de consumo y la frecuencia de alimentación. Por esto una subnutrición contribuye a incrementar las emisiones de metano. La posibilidad de limitar las emisiones de metano por el ganado en sistemas de producción tropical, provee beneficios económicos y medioambientales. Una opción de reducción consiste en la sustitución de tecnologías convencionales por nuevas alternativas concomitantes con una adecuada producción y mínimos efectos medioambientales. Dentro de estas alternativas de potencial uso en trópico está el manejo de pasturas, tendiente hacia mejorar su calidad. Una alternativa de amplio potencial y que hasta el momento ha tomado fuerza por sus múltiples beneficios son los sistemas silvopastoriles, pero poco se ha investigado su efecto sobre la producción de metano ruminal. Para la determinación de emisiones y la eficiencia de las alternativas implementadas, se debe recurrir a metodologías apropiadas que permitan extrapolar los resultados a las condiciones reales in vivo. Dentro de estas alternativas, el sistema in vitro RUSITEC, ha presentado una alta correlación con las características propias del animal. El objetivo de esta revisión es visualizar diferentes factores que determinan las emisiones de metano, alternativas de medición y opciones para disminuir la producción de este gas en los sistemas ganaderos, en aras de una producción más eficiente y que proteja el medio ambiente.
Palabras clave: efecto invernadero, metanogénesis, rumiante, rusitec, silvopastoril.
Introducción
Los bovinos poseen un sistema digestivo que tiene la capacidad de aprovechar y convertir material fibroso con altos contenidos de carbohidratos estructurales, en alimentos de alta calidad nutritiva, la carne y la leche. Sin embargo por sus características innatas, este mismo sistema digestivo también produce metano, un potente gas con efecto invernadero que contribuye con aproximadamente el 18% del calentamiento global ocasionado por actividades productivas con animales domésticos, superado sólo por el CO2 (20).
La producción de metano en los últimos años ha tomado gran importancia en la producción animal debido a sus efectos negativos en el medio ambiente (6). Otro aspecto de relevancia, es la eficiencia energética de los sustratos alimenticios fermentados en el rumen, la cual disminuye en proporciones variables dependiendo de las características de la dieta, debido a que las emisiones de gases, específicamente de metano, involucra pérdidas a través del eructo. Johnson y Johnson (13) señalan que, el metano colabora en los efectos climáticos directamente, a través de su interacción con la energía infrarroja e indirectamente a través de las reacciones de oxidación atmosféricas que producen CO2. Por esto se considera que en la actualidad los sistemas de producción animal sostenibles deben propender por una menor producción de metano.
La manipulación de la dieta de los rumiantes se considera una alternativa viable para aminorar la producción de metano y a la par disminuir las pérdidas energéticas en el animal. Esta alternativa toma mayor fuerza en las condiciones de trópico, donde la mayoría de los sistemas de producción ganadera tienen bajos rendimientos debido a las dietas de baja calidad. Los reportes en la literatura señalan que los sustratos de baja calidad que, generalmente están relacionados con bajo consumo debido a su baja tasa de pasaje, no sólo tienen efecto sobre bajos rendimientos por animal, sino que también involucran aspectos tan importantes como lo es el incremento de las emisiones de metano. El mejoramiento de las características nutricionales del forraje y la implementación de sistemas estratégicos de suplementación, como la presencia de otros estratos vegetales en el ecosistema pastura, tipo sistemas silvopastoriles, pueden mejorar las características fermentativas a nivel ruminal, reflejándose en mayor productividad y generalmente en una disminución en las emisiones de metano.
El objetivo de esta revisión es analizar los diferentes factores involucrados en las emisiones de metano de los animales rumiantes, las diferentes alternativas de medición y las opciones para disminuir la producción de este gas en los sistemas ganaderos, en aras de una producción más eficiente y concomitante con la protección del medio ambiente.
Producción de gas metano en rumiantes y su impacto ambientalLa agricultura y la producción pecuaria contribuyen ampliamente a las emisiones antropogénicas de metano (CH4), dióxido de carbono (CO2) y óxido nitroso (N2O) a la atmósfera. El aumento de las concentraciones de estos gases provoca un calentamiento de la superficie terrestre y la destrucción de la capa de ozono en la estratosfera (24). Dentro de la gama de gases a los que se les atribuye efecto invernadero, se considera el CO2 el más abundante y el que actualmente tiene un mayor aporte al incremento del calentamiento global. Hoy día las concentraciones de metano son inferiores a las de CO2, sin embargo el primero, se está incrementando rápidamente y además posee un efecto 21-30 veces más contaminante con respecto al CO2,considerándose que en el tiempo el metano pueda ser predominante (18, 19). Las tasas de acumulación de metano y dióxido de carbono en la atmósfera han cambiado drásticamente en los últimos años presentándose un incremento de forma exponencial (23).
Cerca de 500 millones de toneladas métricas/año de metano ingresan a la atmósfera debido a actividades antropogénicas y fenómenos naturales. A esta tasa se
espera que el metano cause cerca del 15-17% del Tabla 1), pero el grado de incidencia y la proporción calentamiento global. Actualmente se tienen definidas exacta de muchas de estas fuentes no son claras las fuentes de metano causantes de este efecto (véase (13)

Kurihara et al (16) y Johnson y Johnson (13), señalan que las emisiones de gas metano por el ganado bovino, están estimadas en 58 millones de toneladas/ año, lo que representa el 73% del total de emisiones (80 millones) de todas la especies domésticas. McCaughey et al (18) y Moss et al (22) indican que los animales domésticos, principalmente el ganado bovino son responsables de aproximadamente el15% de la producción de metano global. Otros contribuyentes significativos son los pantanos naturales (21%), los cultivos de arroz (20%), pérdidas por combustión de hidrocarburos (14%), combustión de biomasa (10%) y rellenos sanitarios (7%) (18).
Las características de la dieta tienen un gran efecto en la producción de gas metano a nivel global, de ahí que países con pocas limitaciones alimentarias para sus ganados, reportan datos de menores emisiones de gas metano y mayores eficiencias energéticas. Kinsman et al (15), corrobora lo anterior al mencionar que, en los países en vía de desarrollo, las emisiones son aproximadamente de 55 kg CH4/año por animal, en contraste a lo reportado en países desarrollados, de 35 kg CH4/año por animal.
Según González y Rodríguez (11), la proyección de emisiones de metano en Colombia, a partir de actividades en el sector pecuario para el 2010, representarán el 70% de la participación de los gases de efecto invernadero (GEI). Siendo un 95% de este total, las emisiones digestivas del ganado de leche y de carne.
Generalidades metabólicas a nivel ruminalLa actividad ganadera produce entre 15-20% de la emisión mundial de gas metano (18, 20, 21). El ganado bovino emite gas metano porque en su proceso digestivo, que ocurre bajo condiciones anaeróbicas, participan diferentes tipos de bacterias. Éstas, degradan la celulosa ingerida a glucosa, que fermentan luego a ácido acético y reducen el dióxido de carbono, formando metano en el proceso. La emisión de metano representa energía alimenticia que se transforma en forma de gas y no es aprovechada por el animal (18, 19, 20)
La eliminación de metano vía eructo en el ganado inicia aproximadamente a las cuatro semanas de vida, cuando los alimentos sólidos empiezan a ser retenidos en el retículo-rumen, aumentándose la fermentación y la producción de gases a medida que el retículo-rumen se va desarrollando. Se considera que en sistemas de producción de alta tecnificación la producción anual de metano en animales adultos está entre 60 y 126 kg. (13). DeRamus et al (8), igualmente reportan en sus investigaciones, que las emisiones anuales de metano por novillas de carne en pastoreo estuvieron entre 32 y 83 kg. y entre 60 y 95 kg. para vacas adultas, que pastoreaban diferentes tipos de praderas. El dato más alto en cada tipo de animal, corresponde a gramíneas de baja calidad nutricional, con sistemas de pastoreo continuo y baja disponibilidad forrajera, mientras que los datos más bajos corresponden a praderas mejoradas, a sistemas de pastoreo rotacional, fertilización y con alta disponibilidad de forraje (véase Figura 1). En estos reportes, se observa, que de acuerdo a las condiciones de la dieta, las emisiones de metano pueden variar ampliamente, indicando que las características nutricionales de la pastura tienen un efecto marcado en la producción de dicho gas.

McCaughey et al (19), reportan que el 87% de la producción de metano se da en el rumen, y 13% en el tracto digestivo posterior. De este último, aproximadamente el 89% es absorbido hacia la sangre y expirado a través de los pulmones. Esto indica que cerca del 98% del total de metano producido por los rumiantes puede ser expirado a través de la boca y los orificios nasales.
La producción de metano en los bovinos normalmente representa entre 5.5-6.5% del total de energía potencial consumida en la dieta, sin embargo valores entre 2-12% se reportan en condiciones de pastoreo en zonas templadas (2, 16, 30). Pero cuando la alimentación es con forrajes de baja calidad nutritiva, la producción de metano puede representar entre el 15 y el 18% de la energía digestible. Se considera que la corrección de estas deficiencias nutricionales podría reducir estos valores hasta un 7% (20).
Las pasturas en el trópico, generalmente son de regular a baja calidad, por la cantidad de material lignocelulósico de poca digestibilidad y por los bajos tenores de otros nutrientes, principalmente proteína y carbohidratos solubles. Presentándose excepciones, donde algunas pasturas mejoradas y sometidas a programas de fertilización y manejo en general, permiten un mayor aporte de nutrientes y por ende un mejor comportamiento de los animales desde el punto de vista productivo, reproductivo y sanitario.
Johnson y Johnson (13) indican que los dos principales factores responsables de las variaciones en la producción de metano son: la cantidad de carbohidratos fermentados en el retículo-rumen, lo cual implica diversas interacciones dieta-animal, que afectan el balance entre las tasas de fermentación de estos carbohidratos y la tasa de pasaje. El otro mecanismo es la relación de ácidos grasos volátiles (AGV) producidos, la cual regula la producción de hidrógeno y la subsecuente producción de metano.
El aspecto de mayor impacto en la metanogénesis es la relación ácido acético:ácido propiónico. Si esta relación llega a 0.5 la pérdida energética puede ser de 0%. Pero si todos los carbohidratos fuesen fermentados a ácido acético y no se produjera propiónico las pérdidas energéticas podrían llegar a ser del 33%. La relación acético:propiónico puede variar entre 0.9 a 4, por lo tanto las pérdidas por metano varían ampliamente (13).
Moss et al (22) señalan que la estequiometría de las principales rutas de fermentación se resumen así:
Reacciones productoras de H2:
Glucosa → 2 piruvato + 4 H2
Piruvato + H2O → Acetato (C2) + CO2 + 2H
Reacciones que utilizan de H2:
Piruvato + 4H → Propionato (C3) + H2O
2 C2 + 4H → Butirato (C4) + 2H2O
CO2 + 8H → Metano (CH4) + 2H2O
Cuando sustancias reducidas son transferidas de bacterias ruminales fermentadoras de carbohidratos a bacterias metanógenas, el acetato se incrementa y generalmente el propionato disminuye. El acetato es preponderante bajo dichas condiciones pero no se considera precursor significativo de metano en el rumen (28). Moss et al (22) al respecto señalan que el acetato y el butirato promueven la producción de metano, mientras que la formación de propionato puede ser considerada como una forma competitiva en el uso del H2 en el rumen.
Las emisiones de metano normalmente no se cuantifican de forma independiente cuando se discriminan los tipos de energía, por lo tanto, estas emisiones han sido relativamente ignoradas como explicación de la eficiencia alimenticia. Muchos valores de energía metabolizable involucran la estimación de excreción de metano de los carbohidratos digestibles y no consideran la asociación negativa entre la producción de metano y la formación de propionato. Sin embargo, la baja eficiencia está asociada con una alta proporción acetato:propionato, lo que involucra la producción de metano (29).
Moss et al (21) indican que los modelos basados en la estequiometría de la fermentación, se han usado para predecir la producción de metano, implicando una relación negativa entre la proporción molar de propionato y la excreción de metano por unidad de sustrato fermentado. También reportan que la reducción en la proporción molar de propionato se asocia con un incremento en la excreción de metano por kilogramo de materia orgánica fermentada. Sin embargo estos autores observaron que estos modelos ignoran la fermentación de sustratos no carbohidratados como la proteína, la cual debe ser considerada.
De forma similar Van Kessel y Russell (27), señalan tres aspectos: 1) las bacterias metanógenas son sensibles a pH bajos, 2) la disminución en la relación acetato:propionato dependiente del pH es posiblemente causada por una inhibición de la metanogénesis, y 3) la inhibición de la metanogénesis es causada por la toxicidad de los ácidos de la fermentación que se produce a un pH bajo.
Otro aspecto importante a considerar es el incremento térmico negativo, producto del metabolismo del formato, el cual se relaciona con la producción de metano, debido a que el formato es un precursor de éste (29).
Un factor importante cuando se relaciona la producción de metano con el metabolismo ruminal, es el efecto del pH. Así, Van Kessel y Russell (28) señalan, que cuando bacterias provenientes de vacas alimentadas con forraje fueron incubadas en un medio basal que contenía 100 mM de acetato (pH 7.0), el hidrógeno y el dióxido de carbono se convirtieron en metano, pero la tasa de producción de este fue dependiente del pH. Estos autores observaron que la producción de metano disminuyó drásticamente a pH menores a 6.5, y virtualmente no se produjo a pH menores a 6.0 (véase Figura 2).
La dependencia de pH de las bacterias metanógenas, aporta un factor importante para la predicción de la producción de metano. Dietas con forrajes de baja calidad no causan una significante disminución en el pH ruminal y están asociadas a una alta producción de metano. Mientras que dietas con altos contenidos de alimentos concentrados, generalmente disminuyen la producción de metano, pero sólo si el consumo es lo suficientemente alto para se causar una reducción en el pH ruminal (28). Moss et al (21) indican, que en dietas basadas en forrajes, pero con bajo pH, se disminuye la metanogénesis, independiente de la formación de propionato.
Población microbiana y su efecto en la metanogénesis
Aunque existe gran cantidad de microorganismos en todo el sistema digestivo del rumiante, solo los del rumen mantienen una compleja simbiosis con el hospedero. Las bacterias, protozoos y hongos en el retículo-rumen son responsables de la digestión de nutrientes, pero igualmente de otros aspectos que afectan de forma positiva y eventualmente de forma negativa, al animal en sí y al medio ambiente.
El metano es producido por microorganismos del rumen durante la fermentación anaeróbica de carbohidratos solubles y estructurales principalmente, siendo estos últimos preponderantes en dietas basadas en forrajes (16).
El metano es producido por las bacterias metanógenas archaea (25, 29, 30), un grupo microbial filogenéticamente distinto de las eubacterias (verdaderas bacterias) (29, 30). Moss et al (22) señalan que unas y otras divergen en algunos aspectos, entre los que resaltan que las Archaea no tienen polímeros de peptidoglicanos en su pared celular y los lípidos intracelulares son diferentes en composición.
La producción de metano en la naturaleza se da principalmente por dos vías:
- 4H2 + CO2 → CH4 + 2H2O
- CH3COO-+ H+ → CH4 + CO2
En el rumen la producción de metano se da principalmente por la primera vía, debido a que esta requiere menos tiempo para la generación de las poblaciones (4-12 h.). La segunda vía, denominada aceticlástica, no tiene tanto efecto debido a que los tiempos de generación para las poblaciones metanógenas deben ser mayores (17, 30).
Las bacterias metanógenas incluyen: Methanobrevibacter ruminantium, Methanobacterium formicicum, Methanomicrobium mobile. Otra especie, Methanosarcinas, es frecuente sólo en ovejas con dietas basadas en melaza (25, 29, 32).
Las bacterias metanógenas constituyen una clase especial en la población ruminal por su papel en la regulación de la fermentación total al eliminar H2. Como ya se mencionó, la reducción de CO2 con H2 es el método primario por el que se produce CH4 en el rumen, sin embargo algunas bacterias metanógenas como Methanosarcina barkerii, utilizan metanol, metilamina y acetato para producir CH4. Al mantener baja la concentración de H2, mediante la formación de CH4, las bacterias metanógenas promueven el crecimiento de otras especies bacterianas y permiten una fermentación más eficaz (32).
La eliminación de H2 por las especies metanógenas estimula a importantes especies productoras de H2 tales como Ruminococcus albus, Ruminococcus flavefasciens, Selenomonas ruminantium a producir más H2 y así, alterar su metabolismo hacia vías con mayores rendimientos de energía (32).
Weimer (30) señala que, la producción de metano es una vía aceptora de electrones, en la que la metanogénesis continuamente remueve H2, un producto de la fermentación que al acumularse puede disminuir la degradación de la materia orgánica. De acá se desprende la idea de que la reducción o eliminación de la metanogénesis puede requerir el establecimiento de otras rutas aceptoras de electrones. Este aspecto se opone a los conceptos tradicionales sobre la ineficiencia de la metanogénesis. De acuerdo a trabajos reportados por Yokoyama y Johnson (32), la producción de metano, en lugar de representar ineficiencia para el rumiante, promueve una fermentación más eficaz y mayores rendimientos en la síntesis de ATP al mantener baja la concentración de H2. Esto implica que mayores rendimientos de ATP determinan la formación de más células microbianas con lo que aumenta la proteína disponible para el rumiante.
Los protozoos ruminales también juegan un papel importante en la producción de metano, particularmente cuando el ganado es alimentado con dietas altas en concentrado. Las bacterias metanógenas se han observado adheridas a especies protozoales, sugiriendo esto una posible trasferencia interespecífica de H2 (13, 22). La defaunación de animales recibiendo dietas con alto contenido de concentrados puede llegar a disminuir la producción de metano hasta aproximadamente la mitad. Sin embargo, la defaunación de animales recibiendo dietas altas en forraje no reducen significativamente las pérdidas de metano (13).
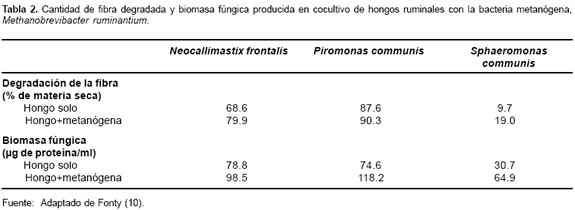
Diversos autores citados por Fonty (10) y Dehority y Tirabasso (7), mencionan un sinergismo entre algunos hongos y bacterias metanógenas, señalando que estas asociaciones incrementan los hongos y la tasa de hidrólisis y degradación de la celulosa (véase Tabla 2). También se ha encontrado que esta asociación incrementa la producción de acetato a expensas de la reducción de lactato y etanol (véase Tabla 3)
Efectos de la dieta en la producción de metano y alternativas para disminuir sus emisionesHay evidencias que muestran que la tasa de emisión de metano, por fermentación entérica, se relaciona con el alimento consumido. También se señala que entre los factores que influyen en su producción están las características físicas y químicas del alimento, las cuales afectan directamente el nivel de consumo y la frecuencia de alimentación. Por tanto una subnutrición contribuye a incrementar los niveles de emisión de metano (20).

Diversos autores señalan que la energía no aprovechada debido a la producción y eliminación de gas metano se debe a muchos factores. Estos incluyen: cantidad y tipo de alimento, manipulación de la fermentación ruminal, adición de lípidos, tipo de carbohidrato en la dieta y procesamiento de los forrajes (13). Estos factores se convierten por tanto en factores alternativos para la disminución de la metanogénesis.
Adición de lípidos
Johnson y Johnson (13), señalan que la grasa en la dieta de los rumiantes afecta la producción de metano por diversos mecanismos, incluyendo la biohidrogenación de los ácidos grasos insaturados, el aumento en la producción de ácido propiónico y la inhibición de protozoos. Se ha demostrado que la adición de ácidos grasos poliinsaturados de cadena larga disminuye la metanogénesis porque se convierte en una alternativa metabólica para el hidrógeno. Sin embargo, la cantidad total de hidrógeno usado en los procesos de biohidrogenación de los ácidos grasos insaturados endógenos es pequeña (1%) comparado con el que es usado para la reducción de CO2 a metano (48%), la síntesis de AGV (33%) y la síntesis bacterial (12%). Dohme et al (9) igualmente señalan que grasas con altas cantidades de ácidos grasos de cadena media pueden ser efectivos en la reducción de metano y de las poblaciones protozoales. Una proporción de bacterias metanógenas es endo y ectosimbióticas con los protozoos. Por lo tanto la reducción del número de protozoos aparentemente contribuye a una declinación en la población de metanógenas (9), además se supone que la toxicidad de algunos ácidos grasos de cadena larga y media sobre las bacterias metanógenas también tienen un gran efecto (9, 22).
Adición de ionóforos
La adición de ionóforos, particularmente la monensina, puede reducir el consumo de alimento entre un 5-6%, disminuir la relación acetato:propionato y disminuir las emisiones de metano (13, 22). Johnson y Jonson (13) y Moss et al (22), indican que la reducción en la producción de metano en animales suplementados con ionóforos es probable que se relacione con la reducción en el consumo de alimento (cuando es de alta calidad), debido al incremento en la eficiencia fermentativa y no por un efecto directo en las poblaciones metanógenas. En este sentido Van soest (29), señala que los ionóforos no son inhibidores directos de las bacterias metanógenas. Estos, lo que hacen es restringir la producción de hidrógeno y por ende la formación de metano. Los verdaderos inhibidores incluyen los alógeno-metanos y metilderivados.
Tipo de carbohidrato
El principal componente en la producción de metano es el tipo de carbohidrato y la tasa de fermentación (6). El tipo de carbohidrato fermentado afecta la producción de metano probablemente a través de impactos en el pH y la población microbial. Así, la fermentación de los carbohidratos fibrosos producen una alta relación de acetato:propionato y una alta producción de metano. Pero dependiendo de la tasa de degradación de la fibra y del consumo relacionado, la producción de metano varia. Este aspecto se evidencia en algunos subproductos de destilería que tienen buena cantidad de fibra, pero de buena digestibilidad, resultando en disminuciones en la producción de metano (13).
Moss et al (22) indican que con dietas altas en almidón se favorece la producción de propionato y se disminuye la relación metano/materia orgánica fermentada en el rumen. Como se discutió previamente, el efecto de estas dietas sobre el pH ruminal pueden explicar la disminución en las emisiones de metano. Chandramoni et al (6) señalan que hay menos producciones de metano con dietas ricas en almidones que con dietas donde la fibra es preponderante. Igualmente Johnson et al citados por Chandramoni et al (6) indican que se da una disminución en la producción de metano cuando se incrementa el consumo de fuentes energéticas. Altas fermentaciones propiogénicas (indicativas de reducciones de metano) son comunes a los sistemas de alimentación donde se suministran altas cantidades de concentrados.
Kurihara et al (16) reportan que la producción de metano (gr/día) en ganado alimentado con Rodhes (Chloris gayana) fue mayor que en los animales alimentados con Ángleton (Dichantium aristatum) y aun mayor que los alimentados con dieta alta en grano. Siendo estos dos tipos de forrajes característicos de zonas tropicales, la producción de metano es mayor y la utilización de la energía menor que lo observado en los forrajes de zonas templadas. Estos aspectos en la dieta con alto grano, fueron similares a los reportados por diversos autores en dietas tradicionales de de zonas templadas. En este trabajo se presume que la alta producción de metano en forrajes tropicales está relacionada con los altos niveles de fibra y de lignina, a los bajos niveles de carbohidratos solubles y a su baja digestibilidad.
Nivel de consumo relacionado con características de la dieta
Con altos consumos de dietas de buena digestibilidad, se presentan menores niveles de energía no aprovechada, debido a menores producciones de metano (13). Un aspecto relacionado con el nivel de consumo es la tasa de pasaje del alimento, así Moss et al (22,) señalan que la producción de metano se redujo en aproximadamente 30% cuando la tasa de pasaje de las fases líquida y sólida se incrementaron de un 54 a un 68%. Los mismos autores señalan que las emisiones de metano se encuentran relacionadas con la cantidad de materia orgánica digestible en el rumen debido a que más del 50% de la digestión ocurre allí.
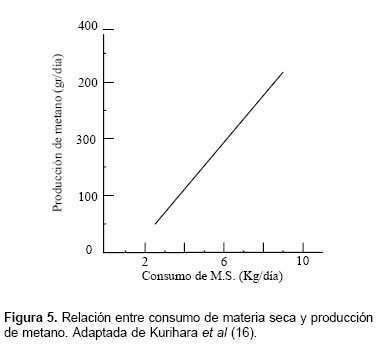
Kurihara et al (16) señalan que se da una asociación lineal entre la relación alimento:ganancia y la producción de metano, lo que presumiblemente puede explicar que, un incremento en la calidad de la dieta sea un medio práctico para reducir la producción de metano (véase Figura 3). Preston y Leng (23) reportan que, rumiantes alimentados con heno de baja calidad pueden generar metano a nivel de 2 kg/kg. de carne producido, en cambio con una suplementación con urea, minerales y proteína no degradable en rumen, la producción de metano se puede reducir hasta 0.36 kg. de carne; encontrando una proporción de energía digestible fermentada a metano de 8 y 15% para la dieta suplementada y la sin suplemento, respectivamente. Kurihara et al (16) señalan que la asociación entre ganancia de peso y producción de metano es de tipo curvilíneo, sugiriendo, que con dietas que posibiliten una respuesta animal alta en términos de ganancia de peso, la producción de metano es menor (véase Figura 4). Respecto al consumo de materia seca, este fue lineal respecto a la producción de metano (véase Figura 5). La tendencia de estos parámetros es similar a los reportados por diversos trabajos citados por Kurihara et al (16), respecto a forrajes tropicales de regular calidad.


Procesamiento de los forrajes
El picado y peletizado de los forrajes son los factores más preponderantes en el procesamiento de estos que pueden disminuir fuertemente la producción
de metano. Este aspecto al parecer se debe a la rápida tasa de pasaje que contribuye a la disminución en la producción de este gas. Pero cuando el consumo no es adecuado este efecto se pierde (13).
Efecto de las dietas forrajeras tropicales en las emisiones de metano
Según Montenegro y Abarca (20), en ensayos realizados en Costa Rica, el Kikuyo (Pennissetum clandestinum) presenta una mayor eficiencia en la producción de leche que la Estrella Africana (Cynodon nlenfuensis), en gran parte por su menor producción de metano. Igualmente se concluye, que para ambas pasturas la producción de metano es mayor a medida que la edad de pastoreo se aumenta, debido al aumento en el material lignocelulósico.
La emisión total anual de metano que se produce como consecuencia del consumo y del proceso digestivo del pasto, independiente de la especie forrajera, varía en función de los días de rebrote que presenta la pradera. La magnitud de la emisión es diferente según la especie ofrecida. Así, se ha determinado que cuando en la dieta de los bovinos en pastoreo se dsipone de leguminosas, se mejoran los parámetros productivos concomitantes con una disminución en las producciones de metano (20), lo que permite visualizar a los sistemas silvopastoriles como una buena alternativa.
Se ha señalado, que como el H2 y el formato son sustratos para la metanogénesis se puede pensar que los procesos de reducción de componentes nitrogenados pueden competir por esta reducción de sustratos, y desviarla de la biosíntesis de metano (2). Es factible pensar en el efecto positivo de las leguminosas forrajeras en la dieta de los rumiantes, por su aporte de nutrientes, y por su efecto en la disminución de la metanogénesis.
Según DeRamus et al (8), la mejor estrategia para mitigar la producción de metano, es a través de metodologías que mejoren la eficiencia de la energía de los alimentos. La opción de reducción de emisiones de metano consiste en la sustitución de tecnologías convencionales por alternativas concomitantes con una adecuada producción y mínimos efectos medioambientales. La implementación de prácticas de manejo en las pasturas que mejoren su calidad, incrementan la productividad y generalmente tienen un efecto significativo en la reducción de las emisiones de metano.
Parece evidente, el efecto positivo que tiene el balance de dietas con leguminosas y otras especies arbóreas, en la producción de metano en los sistemas silvopastoriles. Estos sistemas que involucran leguminosas rastreras, arbustivas o arbóreas, y otros tipos de especies con potencial alimenticio, se han considerado una alternativa de amplia viabilidad con animales bajo pastoreo de gramíneas tropicales de baja calidad. Además, se consideran otros aspectos que redundan en una mejoría de las características edáficas y de bienestar en el animal.
En trabajos como el reportado por Hess et al (12) se indica que la liberación de metano se puede reducir con el uso de frutos del árbol tropical Sapindus saponaria cuando se suministra en dietas con pastos de baja calidad con o sin suplementación de leguminosa. Abreu et al (1) señalan sin embargo, que el uso del mismo árbol (8% de fruto, 5% de pericarpio o 1.2% de extracto de saponinas semipurificadas, en base seca de la dieta basal) en una dieta compuesta por Brachiaria dictyoneura (60%) y Cratylia argentea (40%) no mostró efectos sobre la disminución de las emisiones de metano. Los autores señalan que aunque el efecto de los tratamientos sobre la metanogénesis no fue alta, si se observó una reducción significativa en la liberación de metano en relación con la cantidad degradada de materia seca y materia orgánica. In vivo, esto podría reflejarse en una reducción de metano emitido por unidad de proteína animal producida, que sería útil aun cuando la cantidad total de metano emitido por animal no disminuya. Moss et al (22) señalan que, las saponinas mencionadas, tienen un efecto significativo en la disminución o eliminación de los protozoos del rumen sin inhibir la actividad bacteriana. Igualmente señalan que las saponinas son glucósidos los cuales aparentemente interactúan con el colesterol presente en la membrana celular de eucarióticas pero no en las células procarióticas.
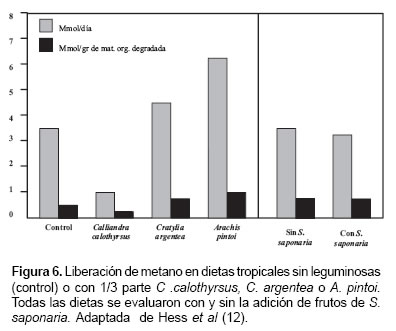
Hess et al (12), señalan que los taninos presentes en muchas leguminosas, como Calliandra calothyrsus, pueden estar asociados con la reducción en la producción de metano, hasta en un 50%, respecto a la dieta tradicional de pasto solo. Pero esto no está asociada a todas las leguminosas de potencial forrajero en el trópico, así, estos autores reportan que especies como Cratylia argentea y Arachis pintoi incrementaron los niveles de metano, hasta 3-4 veces la cantidad determinada en dietas con solo pasto (véase Figura 6).
Metodologías para la determinación de producción y emisiones de metano
Para desarrollar estrategias que mitiguen las emisiones de metano en la ganadería, debe ser posible cuantificar estas emisiones bajo un amplio rango de circunstancias. El metano puede ser medido usando espectroscopía infrarroja, cromatografía de gas, espectroscopía de masa y técnicas de diodo láser (13). Las mediciones de metano son difíciles de realizar sin cámaras respiratorias; una alternativa es estimar el metano a través de cálculos. Esto usualmente se realiza por ecuaciones de regresión de consumo de energía digestible (ED), las cuales ignoran las relaciones de ácidos grasos volátiles y el balance de carbono. Esto conlleva a que los valores de energía metabolizable (EM) puedan no ser buenos estimativos de la producción de metano (29).
Hay diversas opciones para medir las emisiones de metano in vivo. El muestreo individual o a nivel grupal puede ser efectuado utilizando técnicas con cámaras cerradas o métodos con trazadores.
Técnicas de medición
Técnicas cerradas. Las técnicas calorimétricas de respiración tales como las cámaras cerradas, cajas en la cabeza, o capuchas ventiladas y máscaras faciales han sido usadas con efectividad para la determinación de las emisiones de metano (8, 13). Estas emisiones son determinadas por la medición del flujo total de aire por el sistema y la diferencia en la concentración entre el aire inspirado y espirado. En las cámaras, la mayor ventaja radica en las mediciones de metano tanto proveniente de la fermentación ruminal como de la fermentación del tracto posterior. Las desventajas de esta técnica involucra: los costos de construcción y de mantenimiento, la restricción de movimiento de los animales y la alta mano de obra (13). Además, McCaughey et al (18) y Kinsman et al (15), señalan que la mayoría del ganado se encuentra bajo pastoreo de 5 a 12 meses del año, dependiendo de la localización geográfica, lo que implica amplias diferencias de comportamiento alimenticio y cinética ruminal que pueden afectar el consumo voluntario y la tasa de pasaje, difiriendo así, de lo observado en los sistemas cerrados tipo cámaras metabólicas. Otro aspecto que señalan McCaughey et al (19), es que los animales en pastoreo pastan por largos períodos de tiempo, hasta un total de 8-12 h/ día.
Técnicas con trazadores. Las técnicas con trazadores isótopos y no isótopos son útiles. Los métodos con isótopos involucran el uso de [3H-] metano o [14C-] metano y animales canulados a rumen. La mayor limitación con trazadores isótopos es la dificultad en la preparación de la solución por la baja solubilidad del metano. Las técnicas con trazadores no isótopos también son útiles en la medición de producción de este gas. Un ejemplo de estas técnicas es el exafluorosulfuro (SF6), un gas trazador inerte colocado en el rumen. Todas estas técnicas tienen la ventaja que no limita al animal en su comportamiento normal en pastoreo (13, 18, 19). Respecto a esta técnica McCaughey et al (19) y DeRamus et al (8), reportan que puede determinar del 93-98% del total de metano producido, comparado con las cámaras de respiración. DeRamus et al (8), señalan que esta técnica es sencilla, en la cual muestras de gas eructado son continuamente obtenidas a través de un tubo capilar, conectado a un colector localizado en el cuello del animal. Luego de la recolección de las muestras el colector es presurizado con nitrógeno (N2) y con cromatografía de gases se determina el metano y el SF6.
Ecuaciones de predicción. Un método desarrollado en 1960 por Wolin permite calcular las emisiones de metano a través de la distribución molar de los AGV. El balance fermentativo se ha usado para predecir la producción de metano por la conversión de carbohidratos de la dieta a AGV. Esta metodología asume que todo el exceso de H2 es convertido en metano y no hay hidrógeno asociado con la síntesis de células microbiales (13, 28) y que de la fermentación de los sustratos no carbohidratados no se producen AGV. Van Kessel y Russell (28), indican que cuando las células microbiales son incluidas en la estequiometría de la fermentación, los estimativos de la producción de metano disminuyen. Como es lógico esta técnica es criticada por muchos autores, pero es útil para propósitos comparativos (13).
Las características del alimento también se usan para calcular la producción de metano. La ecuación de Blaxter y Claperton formulada en 1965, consideró inicialmente las características del alimento y es la base de la cual la mayoría de los estimativos de producción de metano se han derivado. Otra ecuación fue propuesta por Moe y Tyrrel en 1979, la cual también incorpora las características del alimento. Se deriva de mediciones realizadas en ganado con raciones diarias de alta calidad y su relación con residuos solubles, hemicelulosa y celulosa en la producción de metano (13). Esta se determina:
CH4 = 3.406 + 0.510 (residuo soluble) + 1.736 (hemicelulosa) + 2.648 (celulosa)
Donde CH4 está en Megajoules/día y los residuos solubles, hemicelulosa y celulosa en Kkg/día. Sinembargo, Johnson y Johnson (13) anotan que es poco probable que una ecuación simple basada en las características del alimento tenga una predicción exacta de la producción de metano bajo condiciones perturbadoras en campo. Van Soest (29) señala que las nuevas ecuaciones implementadas en la estimación de producción de metano indican que la digestibilidad del carbohidrato (hemicelulosa más que almidón) tiene efecto sobre la producción de metano.
En términos generales Benchaar et al (3), señalan que la ecuaciones de predicción de producción de metano en el ecosistema ruminal requieren información de consumo de materia seca, composición química de la dieta (incluyendo solubilidad y tasa de degradabilidad) y otras variables como la tasa de pasaje de las fracciones sólida y líquida del rumen, volumen ruminal y pH del fluido ruminal.
Métodos fermentativos in vitro. Los estudios en fermentación y digestión juegan un papel crucial en los estudios nutricionales y fisiológicos en rumiantes. Desde la década de los 50´s muchos métodos han sido desarrollados para simular el ecosistema ruminal. Auque los estudios in vivo han sido de gran importancia, las simulaciones in vitro del medio ambiente ruminal son frecuentemente efectivas y eficientes por su rapidez y bajo costo de operación. Además, porque se pueden definir factores específicos, que en condiciones in vivo, pueden ocultarse por una gran complejidad de factores (14).
Dentro de las técnicas más conocidas in vitro están la de Tilley y Terry implementada en 1963, o sus diversas modificaciones. Entre algunas desventajas de estos métodos se tienen: largo tiempo requerido para realizar un análisis, la gran cantidad de pasos y que la muestra no tiene flujo de recambio (26)
Rumen artificial (RUSITEC). Esta hace parte de las técnicas in vitro, con modificaciones que permiten una mayor similitud a las características del ecosistema ruminal. Kajikawa et al (14) indican que en la década de los 70´s se desarrollaron dos de los sistemas de flujo continuo más adoptados por los investigadores: el “sistema de cultivo de flujo continuo doble” originalmente desarrollado por Hoover et al y el “Rusitec” desarrollado por Czerkawsky y Breckenridge.
Esta última metodología ha demostrado una adecuada correlación con los datos obtenidos in vivo, involucrando como ya se mencionó, mayor rapidez y menos costos, además de una mayor independencia de animales canulados.
Diversos autores reportan el diseño del equipo y la metodología de su uso, pero no existe un manual unificado, debido a que su principal restricción está en que no es un equipo comercialmente disponible. En términos generales es un equipo que normalmente consiste de ocho sistemas de flujo continuo donde interactúan la fase líquida y sólida del rumen (obtenida del rumen de animales donadores), las muestras a analizar (confinadas en bolsas de nylon) y un sistema buferante o de saliva artificial que permite en el sistema un control del pH, a través de sus componentes tipo carbonatos y fosfatos (14).
En cada sistema compuesto por vasos secuenciales, generalmente con una capacidad aproximada de un litro, se colocan más o menos 500 ml. de fluido ruminal con 200 ml. de saliva artificial y 100 ml. de agua, posteriormente agregando agua hasta un nivel total de un litro. Inicialmente, una porción sólida de contenido ruminal se adiciona (80 a 100 gr./litro) para garantizar una fuente de microorganismos que habitan en esta fracción (5, 14).
Czerkawsky y Breckenridge, citados por Kajikawa et al (14), sugieren que un período de acople en la fermentación entre cuatro y seis días debe ser adecuado, aunque hasta 11 días se requieren en algunos casos cuando se evalúan dietas de baja calidad. Una semana de período de adaptación es probablemente suficiente para dietas de buena calidad. Aunque Wettstein et al (31), reportan que en algunas investigaciones se ha considerado suficiente un período de adaptación de tres días.
La cantidad sugerida de alimento a evaluar está entre 12 y 18 gr. (10.3 - 15.5 gr. de materia seca) en cada sistema, porque la eficiencia de crecimiento microbial cambia curvilíneamente cuando la cantidad de alimento se incrementa. Dicho material se ubica en bolsas de nylon, similares a las utilizadas en los métodos in situ, las cuales generalmente tienen un tamaño de poro entre 40 y 100 mm (14).
La tasas de dilución mayores a 4%/hora pueden diluir los productos de fermentación en el Rusitec, mientras que una tasa de dilución más baja de 2% por hora puede conducir a una disminución en el pH debido a la acumulación de productos finales. Por lo tanto la mayoría de los estudios asumen tasas entre estos valores (14). Así, Carro y Miller (4) reportan que el flujo a través de los vasos se mantuvo con una infusión continua de saliva artificial (McDougall, pH 8.4) a una tasa de 2.9%/h.
Esta metodología permite múltiples tipos de análisis fermentativos, fisiológicos, microbianos, etc. extrapolables a rumen. Entre muchas mediciones, se encuentra la determinación de producción de gases en el sistema.
Producto de la fermentación se produce, en un tiempo determinado, un volumen de gas que es almacenado de forma individual por fermentador en bolsas con película de fluoropolivinil. La producción de metano, presente en el volumen total de cada bolsa, se determina por diferentes métodos, siendo la cromatografía de gases el más utilizado en este caso. Para este efecto las bolsas se acoplan al sistema cromatográfico, en el cual se determina el porcentaje de gas en la muestra y se valora la cantidad total teniendo en cuenta el volumen total de gas en la bolsa (1, 14)
Consideraciones finalesLas emisiones de metano por los rumiantes tienen un efecto considerable a nivel medioambiental debido al aporte que este gas hace al calentamiento global y a la disminución de la capa de ozono, aspectos que llevan a cambios climáticos que afectan drásticamente, entre otros, a los sistemas de producción agrícola y pecuaria. Otro aspecto relevante observado en los sistemas de producción ganadera es la energía alimenticia que se transforma en forma de gas metano y no es aprovechada por el animal. Considerando las situaciones anteriores, es importante plantear alternativas que disminuyan estas emisiones, mejorando las condiciones productivas de los sistemas ganaderos, pero a la vez intentando aminorar los efectos en la contaminación medioambiental.
Considerar la alternativa de disminuir la población bovina, en aras de disminuir los efectos medioambientales bajo las condiciones tropicales, donde la baja productividad por condiciones edáficas y baja disponibilidad de recursos alimenticios de buena calidad es una constante, no se considera una alternativa viable, máxime cuando el incremento de la población humana mundial y principalmente en los países tropicales, demandan un alto suministro de fuentes alimenticias de origen animal. Por tal motivo, la implementación de técnicas alimentarias en la producción bovina acorde a las condiciones del trópico, se deben basar en la búsqueda de alternativas factibles y que propendan por mejorar las características de la dieta y por ende, los parámetros fermentativos en nuestros ganados.
Considerando las condiciones generales de la dieta de los bovinos en el trópico, la implementación de sistemas que mejoren sus características, deben ser un constante reto del personal involucrado en esta área del sector pecuario. Las diferentes alternativas involucradas en el manejo de las pasturas y en la suplementación a la dieta forrajera, que involucren cambios en los parámetros fermentativos, deben ser consideradas para obtener mejores parámetros productivos simultáneos con menores emisiones de metano.
Bajo las condiciones mencionadas, los sistemas silvopastoriles pueden ser una alternativa para mejorar la calidad de la dieta y por lo tanto disminuir las emisiones de gases, principalmente de metano. Estos sistemas se convierten en alternativas interesantes desde el punto de vista de ampliación del rango de nutrientes disponibles para el ganado bovino y en la mejoría de los parámetros fermentativos a nivel ruminal, implicando mayor eficiencia en el uso de los nutrientes y por ende unos mayores parámetros productivos. Sin embargo, son pocos los estudios que se han realizado al respecto, reportándose resultados contradictorios según la especie forrajera utilizada. A la par que se implementan estos sistemas, con visión hacia la mejoría de los aspectos ya mencionados, se debe acudir a metodologías con rigor científico que permitan una cuantificación y cualificación acertada de los parámetros fermentativos, entre ellos las emisiones de metano.
En esta revisión, punto de partida para asumir un proceso investigativo en producción y medición del gas metano en los sistemas silvopastoriles, se asume como una alternativa confiable el uso del método in vitro RUSITEC (rumen artificial), el cual permite la cuantificación de diversos parámetros fermentativos, entre ellos la producción de metano de las dietas propias del trópico. Sistema que a pesar de obviar muchos aspectos inherentes al animal y a su entorno, permite conocimiento de la ecofisiología ruminal de nuestros una adecuada correlación de los resultados con los ganados, bajo condiciones medioambiantales propias que se obtienen in vivo y una mayor aproximación al de trópico.
Summary
The gas methane in the cattle production and alternative to measure its emissions and to lessen its impact at environmental and productive level
The ruminants are great contributors to the global heating and deterioration of the ozone layer, by the liberation of high amounts of gases to the atmosphere, among them, the carbon dioxide gas and the methane. The methane produced in rumen is generated mainly by fermentative processes of the food that enters to rumen. The main biotic factor at level of rumen in the methane production are the metanogenic anaerobic bacteria. These bacterias use different substrates for the methane production, but the main ones are the H2 and the CO2. The elimination of these gases, mainly of the H2 implies the removal of a factor implied in the stability of pH ruminal being this essential for an optimal fermentation. But at the same way this methane production is considered like a loss of potentially usable energy. The effects of the metanogenic bacteria are employees mainly of the present substrates in the diet and of the interactions with other populations. Interventions in the feeding offered to the animals, oriented towards optimizing the process of ruminal fermentation, repel generally in an improvement of the productive and reproductive parameters, had, among other aspects, to one better use of the energy. In addition, the diminution of the emissions to the atmosphere receives great importance in the protection of the environment. The diverse evidences show that the rate of methane emission by ruminal fermentation, is related to the characteristics physical-chemistries of the diet, which affect the level of consumption and the frequency of feeding. By this the subnutrition contributes to increase the methane emissions. The possibility of limiting the methane emissions by the cattle in tropical production systems, provides economic and environmental benefits. A reduction option consists of the substitution of conventional technologies by new concomitants alternatives with a suitable production and minimums environmental effects. Within these alternatives of potential use in tropic it is the handling of pastures, tending towards improving his quality. An alternative of wide potential and that until the moment has taken force by its multiple benefits are the silvopastoral systems, but little has investigated its effect on the ruminal methane production. For the determination of emissions and the efficiency of the implemented alternatives, one is due to resort to appropriate methodologies that allows to extrapolate the results to the real conditions in alive. Within these alternatives, the system in vitro RUSITEC, has presented a high correlation with the own characteristics of the animal. The objective of this revision is to visualize different factors that determine the methane emissions, measurement alternatives and options to diminish the production of this gas in the cattle systems, for the sake of a more efficient production and than it protects the environment.
Key words: greenhouse effect, methane generation, ruminants, rusitec, silvopastoral system.
Referencias
1. Abreu A, Carulla JE, Kreuzer M, Lascano C, Díaz TE, Cano A, Hess, HD. Efecto del fruto, del pericarpio y del extracto semipurificado de saponinas de Sapindus saponaria sobre la fermentación ruminal y la metanogénesis in vitro en un sistema RUSITEC. Rev Col Cienc Pec. 2003; 16: 147-154
[ Links ]2. Anderson RC and Rasmussen MA. Use of a novel nitrotoxinmetabolizing bacterium to reduce ruminal methane production. Bioresource Technology. 1998; 64: 89-95
[ Links ]3. Benchaar C, Rivest J, Pomar C, Chiquette J. Prediction of methane production from dairy cows using existing mechanistic models and regression equations. J Anim Sci 1998; 76: 617- 627.
[ Links ]4. Carro MD, Miller EL. Effect of supplementing a fibre basal diet with different nitrogen formus on ruminal fermentation and microbial growth in an in sacco semicontinuous culture system (RUSITEC). British Journal of Nutrition, 1999; 82: 149-157
[ Links ]5. Czerkawski JW, Breckenridge G. Design and development of a long-term rumen simulation technique (Rusitec). British Journal of Nutrition, 1977; 38: 371-384.
[ Links ]6. Chandramoni SB, Jadhao CM, Tiwari CM, Khan MY. Energy metabolism with Particular reference to methane production in Muzaffarnagari sheep fed rations in roughage to concentrate ratio. Animal Feed Science and Technology, 2000; 83: 287- 300
[ Links ]7. Dehority BA, Tirabasso PA. Antibiosis between ruminal bacteria and ruminal fungi. Appl Environ Microbiol 2000; 66(7): 2921-2927
[ Links ]8. DeRamus HA, Clement TC, Giampola DD, Dickison PC. Methane emissions of beef cattle on forages: efficiency of grazing management systems. Journal Environ Qual, 2003; 32: 269-277
[ Links ]9. Dohme F, Machmüller A, Wasserfallen A, Kreuzer M. Comparative efficiency of various fats rich in medium-chain fatty acids to suppress ruminal methanogenesis as measured with RUSITEC. Canadian Journal of Animal Science, 2000; 80: 473-482.
[ Links ]10. Fonty G. The rumen anaerobic fungi. In: Jouany, JP. Rumen microbial metabolism and ruminant digestion. INRA editions, Paris, France, 1991; pp 53-70
[ Links ]11. González F, Rodríguez H. Proyecciones de las emisiones de gases de efecto de invernadero (GEI) Colombia 1998-2010. Rev Acad Colomb Cienc, 1999; 23 (89): 497-505
[ Links ]12. Hess HD, Monsalve LM, Carulla JE, Lascano CE, Díaz TE, Kreuzer M. In vitro evaluation of the effect of Sapindus saponaria on methane release and microbial populations (1.4.1). 2002. URL: http://www.ciat.cgiar.org/forrajes/pdf/output1_2002.pdf
[ Links ]13. Johnson KA, Johnson DE. Methane emissions from cattle. J Anim Sci, 1995; 73: 2483-2492
[ Links ]14. Kajikawa H, Hai J, Terada F, Suga T. Operation and characteristics of newly improved and marketable artifical rumen (Rusitec). In: Memoirs of National Institute of Livestock and Grassland Science. Nº 2. Mar. 2003
[ Links ]15. Kinsman R, Sauer FD, Jackson HA, Wolynetz, MS. Methane and carbon dioxide emissions from cows in full lactation monitored over a six-month period. J Dairy Sci, 1995; 78 (12): 2760-2766
[ Links ]16. Kurihara M, Magner T, McCrabb H, McCrabb G. Methane production and energy partition of cattle in the tropics. British Journal of Nutrition, 1999; 81: 227-234.
[ Links ]17. López S, McIntosh FM, Wallace RJ, Newbold CJ. Effect of adding acetogenic bacteria on methane production by mixed rumen microorganisms. Animal Feed Science and Technology, 1998; 78: 1-9.
[ Links ]18. McCaughey W, Wittenberg K, Corrigan D. Methane production by steers on pasture. Can J An Sc, 1997; 76 (3): 519-524.
[ Links ]19. McCaughey W, Wittenberg K, Corrigan D. Impact of pasture type on methane production by lacting beef cows. Can J An Sc, 1999; 79 (2): 221-226.
[ Links ]20. Montenegro J, Abarca S. Fijación de carbono, emisión de metano y de óxido nitroso en sistemas de producción bovina en Costa Rica. En: Intensificación de la ganadería en Centroamérica: beneficios económicos y ambientales. CATIE- FAO - SIDE. Ed Nuestra Tierra. 2000. 334 p.
[ Links ]21. Moss AR, Givens DI. The effect of supplementing grass silage with soya bean meal on digestibility, in sacco degradability, rumen fermentation and methane production in sheep. Animal Feed Science and Technology, 2002; 97: 127- 143
[ Links ]22. Moss AR, Jouany JP, Newbold J. Methane production by ruminants: its contribution to global warming. INRA EDP Sciences. Ann Zootech, 2000; 49: 231-253
[ Links ]23. Preston TR, Leng RA. Friendly development. Livestock Research for Rural Development, 1(1), November, 1989. URL: http://www.cipav.org.co/lrrd/lrrd1/1/preston.htm
[ Links ]24. Primavesi O, Shiraishi RT, Dos Santos M, Aparecida M, Teresinha T, Franklin P. Metano entérico de bovinos leiteiros em condiçðes tropicais brasileiras. Pesq agropec bras, 2004 39 (3): 277-283
[ Links ]25. Stewart CS. The rumen bacteria. In: Jouany, JP. Rumen microbial metabolism and ruminant digestion. INRA editions, Paris, France, 1991; 15-26
[ Links ]26. Tejido ML, Ranilla MJ, Carro MD. In vitro digestibility of forages as influences by source of inoculum (sheep rumen versus Rusitec fermenters) and diet of the donor sheep. Animal Feed Science and Technology, 2002; 97: 41-51
[ Links ]27. Van Kessell JS, Russell JB. The effect of pH in vitro methane production from ruminal bacteria (Abstract # 2). In: 23rd Biennial Conference on Rumen Function. Chicago, Illinois, E.U. November 14-16, 1995; Vol. 23.
[ Links ]28. Van Kessell JS, Russell JB. The effect of pH on ruminal methanogenesis. FEMS Microbiology Ecology, 1996; 20: 205-210.
[ Links ]29. Van Soest PJ. Nutritional ecology of the ruminant. Second edition. Cornell University Press, 1994; 476 p.
[ Links ]30. Weimer PJ. Manipulating ruminal fermentation: a microbial ecological perspective. J Anim Sci, 1998; 76: 3114 - 3122
[ Links ]31. Wettstein HR, Machmüller A, Kreuser M. Effects of raw and modified canola lecithins compared to canola oil, canola seed and soy lecithin on ruminal fermentation measured with rumen simulation technique. Animal Feed Science and Technology, 2000; 85: 153-169.
[ Links ]32. Yokoyama MT, Johnson KA. Microbiología del rumen e intestino. En: Church DC. El rumiante: fisiología digestiva y nutrición. Editorial Acribia S.A. Zaragoza, España, 1993; 137-156
[ Links ]

